春小麦脂肪氧化酶TaLox-B位点等位变异检测及分布规律
2021-03-21董一帆严勇亮库尼都孜阿依吐尔汗战帅帅耿洪伟
董一帆 严勇亮 谢 磊 库尼都孜阿依·吐尔汗 战帅帅 耿洪伟*
(1.新疆农业大学 农学院/生物技术重点实验室,乌鲁木齐 830052;2.新疆农业科学院 农作物品种资源研究所,乌鲁木齐 830091)
我国消费者的饮食习惯多以面制品为主,对于面粉色泽要求较高。面粉的色泽与其制品的商品质量密切相关,是衡量小麦品质的重要指标之一[1]。因此对面粉及其制品色泽的研究是小麦品质遗传改良的重要内容。面粉的白度在很大程度上受小麦籽粒中脂肪氧化酶(lipoxygenase, 简称LOX)活性高低的影响[2-3]。小麦籽粒中LOX能通过偶联氧化小麦面粉中的类胡萝卜素,进而使面粉色泽变白,起到漂白的效果[4]。同时,小麦籽粒中LOX的活性高低还能影响小麦籽粒储藏期长短和面筋强度等特性[2,5-7],由于低LOX活性显著减缓脂质降解,因此使小麦籽粒氧化变质时间得以推迟,从而有助于延长小麦籽粒的储藏期[6]。高LOX活性在LOX催化反应过程中能将更多蛋白质分子中的巯基(-SH)氧化为二硫键(-S-S),因此蛋白质分子能够高度聚合,防止面筋蛋白水解,从而有助于增强面筋强度[5,7]。
LOX蛋白家族,是含非血红素铁的一类蛋白质,属于多元不饱和脂类的酶系,在自然界中普遍存在[8-9],参与植物的生长发育和一些抵御逆境胁迫的生理过程。目前,对小麦LOX已有大量的研究报道,研究表明环境和基因型对LOX活性均有影响,但其活性主要受基因型影响[10-11]。Geng等[12-13]研究发现小麦4BS染色体上的TaLox-B1位点是LOX活性的主效基因,并基于此位点基因的序列多态性开发了互补的显性功能标记LOX16和LOX18,可用于区分TaLox-B1位点上的等位变异TaLox-B1a和TaLox-B1b。用LOX16标记对材料进行扩增,可以得到489 bp的片段,表明材料含有优异等位变异TaLox-B1a类型,用LOX18标记对材料进行扩增,可以得到791 bp的片段,表明材料含有等位变异TaLox-B1b类型。杨杰等[14]和相吉山等[15]用LOX16和LOX18标记分别对宁夏回族自治区180份和新疆维吾尔自治区195份小麦种质资源进行标记检测,发现该标记能在供试材料中清晰且稳定地扩增出489和791 bp的条带,能够准确鉴定出材料的TaLox-B1a和TaLox-B1b基因类型。随后Zhang等[16]利用同源克隆技术在小麦4BS染色体上克隆出TaLox-B2和TaLox-B3基因,并基于TaLox-B2和TaLox-B3基因开发了共显性标记LOX-B23。利用标记LOX-B23对材料进行扩增,若同时扩增出788和677 bp的片段,则该材料为等位变异TaLox-B2a和TaLox-B3a类型;只能扩增出788 bp的单一片段,则该材料为TaLox-B2a和TaLox-B3b等位变异;只能扩增出660 bp的单一片段,则该材料为等位变异TaLox-B2b和TaLox-B3b类型。LOX-B23标记能有效区分TaLox-B2和TaLox-B3基因位点上的不同等位变异,可应用于小麦LOX基因活性的筛选。Zhang等[16]利用LOX16、LOX18和LOX-B23等标记对我国小麦种质资源进行了TaLox-B位点等位变异检测和LOX活性的分析,结果表明在TaLox-B位点的等位变异基因组合中TaLox-B1a/TaLox-B2a/TaLox-B3a或TaLox-B1b/TaLox-B2a/TaLox-B3a,TaLox-B1a/TaLox-B2a/TaLox-B3b或TaLox-B1b/TaLox-B2a/TaLox-B3b和TaLox-B1a/TaLox-B2b/TaLox-B3b,分别与LOX高、中等和低活性显著正相关。白璐等[17]研究表明等位变异基因组合TaLox-B1b/TaLox-B2b/TaLox-B3b与高LOX活性负相关。
虽然目前利用与LOX活性相关的功能标记LOX16、LOX18和LOX-B23对新疆小麦种质资源TaLox-B位点的等位基因分布已有报道[15,17],但研究的小麦类型多为冬小麦。目前对国内外春小麦种质资源中TaLox-B位点等位基因分布特征的研究鲜有报道。本研究利用LOX16、LOX18和 LOX-B23 等功能标记对237份国内外春小麦品种(系)的TaLox-B位点进行检测,旨在分析脂肪氧化酶TaLox-B位点等位基因在国内外不同地区春小麦种质资源中的分布规律,以期为春小麦面粉色泽遗传改良提供参考。
1 材料与方法
1.1 供试材料
供试237份春小麦品种(系)由新疆农科院农作物品种资源研究所严勇亮研究员课题组提供,其中国内新疆春小麦地方品种(系)154份,国外春小麦品种(系)83份。新疆春小麦生态类型多样,一般分为南疆、北疆、东疆和伊犁河谷等4个生态区。新疆154份春小麦地方品种(系)来自新疆的北疆95份、南疆31份、东疆20份和伊犁河谷8份;国外春小麦品种(系)来自于亚洲其他国家8份、欧洲33份、美洲29份、非洲7份和大洋洲6份。
1.2 基因组DNA提取
每个小麦品种(系)均取3粒完整且大小均匀的籽粒,粉碎后置于2.0 mL的离心管中,按照SDS方法[18]提取DNA。
1.3 PCR扩增及检测
利用Geng等[13]和Zhang等[16]开发的功能标记LOX16、LOX18和LOX-B23,检测小麦4B染色体上TaLox-B位点的等位变异。引物由上海生工生物工程技术服务有限公司合成,见表1。
以小麦基因组DNA为模板,在Eppendorf AG PCR仪(Eppendorf,德国)上进行PCR扩增。15 μL 的PCR反应体系:以1.0 μL的DNA为模板,2×Es Taq MasterMix(Dye)7.5 μL(北京康为世纪生物科技有限公司),上、下游引物(10 μmol/L)各1.0 μL,用ddH2O补充至15.0 μL。按照白璐等[17]LOX16、LOX18和LOX-B23标记的扩增程序进行PCR扩增。PCR扩增产物用1.5%的琼脂糖凝胶电泳分离,ddH2O漂洗后用VILBER LOURMAT凝胶成像系统拍照并记录。

表1 检测LOX的 TaLox-B位点等位变异的标记信息Table 1 Marker′s information of allelic variation at TaLox-B locus of LOX was detected
1.4 统计方法
选取符合目的条带大小、条带清晰且无杂带的PCR产物,进行统计,根据此结果判断该品种的TaLox-B位点等位基因的变异类型。若某一品种(系)的3粒籽粒PCR产物条带大小不一致,则需重新提取该品种(系)的DNA,重新进行PCR检测,以3次结果一致为准。
2 结果与分析
2.1 TaLox-B位点等位基因变异类型及分布频率
由图1可知,所选国内和国外材料在TaLox-B1位点上,TaLox-B1b变异类型的分布频率分别为95.5%和85.5%,远高于优异等位变异类型TaLox-B1a(与高LOX活性极显著正相关)的频率(平均8.0%),因此在TaLox-B1位点上,优势等位变异(分布频率较高的等位变异类型)均为与低LOX活性相关的TaLox-B1b变异类型,说明国内外材料具有优异等位变异TaLox-B1a的分布频率均很低。但国外材料具有的优异等位变异TaLox-B1a的频率(14.5%)远高于国内材料(4.5%),说明国外春小麦品种(系)具有的高LOX活性基因TaLox-B1a的分布频率远高于国内新疆春小麦地方品种(系)。国内外供试小麦材料在TaLox-B2和TaLox-B3位点,TaLox-B2a和TaLox-B3a变异类型的分布频率分别为93.2%和86.1%,为优势等位变异,其中,国内材料的优异等位变异TaLox-B2a和TaLox-B3a的分布频率均高于国外材料。
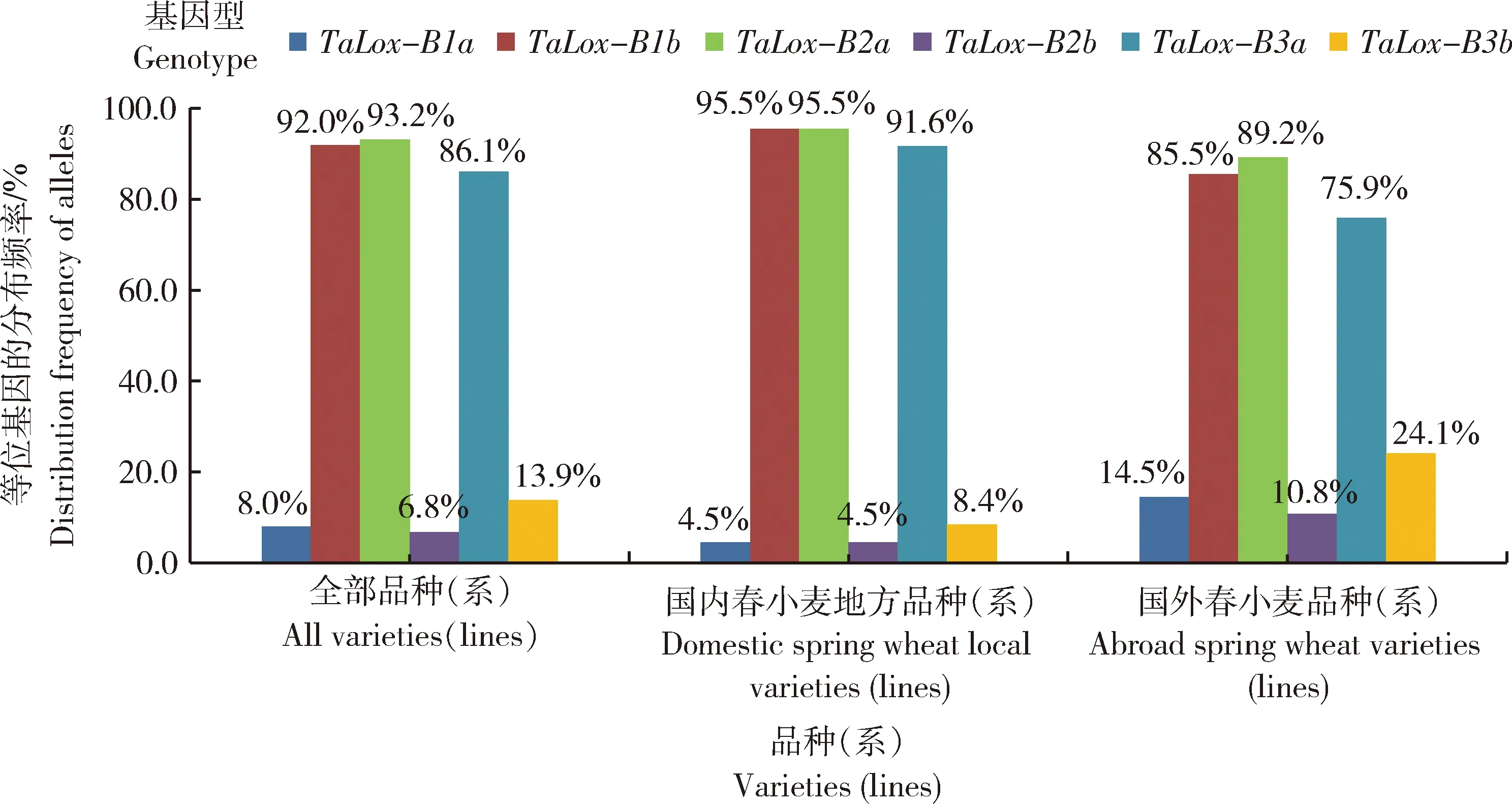
图1 春小麦的TaLox-B位点等位基因的分布频率Fig.1 Distribution frequency of TaLox-B locus alleles in spring wheat
2.2 新疆不同生态区的春小麦TaLox-B位点等位基因的分布及其组合类型频率
对154份新疆不同生态区的春小麦TaLox-B位点进行等位变异检测结果表明,在TaLox-B1位点,优异等位变异类型TaLox-B1a的材料7份(4.5%)。在TaLox-B2和TaLox-B3位点,优异等位变异类型TaLox-B2a和TaLox-B3a的材料分别有147(95.5%)和141份(91.6%)。优异等位变异TaLox-B1a的分布频率表现为北疆地区(6.3%)>南疆地区(3.2%)>整体水平(4.5%);优异等位变异TaLox-B2a在伊犁河谷、南疆、北疆和东疆地区逐次升高(分布频率依次为:75.0%、93.5%、96.8%和100.0%),优异等位变异TaLox-B3a在伊犁河谷、北疆、南疆和东疆地区逐次升高,分布频率依次为:75.0%、90.5%、93.5%和100.0%,见图2。
154份新疆不同生态区的春小麦地方品种(系)共检测出5种基因型组合,TaLox-B1a/TaLox-B2a/TaLox-B3a、TaLox-B1b/TaLox-B2a/TaLox-B3a、TaLox-B1a/TaLox-B2a/TaLox-B3b、TaLox-B1b/TaLox-B2a/TaLox-B3b和TaLox-B1b/TaLox-B2b/TaLox-B3b,其频率分别为3.9%、87.7%、3.2%、0.6%和4.6%。新疆不同生态区的春小麦地方品种(系)具有的最高LOX活性基因型组合TaLox-B1a/TaLox-B2a/TaLox-B3a、TaLox-B1b/TaLox-B2a/TaLox-B3a在伊犁河谷、北疆、南疆和东疆地区逐次升高,分布频率之和依次为75.0%、90.6%、93.5%和100.0%,见图3。其中20份东疆地区材料均检测为高LOX活性基因型组合TaLox-B1b/TaLox-B2a/TaLox-B3a。
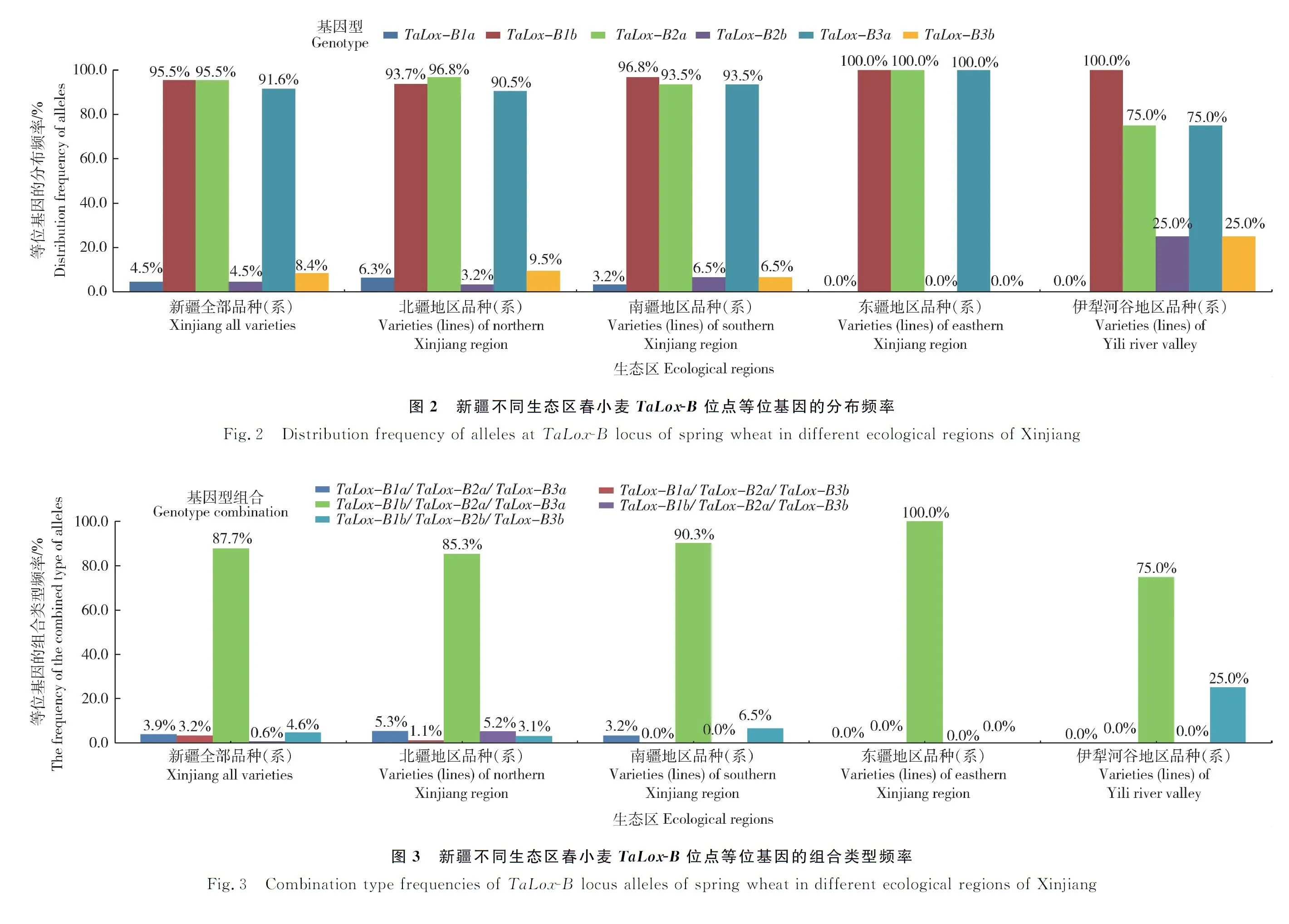
2.3 国外春小麦TaLox-B位点等位基因的分布及其组合类型频率
由图4可知,比较83份国外春小麦品种(系)的TaLox-B1位点,大洋洲地区(主要来自澳大利亚)的材料均为与低LOX活性相关的等位变异类型TaLox-B1b,而优异等位变异TaLox-B1a分布频率在亚洲、美洲、非洲和欧洲地区中依次升高。在TaLox-B2位点,大洋洲地区的材料均为优异等位变异TaLox-B2a。在TaLox-B3位点,美洲地区的材料具有优异等位变异TaLox-B3a的频率(82.8%)高于其他地区(75.9%)。
在83份国外材料中,5种基因型组合TaLox-B1a/TaLox-B2a/TaLox-B3a、TaLox-B1b/TaLox-B2a/TaLox-B3a、TaLox-B1a/TaLox-B2a/TaLox-B3b、TaLox-B1b/TaLox-B2a/TaLox-B3b和TaLox-B1b/TaLox-B2b/TaLox-B3b的频率分别为12.1%、63.9%、10.8%、2.4%和10.8%,见图5。供试的国外春小麦品种(系)具有的高LOX活性基因型组合TaLox-B1a/TaLox-B2a/TaLox-B3a、TaLox-B1b/TaLox-B2a/TaLox-B3a在大洋洲、非洲、亚洲、欧洲和美洲地区的分布频率之和依次为50.0%、71.4%、75.0%、75.8%和82.8%。
2.4 国内外春小麦TaLox-B位点等位变异基因组合频率
在237份国内外春小麦品种(系)中,共检测出5种基因型组合:TaLox-B1a/TaLox-B2a/TaLox-B3a、TaLox-B1b/TaLox-B2a/TaLox-B3a、TaLox-B1a/TaLox-B2a/TaLox-B3b、TaLox-B1b/TaLox-B2a/TaLox-B3b和TaLox-B1b/TaLox-B2b/TaLox-B3b,其频率分别为6.8%、79.3%、5.9%、1.3%和6.7%,见图6。国内外供试品种(系)的优势等位变异基因组合(分布频率较高的等位变异基因组合)为TaLox-B1b/TaLox-B2a/TaLox-B3a,是与高LOX活性显著正相关的优异等位变异基因组合。供试品种(系)中新疆春小麦地方品种(系)所具有的优异等位变异基因组合TaLox-B1a/TaLox-B2a/TaLox-B3a和TaLox-B1b/TaLox-B2a/TaLox-B3a的频率之和为91.6%高于国外春小麦品种(系)(76.0%),但国外春小麦品种(系)具有高LOX活性基因组合TaLox-B1a/TaLox-B2a/TaLox-B3a频率(12.1%)远高于国内新疆春小麦地方品种(系)(3.9%)。
3 讨 论
3.1 功能标记的应用价值
功能标记是基于控制表型性状功能基因内部的单核苷酸多态性(single nucleotide polymorphism, SNP)核苷酸序列多态性位点而开发的分子标记[19],直接反映作物的表型性状,具有便捷、稳定和高效等特点。利用功能标记辅助选择育种能够提高育种效率,缩短育种时间。目前,已大量开发出与品质、产量和抗逆等重要性状有关的功能标记,并广泛应用于育种实践中[20-22],已成为小麦等主要农作物分子育种的重要手段[23]。
功能标记能够准确而快速地检测小麦种质资源所携带的等位基因,提高了育种工作者对小麦特定性状的选择效率,进而加速育种进程。产量、品质和抗性多为复杂的数量性状,多个相关基因的功能标记能对某一性状的选择更加有效,也能提高功能标记的应用价值和分子标记辅助选择的作用。因此,应加大功能标记开发和利用,在针对小麦某一性状时,结合其多个功能标记进行检测,更能准确快速地对小麦种质资源进行筛选,从而提高育种进程。
3.2 小麦LOX的TaLox-B基因的等位变异分布规律
已有研究表明,在TaLox-B1位点,优异等位变异TaLox-B1a的频率远低于TaLox-B1b[14-15,17,11,24-25];在TaLox-B2和TaLox-B3位点,与高LOX活性相关的优异等位变异TaLox-B2a和TaLox-B3a均为优势等位变异[17]。本研究利用LOX16、LOX18和LOX-B23等功能标记对237份国内外春小麦品种(系)进行检测,结果表明,不同TaLox-B位点的等位变异分布与已有的报道[14-15,17,11,24-25]基本一致。新疆不同生态区春小麦地方品种资源中,北疆生态区材料具有高LOX活性TaLox-B1a的基因型频率(6.30%)高于相吉山等[15]研究的新疆春小麦地方品种资源的基因型频率(4.55%)。这可能与相吉山等[15]研究所用的供试春小麦品种数量较少和代表性不强有关,也可能与北疆生态区位于新疆北部较发达地区有关,相较于新疆其他地区而言,育种工作者对小麦色泽品质性状重视度较高和研究起步较早有关。
在不同小麦类型(冬小麦和春小麦)中,TaLox-B1、TaLox-B2和TaLox-B3位点上,冬小麦具有的与高LOX活性相关的TaLox-B1a、TaLox-B2a和TaLox-B3a优异等位变异分布频率均高于春小麦[15,17,24]。由此可见,春小麦的色泽品质相比之冬小麦较差,因此在未来的育种过程中应加大对春小麦的色泽改良力度,以期提高春小麦的色泽品质。
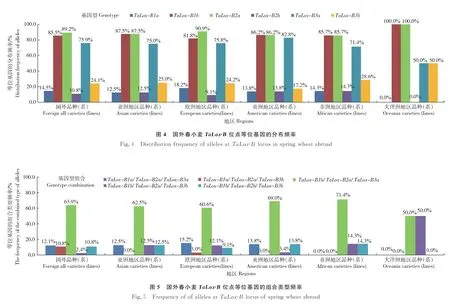
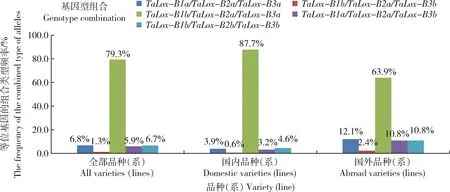
图6 国内外春小麦TaLox-B位点等位基因组合类型的分布频率Fig.6 Distribution frequencies of allele combinations at TaLox-B loci in spring wheat at domestic and abroad
3.3 不同来源的小麦LOX分布规律
本研究利用功能标记LOX16和LOX18对237份国内外春小麦品种(系)进行分子标记检测,结果表明,供试品种(系)中的新疆春小麦地方品种(系)的TaLox-B1a基因型分布频率远低于国外春小麦品种(系),这表明新疆春小麦地方品种(系)具有的与高LOX活性相关的优异等位变异TaLox-B1a分布频率远低于国外水平。即新疆春小麦地方品种在色泽品质方面与国外春小麦品种仍有较大差距。这可能与新疆早期小麦育种目标多以高产为主[26],对小麦色泽品质改良的关注较少有关。因此未来可通过引进国外优良品种和加大地方品种选育等方式缩小与国外良种的差距。
白璐等[17]利用LOX16、LOX18和LOX-B23等标记对24份新疆春小麦自育品种进行检测,结果显示新疆春小麦自育品种具有的最高LOX活性基因型组合TaLox-B1b/TaLox-B2a/TaLox-B3a的频率仅为16.7%。本研究利用上述功能标记对154份新疆春小麦地方品种(系)进行检测,结果表明,新疆春小麦地方品种(系)所含有的高LOX活性的基因型组合TaLox-B1b/TaLox-B2a/TaLox-B3a频率为87.7%,远高于白璐等[17]研究结果。刘三才等[27]的研究结果表明,我国的小麦品种选育的遗传多样性和品质性状的变异性与我国地方品种相比有所下降。虽然自育品种多为地方品种经多代单株自交系统选育的纯合系,但新疆春小麦自育品种在选育的过程中面粉白度较高的品种未被选育保留下来。因此,未来应加大对新疆春小麦地方品种的开发利用。
本研究发现,新疆不同生态区中,东疆地区的材料均检测为含有高LOX活性的TaLox-B1b/TaLox-B2a/TaLox-B3a基因型组合,因此,新疆东疆地区的春小麦地方品种(系)是新疆不同生态区中选育高LOX活性小麦的优质种质资源。供试的国外不同地区春小麦品种(系)中,美洲地区春小麦材料所具有的高LOX活性的TaLox-B1a/TaLox-B2a/TaLox-B3a或TaLox-B1b/TaLox-B2a/TaLox-B3a基因型组合的频率最高,这可能与美洲地区小麦育种家对小麦色泽品质性状研究起步较早有关。因此在对国外优良品种的引进时可倾向于美洲地区的良种。
4 结 论
本研究利用分子标记检测,从237份国内外春小麦品种(系)中,筛选出16份材料具有高LOX活性基因型组合(TaLox-B1a/TaLox-B2a/TaLox-B3a),可作为新疆地区选育高白度面粉的春小麦种质材料,其中10份国外春小麦品种包括‘Kartner Fruhweizen’、‘Kenhi 13627’、‘RL 2327’、‘Super Helyis’、‘Supremo 211’、‘Swedish 880D’、‘White Fife’、‘Brevrt W 972’、‘Kitchener’和‘荷兰小麦’,6份为新疆春小麦地方品种(系)包括‘白春麦 (CM026)’、‘白春麦(CM121)’、‘阿克牙孜干’、‘奇春4号’、‘奇春5号’和‘黑穗红芒’。
