水稻产量对生殖生长阶段不同时期高温的响应差异
2021-03-17宋有金吴超李子煜唐设李刚华王绍华丁艳锋
宋有金 吴超 李子煜 唐设 李刚华 王绍华 丁艳锋,*
水稻产量对生殖生长阶段不同时期高温的响应差异
宋有金1吴超2,*李子煜1唐设1李刚华1王绍华1丁艳锋1,*
(1南京农业大学 农学院,南京 210095;2中国科学院 广西植物研究所,桂林 541006;*通信联系人, E-mail: cwwuchao@njau.edu.cn;dingyf@njau.edu.cn)
研究水稻产量对不同时期不同类型高温的响应差异,为耐热水稻品种的筛选与培育提供依据。在桶栽条件下,于人工气候室内,以耐热水稻品种Nagina 22、汕优63和热敏感品种两优培九为材料,设置三个处理时期,即穗分化期、开花期和灌浆期;四种处理温度,即白天高温、夜间高温、全天高温和适宜温度,研究水稻产量和产量构成因子对不同时期高温的响应。汕优63在三个时期均表现出较强的耐热性;Nagina 22在花期和灌浆期对高温耐性较强,但易受穗分化期高温影响;两优培九对不同时期高温均较敏感。汕优63在不同时期高温下均表现出稳定的伤流强度,而高温敏感品种两优培九的伤流强度受高温影响均显著下降。水稻对不同时期的高温耐性并不具有相关性,筛选或培育耐热品种时应考虑其对不同时期高温的综合耐受性。
水稻;产量;高温;生殖阶段;伤流强度
由人类活动引发的以全球变暖为特征的气候变化正日益威胁着作物生产和粮食安全[1]。世界上近一半的人口以水稻为主食,频繁发生的极端高温事件给水稻生产造成了严重损失[2]。目前我国大部分水稻种植区域已经处于或即将处于高温胁迫当中,预计未来温度仍将以0.02~0.04℃/年的速度增加[1],水稻生长季节高温持续时间延长,胁迫强度增加,极端高温频发,夜间温度上升已成为限制我国水稻增产的重要环境因素[3]。水稻在生殖生长阶段极易受高温胁迫的伤害,但敏感程度在不同时期、不同品种间存在差异。穗分化期高温降低每穗颖花数和结实率[4],开花期高温降低结实率[5],灌浆期高温降低粒重,水稻生殖生长阶段高温最终均降低了产量[6]。然而,以往研究多侧重于某一时期高温下水稻产量变化情况,关于水稻产量及产量构成对不同时期高温的响应差异研究则并不多见。
水稻对昼夜温度敏感性不同,白天高温造成水稻产量降低的最突出原因是结实率下降,对其他产量因子的影响较小;而夜间高温对结实率、每穗颖花数和粒重均有较大的影响[7]。研究表明,夜间高温对水稻籽粒灌浆的伤害比白天高温更加严重,夜间高温降低粒重,而白天高温下表现不显著[8]。此外,夜间高温会增加垩白的形成,影响稻米品质[9]。高温通过影响水稻产量构成因子影响水稻产量,但不同类型高温下水稻产量构成因子变化对产量的贡献程度及基因型差异尚不明确。由于高温发生时间的不确定性,夜间高温危害的不断加重及昼夜高温伤害的累加性,迫切需要深入和系统地研究不同时期高温(白天高温和夜间高温)对水稻产量形成的影响及其内在机理。
根系是水稻吸收水分、养分和合成激素的重要器官,根系活力代表着根系对矿物质和水分吸收能力,具有重要的生物学意义[10-11],根系伤流强度能较为准确地反映根系活性变化情况[12]。陆定志等[13]研究发现,孕穗至抽穗期的伤流强度分别与单位面积的颖花数、结实率和千粒重呈显著正相关;抽穗至乳熟期间的伤流强度下降百分率与结实率、千粒重和产量呈极显著负相关。陶龙兴等[14]研究表明高温胁迫时根系的伤流强度呈先增加后降低的变化趋势,并且耐热品种伤流强度始终高于热敏感品种。伤流强度差异能在一定程度上反映高温下水稻产量差异,但关于不同时期高温下伤流强度与水稻产量的关系如何研究较为有限。
耐高温水稻品种的选育是应对全球气候变暖的主要策略。目前已经鉴定出部分优异的耐热水稻品种,如Nagina 22和汕优63。Nagina 22在开花期遭遇38℃的高温仍能保持较高的小穗育性(约80%)[15];以高产和适应性广闻名的超级杂交稻汕优63在温室或大田高温条件下均具有较高的结实率[16]。然而,这些水稻品种在花前生殖生长阶段的耐热性却鲜有报道。综合评估不同水稻品种产量及产量构成对不同时期高温的响应将有助于全面认识水稻的高温耐性。本研究利用人工气候室模拟高温处理,研究水稻产量及其构成对不同时期(穗分化期、开花期和灌浆期)不同类型高温(白天高温、夜间高温和全天高温)的响应;以更加全面认识水稻的高温耐受性,为耐热水稻品种的筛选与培育提供理论依据。
1 材料与方法
1.1 供试品种与种植方法
试验于2018年在南京农业大学丹阳试验站(东经119°10′,北纬34°36′)进行,供试品种为两个耐热水稻品种Nagina 22、汕优63和热敏感水稻品种两优培九。通过播期调整使三个水稻品种的穗分化期和开花期保持一致。两优培九、汕优63和Nagina 22的播种日期依次为4月26日、5月2日和5月26日,分别于5月25日、6月1日和6月20日选取长势一致的水稻幼苗移栽至桶中,桶高32 cm,内部直径30 cm,每桶装过筛土10 kg,每桶6穴,每穴1苗。各桶氮、磷、钾肥以N∶P2O5∶K2O=1∶0.5∶0.8的比例施用,肥料为复合肥(N∶P2O5∶K2O=1∶1∶1)、尿素和氯化钾。整个生育期每桶N的用量为2 g,肥料以1∶1的比例分基肥和穗肥两次施用。生长期间同常规栽培方式进行管理,及时拔除杂草,防治病虫害。
1.2 试验设计
试验设置四种温度处理:夜间高温(HNT:全天温度恒定在31℃);白天高温(HDT:07:00-19:00为38℃,19:00-07:00为24℃);全天高温(HDNT:07:00-19:00为38℃,19:00-07:00为31℃)和对照(CK,07:00-19:00为31℃,19:00-07:00为24℃)。所有植株在自然环境条件下生长,然后分别在穗分化期(幼穗长1~5mm)、开花期(始穗)和灌浆期(花后第7 d)移入人工气候室[气候室由南京全有电子科技有限公司设计建造,于2018年投入使用,单间规格为4.6 m×3.5 m×2.5 m,采用飞利浦LED植物生长灯补光,气源通过瓶装液态CO2供应,采用计算机自动控制系统精确控制温度(±0.5℃)、湿度(±10%)和二氧化碳浓度]。分别进行15 d、7 d和30 d的高温处理,室内湿度保持在70%左右,处理结束后搬出温室,于田间自然条件下生长至成熟。
1.3 测定项目及方法
高温处理结束的当天收集根系伤流液,伤流强度采用称重法[17]测定。从距茎基部8 cm处剪去地上部,将称重后的脱脂棉覆盖在基部茎秆剪切口,并用封口膜封口,于19:00-07:00收集12 h 后取下并称重,每处理收集9株。伤流液收集过程中不同温度处理下的植株仍置于对应温室内处理,收集结束后搬到室外自然条件下生长至成熟,采用差减法计算伤流量。脱脂棉收集前后的质量差即为伤流量,根系伤流强度表示为单位时间内单株渗出的伤流量。籽粒成熟后考种,考查单株穗数、每穗粒数、结实率、千粒重和单株产量,每处理每次考查3株,重复4次。
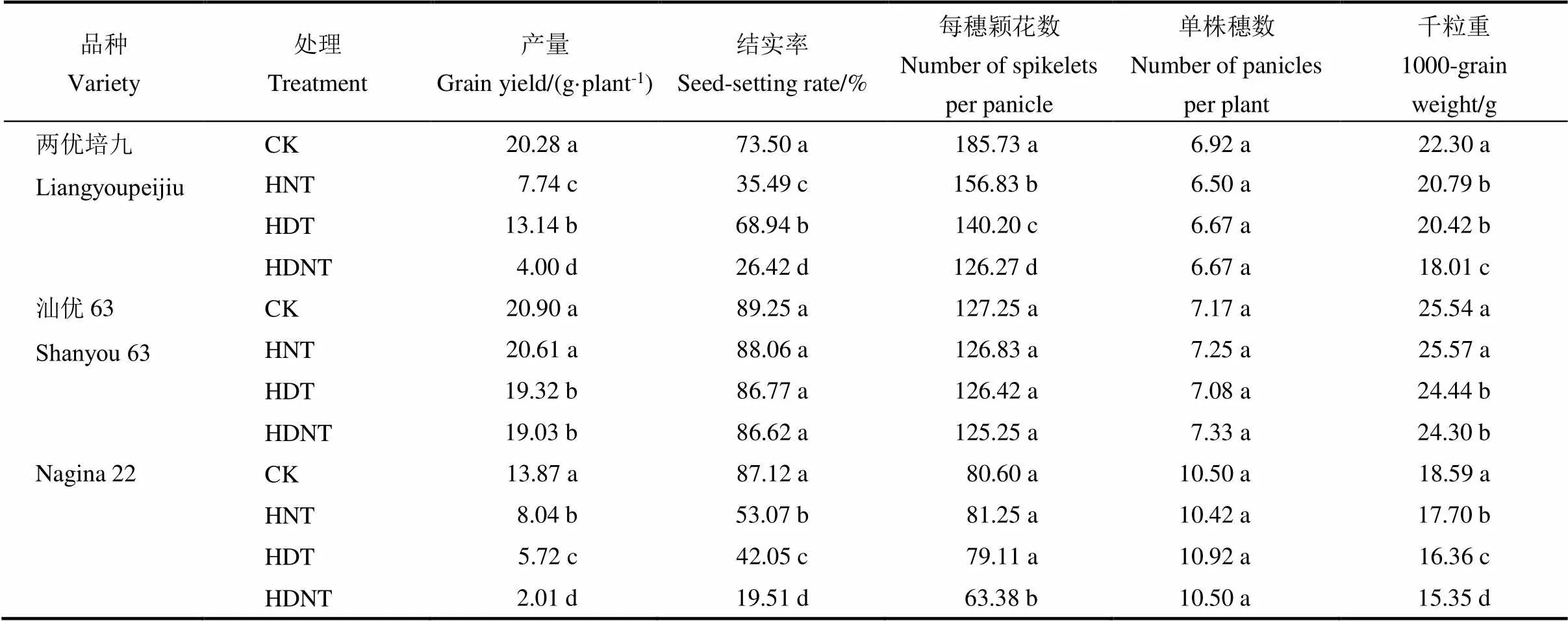
表1 产量及其构成对穗分化期高温的响应
CK-适温处理;HNT-夜间高温处理;HDT-白天高温处理;HDNT-全天高温处理;同一列同一品种中不同小写字母表示处理间差异显著(<0.05,=4)。
CK, Normal temperature; HNT, High night-time temperature; HDT, High daytime temperature; HDNT, High daytime and night-time temperature. Values followed by different letters within the same column for a cultivar are significantly different at< 0.05.
1.4 统计分析
采用Excel进行数据统计,R对不同温度处理下水稻产量和产量构成的平均值进行方差分析和多变量稳定性统计分析(GGE双标图分析),用于评估基因型×高温互作试验的相对产量和产量构成在穗分化期、开花期和灌浆期的表现,并根据产量和产量构成在高温处理下的稳定性对基因型进行排序。
2 结果与分析
2.1 产量和产量构成对穗分化期高温的响应
由表1可以看出,三个品种在穗分化期高温处理下籽粒产量平均降低42%。HNT、HDT和HDNT处理使三个品种的产量分别下降34.7%、33.3%和58%。穗分化期三种高温处理下籽粒减产表现出品种间差异:Nagina 22为61.3%,两优培九为59%,汕优63为5.7%。穗分化期高温处理使三个品种的结实率平均降低33.1%(Nagina 22:56.3%;汕优63:2.3%;两优培九:40.6%),颖花数减少10.5%,粒重降低8.5%;但高温处理对单株穗数影响不大。三个品种在穗分化期高温处理下,以Nagina 22的相对结实率最低,其次为两优培九和汕优63。两优培九的颖花数减幅最大(24%),其次是Nagina 22(6.7%)和汕优63(0.3%)。三种高温处理下粒重下降幅度最大的是Nagina 22(平均11.3%;HNT:5%;HDT:12%;HDNT:17%)和两优培九(平均11.3%;HNT:7%,HDT:8%,HDNT:19%),其次是汕优63(平均3%;HNT:0%;HDT:4%;HDNT:5%)。说明穗分化期高温对水稻产量的影响主要体现在结实率、颖花数和粒重的同步降低,两优培九和Nagina 22受穗分化期高温影响较大。
2.2 产量和产量构成对花期高温的响应
开花期高温处理显著降低水稻结实率(平均减少33.3%),但对颖花数、粒重和穗数影响不大。全天高温处理使三个水稻品种产量平均下降47.7% (17%~88%),其次是白天高温(平均29%;9%~53%)和夜间高温(平均24.3%;6%~56%)。花期高温处理对不同品种的影响差异较大,Nagina 22平均降低10.7% (6%~17%),汕优63平均降低24.7% (11%~38%),两优培九平均降低65.7% (56%~88%)。三个品种的结实率在全天高温下降低45.7% (15%~87%),白天高温下降低28.3% (9%~53%),夜间高温下降低了25.7% (9%~55%)。三种高温处理下,Nagina 22的结实率最高(平均89%;85%~91%),其次为汕优63(平均76.3%;65%~87%)和两优培九(平均35%;13%~47%)。
2.3 产量及其构成对灌浆期高温的响应
灌浆期高温处理使三个品种的籽粒产量平均下降16.6%,结实率降低12.7%。HNT、HDT和HDNT处理使三个品种的籽粒产量平均降低15.7% (4%~28%)、13.7% (7%~18%)和20.3% (12%~29%),结实率分别降低11.7%、11%和15.3%。籽粒灌浆期间的高温导致品种间产量及其构成变化的差异。三种高温处理下Nagina 22的产量平均下降11.3% (7.6%~15%),汕优63下降13.3% (4%~20%),两优培九下降25% (18%~29%)。高温处理使三个品种的结实率平均降低12.7%,但对粒重(降低4.3%)和穗数(增加1%)的影响较小,对颖花数没有影响(0%)。灌浆期高温处理下,Nagina 22保持了最高的结实率(平均93%;89%~96%),其次为汕优63(平均90.3%;83%~100%)和两优培九(平均78.7%;76%~83%)。说明灌浆期高温主要通过降低水稻粒重和结实率降低水稻产量,汕优63和Nagina 22受灌浆期高温影响较小与其结实率和粒重的相对稳定密切相关。
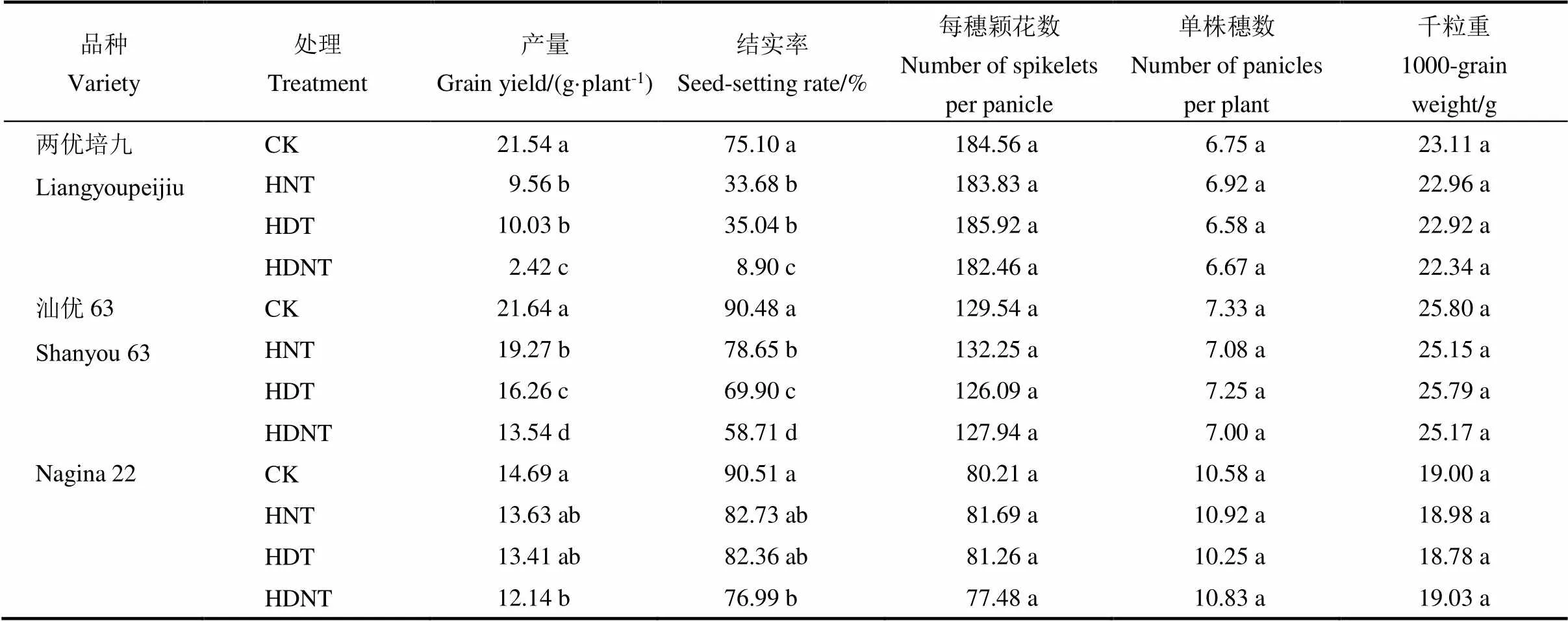
表2 产量及其构成对花期高温的响应
CK-适温处理;HNT-夜间高温处理;HDT-白天高温处理;HDNT-全天高温处理;同一列同一品种中不同小写字母表示处理间差异显著(<0.05,=4)。
CK, Normal temperature; HNT, High nighttime temperature; HDT, High daytime temperature; HDNT, High day- plus night-time temperature. Values followed by different letters within the same column for a cultivar are significantly different at< 0.05.

表3 产量及其构成对灌浆期高温的响应
CK-适温处理;HNT-夜间高温处理;HDT-白天高温处理;HDNT-全天高温处理;同一列同一品种中不同小写字母表示处理间差异显著(<0.05,=4)。
CK, Normal temperature; HNT, High nighttime temperature; HDT, High daytime temperature; HDNT, High day-plus night-time temperature. Values followed by different letters within the same column for a cultivar are significantly different at< 0.05.
2.4 不同时期高温下水稻产量稳定性差异比较
图1显示了水稻产量和稳定性的GGE双标图分析结果。双标图上三个水稻品种的投影(虚线)距离Y轴的平行距离表征其产量和产量构成因子。以坐标轴原点为参考,某品种的投影的位置在X轴上由负方向往正方向表示其相对产量增加,即投影在X轴正方向距离原点越远,其相对产量越高,投影在X轴负方向距离原点越远,其相对产量越低。结果显示,汕优63在三个时期高温胁迫下具有最高的相对产量,其次是Nagina 22和两优培九。每个品种的耐热性(稳定性)通过其在AEC Y轴上的投影(虚线)来估计,投影的绝对长度反应品种产量或产量构成因子的稳定性,某品种的产量或产量构成因子的投影绝对长度越短,表明该品种的产量或产量构成因子的稳定性越高;反之,投影绝对长度越长,则表明该品种稳定性越差。基于此分析,汕优63和两优培九在三个时期的平均产量均比Nagina 22稳定。不同水稻品种对不同时期高温的响应不同,高夜温下,穗分化期和灌浆期以汕优63的产量表现最好,开花期以Nagina 22的产量表现最好;高日温和全天高温下,穗分化期以汕优63的产量表现最好,开花期和灌浆期以Nagina 22的产量表现最好。综合而言,三个时期高温下,以汕优63的产量表现最好,具有高产稳产特性;其次是Nagina 22,相对高产但不稳产;两优培九的产量表现最差,表现出稳定的低产。值得注意的是,尽管两优培九的稳定性最高,但产量却是最低,其稳定性是建立在低产基础上的,因此是没有意义的。

HNT-夜间高温处理;HDT-白天高温处理;HDNT-全天高温处理;Pooled-三种温度下的平均值。
2.5 不同水稻品种的根系伤流强度差异性比较
高温处理对不同品种、不同时期根系伤流强度的影响均不相同。Nagina 22和两优培九在穗分化期和灌浆期高温处理结束后伤流强度显著降低,汕优63在三个时期处理后伤流强度均增加(平均增加13.9%)。穗分化期高温处理使Nagina 22和两优培九的伤流强度分别降低14%和18.3%,汕优63增加18%;花期高温处理下三个品种的伤流强度趋势表现一致,平均增加10.8% (Nagina 22、两优培九和汕优63分别增加12%、5.7%和14.7%)。灌浆期高温处理使Nagina 22和两优培九的伤流强度分别降低17%和26.7%,汕优63增加9.3%;整体上,全天高温对水稻伤流强度的影响最大,其次为白天高温和夜间高温,但灌浆期高温下两优培九的伤流强度对夜间高温较白天高温更加敏感。

表4 不同水稻品种根系伤流强度对高温的响应
CK-适温处理;HNT-夜间高温处理;HDT-白天高温处理;HDNT-全天高温处理;同一列同一品种中不同小写字母表示处理间差异显著(<0.05,=3)。
CK, Normal temperature; HNT, High nighttime temperature; HDT, High daytime temperature; HDNT, High day- plus night-time temperature. Values followed by different letters within the same column for a cultivar are significantly different at< 0.05.
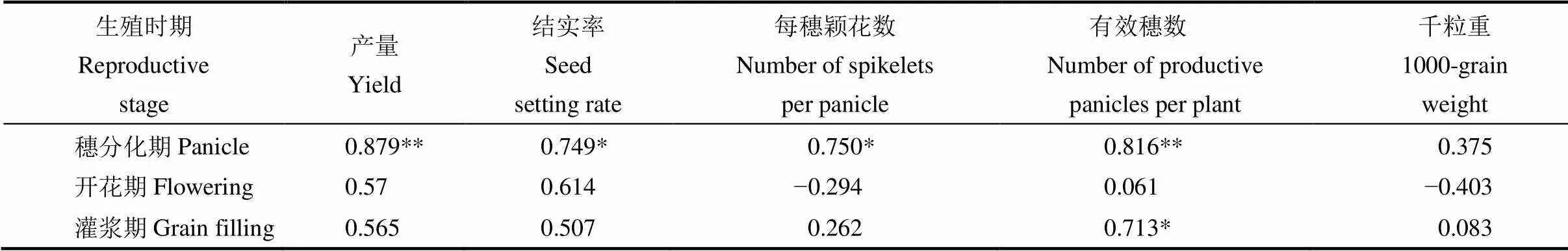
表5 不同时期高温下水稻伤流强度相对值与产量及产量构成因子相对值的相关性
*与**分别表示在0.05与0.01水平上显著相关。
*Significant correlation at the 0.05 probability level. **Significant correlation at the 0.01 probability level.
2.6 不同时期高温下水稻伤流强度与产量的相关性
对高温胁迫下水稻相对伤流强度与相对产量及其组分的相关性分析表明,穗分化期伤流强度与水稻产量、结实率、每穗颖花数和有效穗数呈显著和极显著正相关;开花期和灌浆期伤流强度与产量和结实率均呈正相关,但未达到显著水平。可见,水稻根部伤流强度变化状况与水稻产量及产量构成关系密切,尤其是穗分化期。
3 讨论
众多研究表明,不同时期高温对水稻产量的影响不同,表现出一定的品种间差异[18,19]。本研究发现,开花和灌浆期间高温胁迫下,三个品种中以Nagina 22的产量降幅最小,然而,穗分化期高温下Nagina 22的产量降幅最大(61.3%),表明Nagina 22在开花和灌浆期对高温胁迫具有较强的耐受性,但在穗分化期易受高温影响。
水稻的高温耐性由多种复杂因素决定,不同时期高温对水稻的影响具有一定的共性与特异性。如稳定的光合作用,高活性抗氧化系统等是水稻耐不同时期高温的共性之一[20-22],但由于水稻在不同时期完成特定的生命活动,其耐热机理也存在一定的差异性。从产量构成上看,穗分化期高温影响每穗颖花数、结实率和粒重,花期高温主要影响结实率,灌浆期高温则通过影响结实率和粒重影响水稻产量;从内在机理上,穗分化期高温耐性与穗器官发育密切相关[23],花期高温耐性受花药开裂、花粉萌发等因素影响[24],灌浆期高温耐性则与光合同化物转运密切相关[14]。可见,水稻对不同时期高温的响应机制并不具有一致性,对某一时期耐性强,不代表对另一时期耐性一定强。就Nagina 22而言,颖花开放持续时间较短,降低了其长时间接触高温胁迫的风险,花药开裂程度大、开裂速度快,使得花粉更易散落到柱头上是其耐花期高温的重要原因[25,26];穗分化期高温导致颖壳与维管束发育受阻,花器官发育不完全,颖花退化等降低了Nagina 22的产量[27]。本研究还发现穗分化期高温导致Nagina 22和两优培九穗茎节伸长受限,稻穗包裹在茎鞘中导致花器官畸形,造成颖花不育,这和前人研究结果一致[23, 27]。通过对根系伤流强度的研究,发现穗分化期Nagina 22的伤流强度显著降低,而伤流液中激素与穗器官发育密切相关,Wu等[27]研究表明高温下耐热品种通过稳定伤流速率向穗部输送更多的细胞分裂素和生长素以稳定小穗数。本研究中Nagina 22的耐热优势主要表现在生殖生长的中、后期,而生殖生长前期伤流强度下降较快、降幅较大,可能与Nagina 22不耐穗分化期高温有一定关系;汕优63在不同时期高温处理下均具有较高的伤流强度,维持一定穗粒数、结实率和粒重,表现出高产稳产特征;高温敏感品种两优培九在三个时期均具有最低的伤流强度和产量,表现出稳定的低产,相关性分析表明,生殖生长期高温下伤流强度与水稻产量呈正相关。高温下稳定的伤流强度有助于维持根源物质向地上部的运输以改善地上部生长,缓解高温损伤[14],但关于不同时期高温下伤流成分变化情况及其影响水稻产量及品质的内在机理还有待深入研究。值得注意的是,在本研究中,三个时期高温处理时长并不一致,无法排除由于处理时长不同引起的水稻产量差异,要精确评估不同时期高温下水稻产量,还需要更为细致的试验验证,但从本试验结果可以看出,水稻产量对高温的响应存在明显的品种特性和时期差异,各个时期所表现出来的耐热性/耐热机制并不具有相关性。
研究还发现,两优培九对夜间高温较为敏感,这在三个时期表现一致,在平均温度相同的条件下,高夜温要比高日温对水稻产量的影响更大,并且这种影响差异主要体现在结实率的变化上,三个时期夜间高温均比白天高温下结实率降幅更大。一般来说,白天高温以超过水稻耐受上限的温度直接影响水稻产量[28],而夜间高温通过影响水稻物质积累等过程影响产量[20],短期高温处理往往表现出夜间高温处理小于白天高温,但在本研究及相关研究中[29]表现出与此相反的趋势。目前还无法得知其内在原因,推测可能与品种类型有一定关系,三个品种中两优培九为大穗型品种,源库协调性可能更易受高温影响。庄宝华等[30]研究表明,两优培九的群体干物质向经济器官的分配率较低 ,结实期对源的削弱反应极为强烈,夜间高温下有限的营养物质可能优先供应给发育较早的颖花,这可能是其不耐夜间高温的原因之一。
播期的调整通过改变高温与水稻生育阶段的配置起到缓解高温危害的作用,是目前防御高温危害的重要途径之一,如通过推迟播期使高温易发时间与抽穗开花期错开以减轻高温危害。但随着播期的推迟,水稻穗分化期遭遇高温减产的概率大大增加。王亚梁等[26]在水稻花粉母细胞减数分裂期进行40℃高温处理(9 d)发现,相较适温,黄华占和丰两优6号颖花退化数目分别增加45.7%和81.7%,结实率降低95.2%,97.3%。在本研究中,穗分花期高温使水稻产量平均降低42%,远高于灌浆期16.6%。穗分化期高温通过降低小穗数、结实率、粒重等降低水稻产量,表现出一定的累加效应,产量构成中只要有一个因素对高温敏感即可能造成产量下降。可见,穗分花期高温对水稻产量的影响应引起足够重视。
耐热品种的选育是应对气候变化和不可预见性高温的有效措施,而多数研究只在特定时期(通常是在开花期间)评估水稻品种的耐热性[15,31]。Nagina 22由于对花期高温的极端耐受性被广泛用于耐热性生理和分子机理的研究,位于其第4染色体上的数量性状位点被认为是Nagina 22能在花期高温下保持较高结实率的原因之一[32]。杂交稻品种通常具有较高的产量潜力,但一般对高温敏感,通过分子育种方法,将Nagina 22或其他耐受性供体的耐热性(QTL)基因转移到杂交稻中,能使杂交稻在开花和灌浆期高温下保持相对稳定的产量[33]。但本研究表明,Nagina 22对穗分化期高温耐性较差,可能无法用于提高品种对穗分化期高温的耐性;而在三个时期高温胁迫下,两优培九的相对产量都很低,相反,汕优63无论是在穗分化期(80%)、开花期(72%)还是灌浆期(82%)高温胁迫下均能保持高产稳产,表现出对高温胁迫的综合耐受性,此外,Nagina 22综合性状较差,并未在生产上得到有效利用[15],而汕优63由于产量高、适应性广,被认为是我国杂交稻品种发展和生产的标志性品种[16],可作为耐高温品种选育的重要材料。综上,鉴于当前对水稻穗分化期高温的研究相对不足,而穗分花期高温对水稻产量的影响逐渐增大,有必要加强耐穗分化期高温资源的筛选与挖掘,并深入研究其遗传机理,选育对不同时期综合耐性较强的品种。
4 结论
水稻生殖生长阶段高温通过降低结实率、颖花数和粒重而导致产量损失,不同水稻品种的产量对不同时期、不同类型高温胁迫的响应不同。汕优63在整个生殖生长阶段对高温胁迫具有较强的综合耐受性,Nagina 22在生殖阶段中后期(开花期和灌浆期)具有较强的高温耐性,但对生殖阶段早期(穗分化期)高温高度敏感,两优培九对不同时期高温均较敏感。汕优63在不同时期高温下均表现出稳定的伤流强度,而高温敏感品种两优培九的伤流强度受高温影响显著下降。相关性分析表明,生殖生长期高温下伤流强度与水稻产量密切相关,水稻对不同时期的高温耐性并不具有相关性。为应对未来不可预见性的高温事件,今后培育耐高温品种应致力于提高穗分化期、开花期和灌浆期高温胁迫的综合耐受性。
[1] Pachauri R K, Meyer L A. Climate Change 2014: Synthesis Report. Contribution of working groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[C]. Geneva, Switzerland: IPCC, 2014: 1-151.
[2] 张桂莲, 张顺堂, 肖浪涛, 唐文帮, 肖应辉, 陈立云. 抽穗开花期高温胁迫对水稻花药、花粉粒及柱头生理特性的影响[J]. 中国水稻科学, 2014, 28(2): 155-166.
Zhang G L, Zhang S T, Xiao L T, Tang W B, Xiao Y H, Chen L YEffects of high temperature stress on physiological characteristics of anther, pollen and stigma of rice during heading-flowering stage[J]., 2014, 28(2): 155-166. (in Chinese with English abstract).
[3] Shen X, Liu B, Lu X, Fan G. Spatial and temporal changes in daily temperature extremes in China during 1960–2011[J]., 2016, 130: 1-11.
[4] 王亚梁, 张玉屏, 曾研华, 武辉, 向镜, 陈惠哲, 张义凯, 朱德峰. 水稻穗分化期高温对颖花分化及退化的影响[J]. 中国农业气象,2015, 36(6): 724-731.
WangY L, Zhang Y P, Zeng Y H,Wu H, Xiang J, Chen H Z, Zhang Y K, Zhu D F.Effect of high temperature stress on rice spikelet differentiation and degeneration during panicle initiation stage[J].2015, 36(6): 724-731. (in Chinese with English abstract).
[5] 杜尧东, 李键陵, 王华, 唐湘如, 胡飞. 高温胁迫对水稻剑叶光合和叶绿素荧光特征的影响[J]. 生态学杂志, 2012(10): 101-108.
Du Y D, Li J L, Wang H, Tang R, Hu F. Effects of high temperature stress on the flag leaf photosynthesis and chlorophyll fluorescence parameters of rice[J]., 2012(10): 101-108. (in Chinese with English abstract).
[6] Shi W, Yin X, Struik P C, Solis C, Xie F, Schmidt R C, Huang M, Zou Y, Ye C, Svk J. High day- and night-time temperatures affect grain growth dynamics in contrasting rice genotypes[J]., 2017, 68(18): 5233-5245.
[7] Xiong D L, Ling X X, Huang J L, Peng S B. Meta-analysis and dose-response analysis of high temperature effects on rice yield and quality[J]., 2017, 141: 1-9.
[8] Wu C, Cui K H, Wang W, Li Q, Fahad S, Hu Q, Huang J L, Nie L, Peng S B. Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice[J].2016, 6(1): 560-564.
[9] 董文军, 田云录, 张彬, 陈金, 张卫建. 非对称性增温对水稻品种南粳44米质及关键酶活性的影响[J]. 作物学报,2011, 37(5): 832-841.
Dong W J, Tian Y L, Zhang B, Chen J, Zhang W J. Effects of asymmetric warming on grain quality and related key enzymes activities for Japonica rice (Nanjing 44) under FATI facility[J]., 2011, 37(5): 832-841. (in Chinese with English abstract).
[10] 李婷婷, 冯钰枫, 朱安, 黄健, 汪浩, 李思宇, 刘昆, 彭如梦, 张宏路, 刘立军. 主要节水灌溉方式对水稻根系形态生理的影响[J]. 中国水稻科学,2019(4): 293-302.
Li T T, Feng Y F, Zhu A, Huang J, Wang H, Li S Y, Liu K, Peng R M, Zhang H L, Liu L J. Effects of main water saving irrigation methods on morphological and physiological traits of rice roots[J]., 2019(4): 293-302. (in Chinese with English abstract).
[11] Fahad S, Ihsan MZ, Khaliq A, Daur I, Saud S, Alzamanan S, Nasim W, Abdullah M, Khan IA, Chao W. Consequences of high temperature under changing climate optima for rice pollen characteristics-concepts and perspectives[J]., 2018, 10(1): 340-365.
[12] 沈波, 王熹. 籼粳亚种间杂交稻根系伤流强度的变化规律及其与叶片生理状况的相互关系[J]. 中国水稻科学,2000, 14(2): 122-124.
Shen B, Wang X. Changes of root exudate of indica-japonica hybrid rice and its relation to leaf physiological traits[J]., 2000, 14(2): 122-124. (in Chinese with English abstract).
[13] 陆定志. 杂交水稻根系生理优势及其与地上部性状的关联研究[J]. 中国水稻科学,1987,1(2): 81-94.
Lu D Z. Root physiological dominance of hybrid rice and its correlation with aboveground traits[J].1978, 1(2): 81-94. (in Chinese with English abstract).
[14] 陶龙兴,谈惠娟, 王熹,曹立勇,宋建,程式华. 开花和灌浆初期高温胁迫对国稻6号结实的生理影响[J]. 作物学报, 2009, 35(1): 110-117.
Tao L X, Tan H J, Wang X, Cao L Y, Song J, Cheng S H. Physiological effects of high temperature stress on grain setting for Guodao 6 during flowering and filling stage[J].2009, 35(1): 110-117. (in Chinese with English abstract) .
[15] Jagadish S V K, Craufurd P Q, Wheeler T R. Phenotyping parents of mapping populations of rice(L. )for heat tolerance during anthesis[J]., 2008, 48: 1140-1146.
[16] Xie F, Zhang J. Shanyou 63: An elite mega rice hybrid in China[J].2018, 11(1): 1-6.
[17] Zhi D, She T, Li G, Liu Z, Ding C, Lin C, Wang S, Ding Y. Application of nitrogen fertilizer at heading stage improves rice quality under elevated temperature during grain-filling stage[J]., 2017, 57(4): 2183-2192.
[18] 何洋,刘洋,方宝华,何小娥,杨坚,滕振宁,张玉烛.不同生育期温度逆境处理对早稻产量的影响[J].中国农学通报, 2016, 32(24): 18-24.
He Y, Liu Y, Fang B H, He X E, Yang J, Teng Z N, Zhang Y Z. Effect of temperature on early rice yield at different growth stages[J]., 2016, 32(24): 18-24. (in Chinese with English abstract).
[19] 曹云英, 段骅, 杨立年, 王志琴, 刘立军, 杨建昌. 抽穗和灌浆早期高温对耐热性不同籼稻品种产量的影响及其生理原因[J]. 作物学报,2009, 35(3): 512-521.
Cao Y Y, Duan Y, Yang L N, Wang Z Q, Liu L J, Yang J C. Effect of high temperature during heading and early grain filling on grain yield of Indica rice cultivars differing in heat-tolerance and its physiological mechanism[J].,2009, 35(3): 512-521. (in Chinese with English abstract).
[20] 杨军,蔡哲,刘丹,胡犁月,曲文波,张崇华,王尚明,田俊.高温下喷施水杨酸和磷酸二氢钾对中稻生理特征和产量的影响[J].应用生态学报, 2019, 30(12): 4202-4210.
Yang J, Cai Z, Liu D, Hu L Y, Qu W B, Zhang C H, Wang S M,Tian J. Effects of spraying salicylic acid and potassium dihydrogen phosphate on physiological characteristics and grain yield of single-season rice under high temperature condition[J]., 2019, 30(12): 4202-4210. (in Chinese with English abstract).
[21] 王艳, 高鹏, 黄敏, 陈浩, 杨志荣, 孙群. 高温对水稻开花期剑叶抗氧化酶活性及基因表达的影响[J]. 植物科学学报, 2015, 33(3): 355-361.
Wang Y, Gao P, Huang M, Chen H, Yang Z R, Sun Q. Effects of high temperature on the activity and expression of antioxidative enzymes in rice flag leaves during flowering stage.[J], 2015, 33(3): 355-361. (in Chinese with English abstract).
[22] 刘霞, 唐设, 窦志, 李刚华, 刘正辉, 王绍华, 丁承强, 丁艳锋. 茉莉酸甲酯对武运粳24和宁粳3号灌浆早期高温胁迫生理特性的影响[J]. 中国水稻科学, 2016, 30(3): 291-303.
Liu X, Tang S, Dou Z, Li G H, Liu Z H, Wang S H, Ding C Q, Ding Y F. Effects of MeJA on the physiological characteristics of japonica rice wuyunjing 24 and ningjing 3 during early grain filling stage under heat stress[J]., 2016, 30(3): 291-303. (in Chinese with English abstract).
[23] 王亚梁, 张玉屏, 朱德峰, 向镜, 陈惠哲, 张义凯. 水稻器官形态和干物质积累对穗分化不同时期高温的响应[J]. 中国水稻科学, 2016, 30(2): 161-169.
Wang Y L, Zhang Y P, Zhu D F, Xiang J, Chen H Z, Zhang Y K. Response of rice organ morphology and dry matter accumulation to high temperature at different panicle initiation stages[J]., 2016, 30(2): 161-169. (in Chinese with English abstract).
[24] 张祖建, 王晴晴, 郎有忠, 王春哥, 朱庆森, 杨建昌. 水稻抽穗期高温胁迫对不同品种受粉和受精作用的影响[J]. 作物学报, 2014, 40(2): 273-282.
Zhang Z J, Wang Q Q, Lang Y Z, Wang C G, Zhu Q S, Yang J C. Effects of high temperature stress at heading stage on pollination and fertilization of different types of rice variety.[J].,2014, 40(2): 273-282. (in Chinese with English abstract) .
[25] 穰中文, 周清明. 耐热水稻品种Nagina 22高温胁迫下的生理响应[J]. 植物遗传资源学报, 2012, 13(6): 1045-1049.
Rang Z W, Zhou Q M. Physiological responses of heat-tolerant variety Nagina 22 to high temperature stress in rice (L.)[J]., 2012, 13(6): 1045-1049. (in Chinese with English abstract) .
[26] Satake T , Yoshida S . High temperature-induced sterility in indica rices at flowering[J].,1978, 47(1): 6-17.
[27] Wu C, Cui K, Wang W, Li Q, Fahad S, Hu Q, Huang J, Nie L, Peng S. Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice[J].2016, 6: 34978.
[28] 黄福灯, 曹珍珍, 李春寿, 陆艳婷, 潘刚, 程方民. 花期高温对水稻花器官性状和结实的影响[J]. 核农学报, 2016, 30(3): 565-570.
Huang D F, Cao Z Z, Li C S, Lu Y T, Pan G, Ceng F M. Study on effects of high-temperature stress on rice in floral organ character and seed-setting rate.[J].(in Chinese with English abstract).
[29] 陈畅. 水稻生殖生长期不同时段高温对产量和稻米品质影响的研究[D]. 武汉: 华中农业大学, 2014.
Chen C. Effect of high temperature in different periods during rice reproductive stage on grain yield and rice quality.[D]. Wuhan: Huazhong Agricultural University, 2014.
[30] 庄宝华, 张书标, 章清杞, 杨仁崔. 两系亚种间杂交稻两优培九产量形成的生理特性[J]. 福建农业大学学报, 2003(2): 137-141.
Zhuang B H, Zhang S B, Zhang Q Q, Yang R C. Physiological characteristics of yield formation in Liangyoupeijiu, a two-line intersubspecific hybrid rice[J]., 2003, 32(2): 137-141. (in Chinese with English abstract).
[31] Prasad P V V, Boote K J, Allen L H, Sheehy J E , Thomas J M G. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress[J]., 2006, 95: 398-411.
[32] Ye C, Tenorio F A, Argayoso M A, Laza M A, Koh H J, Redoña E D, Jagadish K S, Gregorio G B. Identifying and confirming quantitative trait loci associated with heat tolerance at flowering stage in different rice populations[J]., 2015, 16(1): 41.
[33] Madan P, Jagadish S V, Craufurd P Q, Fitzgerald M, Lafarge T, Wheeler T R: Effect of elevated CO2and high temperature on seed-set and grain quality of rice[J]., 2012, 63(10): 3843-3852.
Differential Responses of Grain Yields to High Temperature in Different Stages of Reproductive Growth in Rice
SONG Youjin1, WU Chao2,*, LI Ziyu1, TANG She1, LI Ganghua1, WANG Shaohua1, DING Yanfeng1,*
(College of Agronomy,,,;,,,;;)
【】Our aim is to study the response of rice yield to different types of high temperature in different growth periods, and lays a basis for selection and breeding of heat-resistant rice varieties. 【】Heat-tolerant rice varieties Nagina 22, Shanyou 63 and heat-sensitive varieties Liangyoupeijiu were pot-cultured at high daytime temperature, high nighttime temperature, high day- plus night-time temperature, and normal temperature during panicle initiation, flowering, and grain filling in artificial climate rooms, so as to elucidate the response of rice yield and yield components to high temperature in different growth periods.【】Shanyou 63 had comprehensive tolerance in the three growth stages; Nagina 22 had strong tolerance to high temperature during flowering and filling, but was susceptible to high temperature during panicle initiation; Liangyoupeijiu was sensitive to high temperature during all reproductive growthstages.Shanyou 63 showed stable root bleeding intensity at different reproductive growth stages, while the root bleeding intensity of Liangyoupeijiu decreased significantly at high temperature.【】There is no significant correlation between high temperature tolerance of rice in different periods. Comprehensive tolerance to high temperature in different periods should be taken into consideration in selection or breeding of heat-resistant rice varieties.
rice (L.); grain yield; high temperature; reproductive stage;root bleeding intensity
10.16819/j.1001-7216.2021.0203
2020-02-18;
2020-06-29。
国家重点研发计划资助项目(2017YFD0300100);国家自然科学基金青年科学基金资助项目(32001471);江苏省自然科学基金资助项目(BK20180537)。
