淹水缓解直播早籼稻苗期低温冷害的生理特性研究
2021-03-17王文霞陈丽明王海霞刘有清吴自明曾勇军谭雪明潘晓华石庆华曾研华
王文霞 陈丽明 王海霞 刘有清 吴自明 曾勇军 谭雪明 潘晓华 石庆华 曾研华
淹水缓解直播早籼稻苗期低温冷害的生理特性研究
王文霞 陈丽明 王海霞 刘有清 吴自明 曾勇军 谭雪明 潘晓华 石庆华 曾研华*
(江西农业大学作物生理生态与遗传育种教育部重点实验室,南昌 330045;*通信联系人,E-mail: zyh74049501@ 163.com)
【】 探究淹水对低温胁迫下直播早籼稻幼苗生长的影响,为南方稻区直播稻生产与抗逆栽培奠定生理基础。以耐冷品种湘早籼6号和冷敏感品种中嘉早17为材料,设置低温处理(8℃)、低温淹水处理(8℃+ 淹水)与常温对照(25℃)3个处理(处理3 d),分析秧苗农艺性状、抗氧化酶活性、渗透调节物质含量、光合酶活性和内源激素含量等生理特性。与低温处理相比,低温淹水处理3 d后不同耐性品种的苗高、鲜质量、干质量和顶三叶叶长显著增加,根数、最大根长、剑叶叶长、倒2叶叶长也都有不同程度增加,淹水可减轻低温对水稻秧苗农艺性状的影响;耐冷品种湘早籼6号苗期受到伤害的程度小于冷敏感品种中嘉早17。此外,与低温处理相比,低温淹水处理显著降低叶片抗氧化酶活性(SOD、POD、CAT)、可溶性蛋白、渗透调节物质(MDA、脯氨酸)含量和内源生长抑制类激素(ABA)含量,同时显著增加了内源生长促进类激素(GA3)含量;低温条件下淹水可减少植物体内活性氧的积累,减轻膜脂过氧化,加强了植物体内源激素的调控作用。同时,低温和低温淹水处理显著降低叶片叶绿素和ATP含量,导致光合酶(Rubisco、PEPCK)活性降低;但低温淹水处理的影响低于低温处理,低温淹水可起到缓解作用。且在恢复处理后,低温淹水处理各生理活性指标更接近对照处理,而与冷敏感性品种相比,耐冷性品种可缓解低温胁迫产生的伤害。低温处理影响直播早籼稻秧苗生长特性,降低了秧苗光合酶活性;同时提升叶片抗氧化保护酶活性与渗透调节势,但淹水处理有助于缓解低温胁迫造成的叶片生理伤害。
早籼稻;直播稻;低温;淹水;生理特性
水稻是我国最重要的农作物,播种面积和总产量均居我国粮食作物之首。我国65%以上的人口以稻米为主食,因此,确保水稻高产、稳产对保障我国粮食安全意义重大[1]。近年来,随着城镇化、工业化的推进,农村青壮年劳动力大量向城市转移,农村劳动力资源短缺问题日益突出,劳动力成本越来越高,以省工、节本、高效为显著特征的水稻直播种植方式越来越受到农民特别是种粮大户的推崇[2]。随着全球气温上升、极端天气频繁出现,南方稻区直播早籼稻在播后常遭遇“倒春寒”,影响了直播稻产量的稳定性[3]。2010年4月,长江中下游双季早稻遭受“倒春寒”,导致直播稻出现大面积的烂种烂芽,严重影响直播稻生产[4]。同时,当温度低于10℃时,水稻的生理代谢过程也会受到影响,如膜透性增加、活性氧增加、光系统对光能的吸收转换与利用效率降低,直接影响水稻幼苗的生长,导致秧苗发黄,生长迟缓,甚至出现卷叶和死苗现象[5]。
水稻是喜湿耐湿作物,水分影响水稻生长发育过程中的蒸腾作用、光合作用和激素调节等生理生化过程[6],水分调节在水稻的生命活动中发挥着重要作用。近年来,直播水稻常遭遇淹水危害,不利于直播稻的稳产丰产。关于淹水对水稻生理生态特性的影响前人研究已有报道,但得出的结论不一[7-9],且不同的淹水深度对水稻生长的影响差异显著。宣守丽等[10]研究发现水稻分蘖期全淹处理后,绿叶、茎鞘生物量占地上部总生物量的比例出现不同程度的下降,黄叶比例增加,半淹处理对其影响并不明显。在水稻直播生产中,农民往往通过选用耐冷品种,以及田间采用灌溉深水层的管理措施来减少低温对幼苗的损害,从而提高秧苗成活率,降低直播水稻的生产风险。而如何利用水稻喜水耐淹的特性,缓解苗期低温胁迫已成为研究的热点。向镜等[11]研究表明,苗期低温淹水条件下胚芽鞘增长加快,株高显著增加,可有效降低死苗率。也有研究表明,水稻在苗期低温处理期间,湿润处理对水稻生长发育促进作用优于淹水处理[12]。目前已有的研究多集中于全淹胁迫,且大多注重分蘖期淹水,对直播早籼稻苗期低温淹水的响应规律研究较少[8, 13],特别是关于灌溉一定深水层缓解苗期低温冷害的秧苗生理机制尚不清楚。本研究以南方稻区早稻主栽品种为研究对象,采用盆栽试验,基于人工气候箱自动控温模拟早春低温气候条件,研究苗期淹水缓解低温对直播早稻幼苗生长的伤害,阐明不同耐冷性品种苗期低温冷害下生长差异的生理机制,为南方稻区直播稻生产与抗逆栽培提供理论依据。
1 材料与方法
1.1 试验材料
在前期试验基础上[7],供试品种选用冷敏感品种中嘉早17(由江西现代种业公司提供)和耐冷品种湘早籼6号(由湖南省沅江市农业科学研究所提供)。
1.2 试验设计
盆栽试验于2019年6月在江西农业大学作物生理生态与遗传育种教育部重点实验室组建的步入式人工气候室(气候室上下两层共32 m2)进行,设置低温、低温淹水及对照3个处理。低温(LT):8℃(昼夜温度为10℃/6℃);低温淹水(LTF):8℃(昼夜温度为10℃/6℃),保持5~6 cm的淹水层;对照(CK):25℃(昼夜温度为27℃/23℃)。各处理土壤保持湿润状态,光照时间均为12 h(7:00-19:00),光照强度为100 mol/(m2·s),人工气候室相对湿度为75%。选取刚破胸露白的种子进行直播,直播后置于25℃人工气候室中,待其生长到两叶一心后,选取三分之二的盆栽水稻幼苗移到8℃人工气候室进行低温和低温淹水处理3 d。在处理过程中,每个盆不定时地旋转,以避免光照对幼苗生长的影响。每处理重复3次,每重复设20盆。经过3 d的胁迫处理后,将植株置于25℃人工气候室恢复生长3 d。分别于低温处理前(6月29日),低温与低温淹水处理3 d(7月2日),恢复3 d后(7月5日)对水稻叶片取样,每个处理3次重复。除农艺性状调查样本外所有样品均在液氮中快速冷冻,保存于−80℃超低温冰箱中备用。
1.3 种植方法
盆栽用土取自江西农业大学科技园水稻试验田表层土壤(0-20 cm),试验用盆高15.0 cm、长25.0 cm、宽23.0 cm。试验土壤pH值为5.52,有机质含量为33.95 g/kg,全氮为2.03 g/kg,速效磷为24.78 mg/kg,速效钾为99.42 mg/kg。土壤自然风干,用FT-1000A土壤研磨机(常州市伟嘉仪器制造有限公司) 粉碎,然后用100 目筛子过筛。每盆装6 kg风干土,直播前浸泡两周,直播前1 d施复合肥3 g (N、P、K质量比为15∶15∶15),选取刚破胸露白的种子进行直播。
1.4 测定项目和方法
1.4.1 农艺性状
分别于处理前、处理3 d后和恢复处理3 d后取长势均匀的秧苗30株,每处理3次重复,将秧苗洗净擦干后测量鲜质量、苗高、顶三叶长度,记录每株秧苗的根数和最大根长,然后将秧苗分别装于网袋,置于烘箱中80℃下烘干至恒重后称干质量。
1.4.2 抗氧化酶活性及可溶性蛋白含量
超氧化物歧化酶(SOD)、过氧化物酶(POD) 及过氧化氢酶(CAT) 活性测定均采用李合生等[14]的方法进行。称取0.1 g叶片组织,加入1 mL提取液(磷酸缓冲液),进行冰浴匀浆,8000 r/min、4℃下离心10 min后,取上清液置于冰上待测。可溶性蛋白含量利用考马斯亮蓝比色法进行测定[14]。
1.4.3 叶片膜脂过氧化物
丙二醛含量采用硫代巴比妥酸法进行测定。称取0.1 g组织,加入1 mL提取液(磷酸缓冲液),进行冰浴匀浆,8000 r/min、4℃下离心 10 min,取上清液,置于冰上待测。采用茚三酮比色法来测定游离脯氨酸的含量[15],取0.1 g 组织,加入1 mL 提取液(磺基水杨酸溶液),进行冰浴匀浆,之后置于 90℃下振荡提取 10 min,常温离心(10 000 r/min, 10 min)后,取上清液,置冰上待测。
1.4.4 光合酶活性和ATP含量
用80%丙酮溶液提取叶绿素,采用分光光度法测定叶绿素含量[14]。采用苏州科铭生物技术有限公司生产的相关试剂盒对ATP含量、磷酸烯醇式丙酮酸羧激酶(PEPCK)和核酮糖-1,5-二磷酸羧化酶(Rubisco)活性进行测定[16, 17]。
1.4.5 内源激素含量测定
采用高效液相色谱法 (HPLC) 测定脱落酸(ABA)和赤霉素(GA3)内源激素含量[18]。测定方法略作调整:称取新鲜样品0.1 g,放入研钵中磨碎,加入1 mL试剂Ⅰ(80 mL甲醇+20 mL水+1 mL乙酸),4℃下浸提过夜。8000×下离心10 min,离心后取出上清液,氮吹至水相。用试剂Ⅱ(柠檬酸)调pH值至2~3,用乙酸乙酯萃取3次,合并乙酸乙酯相,氮吹至干。甲醇定容至0.5 mL,针头式过滤器过滤于带有内衬管的样品瓶内待测。
1.5 数据分析
采用Microsoft Office Excel 2010和DPS 9.5软件进行数据处理和差异显著性分析。
2 结果与分析
2.1 淹水对遭受苗期低温冷害直播早籼稻农艺性状的影响
由表1可知,与对照相比,低温和低温淹水处理3 d后不同耐冷性品种的苗高显著降低,冷敏感品种中嘉早17的降幅为33.5%、30.0%,耐冷品种湘早籼6号的降幅为30.1%、26.9%,低温淹水处理的苗高降幅小于低温处理,且差异达显著水平,而冷敏感品种的降幅高于耐冷品种;然而,与低温处理相比,低温淹水处理3 d后不同耐冷性品种的鲜质量、干质量和顶三叶长度显著增加,根数、最大根长、顶一叶长、顶二叶长也呈增加趋势。说明低温胁迫处理影响植株的正常生长发育,而淹水可缓解低温对水稻秧苗的影响,且不同耐冷性品种间存在差异,耐冷性品种湘早籼6号受到伤害小于冷敏感品种中嘉早17。同时恢复3 d后,低温淹水处理的秧苗各性状指标恢复要优于低温处理。

表1 淹水对苗期遭受低温冷害直播早籼稻农艺性状的影响
Pre-T-处理前;LT-低温处理3 d;LTF-低温淹水处理3 d;CK-处理3 d后的常温对照;Re-LT-低温处理恢复3 d;Re-LTF-低温淹水处理恢复3 d;Re-CK-恢复3 d后的对照;T1-顶一叶;T2-顶二叶;T3-顶三叶。同列数据后不同小写字母表示差异达显著水平(< 0. 05)。
XZX6, Xiangzaoxian 6; ZJZ17, Zhongjiazao 17; Pre-LT, Before treatment; LT, Low temperature treatment for 3 days; LTF, Low temperature and flooding treatment for 3 days; CK, Control; Re-LT, Low temperature treatment followed by 3 days of recovery; Re-LTF, Low temperature flooding treatment followed by 3 days of recovery; Re-CK, Control after 3 days of recovery; T1, The first leaf from top; T2, The 2nd leaf from top; T3, The 3rd leaf from top. Different lowercase letters in the same column mean significant difference at 0.05 level.
2.2 淹水对苗期遭受低温冷害直播早籼稻抗氧化酶活性及可溶性蛋白含量的影响
当植物受到低温胁迫时,过量的氧化自由基在体内迅速积累,严重损伤细胞的正常生理活动。水稻中的抗氧化酶被激活,保护植株免受严重伤害。与对照相比,低温处理3 d后,不同耐冷性品种的抗氧化酶活性(SOD、POD、CAT)和可溶性蛋白含量显著增加 (<0.05),POD和CAT活性在各处理间差异达显著水平。与低温相比,低温淹水处理的SOD活性显著降低,耐冷品种湘早籼6号降幅为12.6%,冷敏感品种中嘉早17降幅为16.2%,中嘉早17的降幅大于湘早籼6号。恢复处理3 d后,不同耐性品种的抗氧化酶(SOD、POD、CAT)活性和可溶性蛋白含量呈降低趋势,趋近于对照,且低温淹水与对照水平相近。说明低温胁迫后幼苗应对低温冷害机制启动,细胞膜受到严重伤害,但淹水可减缓低温对幼苗的损伤。
2.3 淹水对遭受苗期低温冷害直播早籼稻叶片膜脂过氧化物含量的影响
由图2可知,处理3 d后,低温处理水稻的MDA含量和游离脯氨酸含量较对照和低温淹水处理显著增加,耐冷品种湘早籼6号的低温淹水处理与对照无显著性差异,而冷敏感品种中嘉早17的MDA和游离脯氨酸含量在各处理间差异达显著水平。恢复处理3 d后,不同耐性品种低温和低温淹水处理的MDA含量和游离脯氨酸含量呈降低趋势,但相比低温淹水,低温处理下增加显著,且冷敏感品种增幅明显,恢复处理后中嘉早17 MDA各处理差异显著。表明低温淹水可降低直播早籼稻苗期低温冷害后叶片的膜脂过氧化物,减轻叶片细胞渗透调节物质伤害。
2.4 淹水对遭受苗期低温冷害直播早籼稻光合酶活性及ATP含量的影响
低温冷害胁迫对植物光合作用、叶绿素合成和能量代谢有重要影响。核酮糖-1,5-二磷酸羧化酶(Rubisco)是光合作用中决定碳同化速率的关键酶,与光合速率呈正相关;磷酸烯醇式丙酮酸羧激酶(PEPCK)是糖酵解过程中重要的代谢中间产物,在能量代谢中起着关键作用。ATP作为可被生物直接利用的细胞内能量传递的“分子通货”,是表征生命体活力最直接的因子,可反映植物生理过程的能量代谢水平。由图3可知,处理3 d后,与对照处理相比,低温和低温淹水处理的Rubisco活性、PEPCK活性、叶绿素和ATP含量均呈降低趋势,其中,对照处理与低温处理差异达显著水平,低温淹水处理降幅低于LT处理;此外,相比低温处理,低温淹水增加了不同耐性品种各指标含量,其中PEPCK活性和ATP含量差异显著。恢复3 d后,Rubisco活性、PEPCK活性、叶绿素和ATP含量较处理期间增加,且低温淹水显著高于低温处理,趋近于对照处理。说明淹水可缓解低温胁迫对直播早稻秧苗叶片光合酶活性伤害的影响,且恢复效应优于低温胁迫处理。
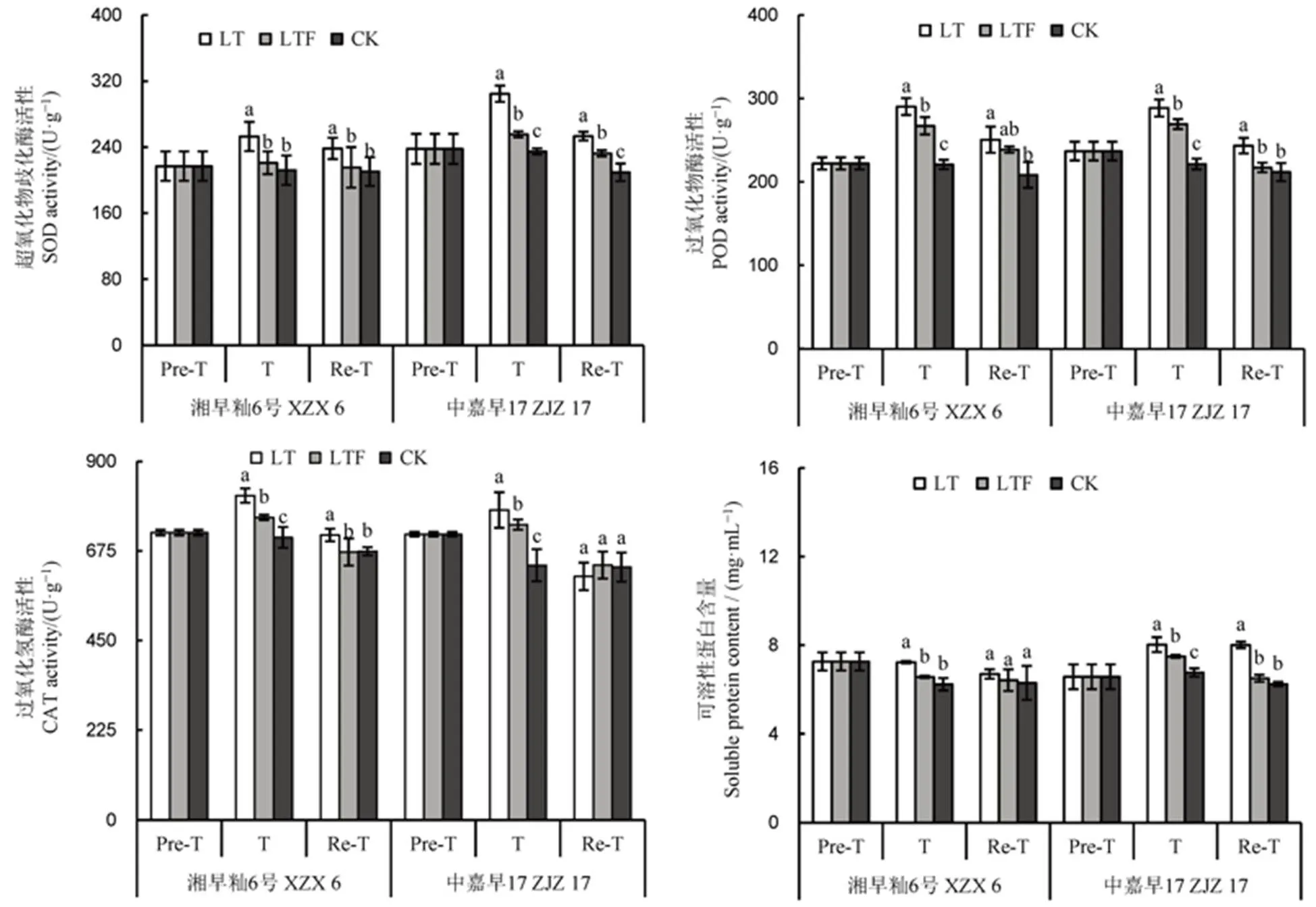
Pre-T-处理前;T-处理3 d后;Re-T-恢复3 d后;LT-低温;LTF-低温淹水;CK-对照。柱上不同小写字母表示在5%水平上差异显著。下同。

图2 淹水对苗期遭受低温冷害直播早籼稻叶片膜脂过氧化物含量的影响
2.5 淹水对苗期低温冷害直播早籼稻内源激素含量的影响
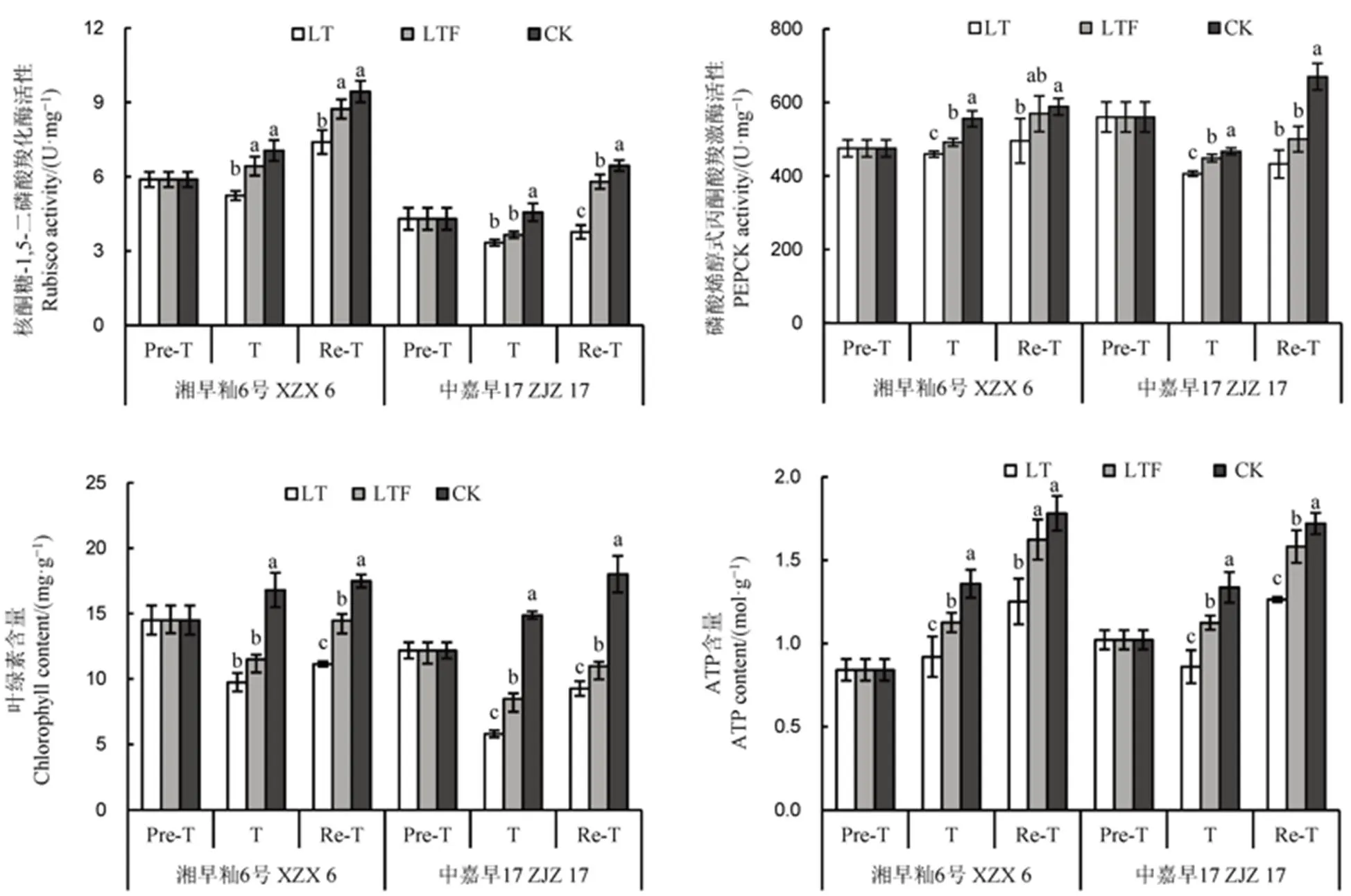
图3 淹水对苗期遭受低温冷害直播早籼稻光合酶活性及ATP含量的影响
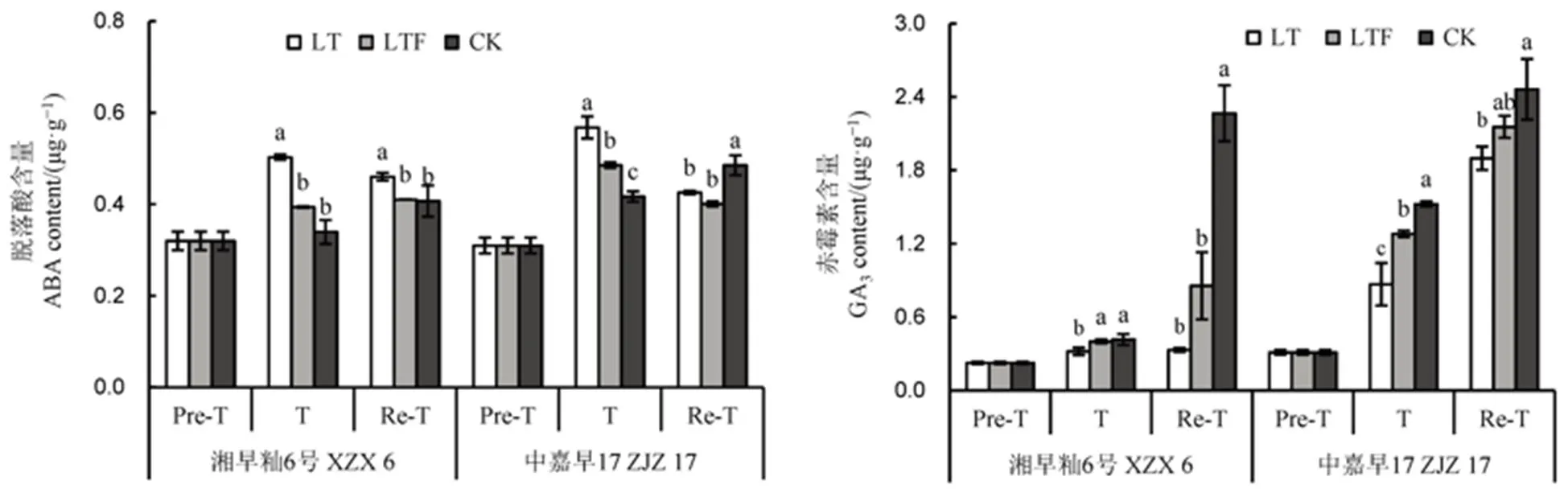
图4 淹水对苗期遭受低温冷害直播早籼稻内源激素含量的影响
由图4可知,处理3 d后,低温处理下各品种ABA含量较对照和低温淹水处理显著增加,其中冷敏感品种中嘉早17的ABA含量对照处理显著低于低温淹水处理,而耐冷品种湘早籼6号对照和低温淹水处理间差异不显著;低温和低温淹水处理各耐性品种GA3含量均低于对照,低温与对照差异显著,而中嘉早17各处理间差异达显著水平,且低温显著低于低温淹水、对照。恢复3 d后,ABA含量较低温处理3 d后有所降低,但低温淹水低于低温;而GA3含量呈增加趋势,各处理表现为低温<低温淹水<对照,且低温与对照差异显著。表明低温冷害后内源生长促进类激素(GA3)显著下降,而内源生长抑制类激素(ABA)显著升高,而淹水可起到对低温冷害的缓解作用。
3 讨论
3.1 淹水对苗期低温冷害直播早籼稻农艺性状的影响
低温冷害造成水稻幼苗形态指标上的变化最为直观,如苗高增长速度降低、叶片卷曲、根数减少等[19]。根数、最大根长、叶长等均可作为水稻耐冷性鉴定指标[20, 21]。吴立群等[22]研究发现,低温处理后,不同耐性品种幼苗的地上部各形态指标和根系形态指标均有一定程度的降低,但其降幅存在差异。6℃低温胁迫下水稻幼苗生长严重受阻,部分籼稻幼苗生长停止,根系活力显著降低[23]。在水稻生产中,适度淹水可以起到增温保湿作用,减轻低温对早稻幼苗的直接损害,从而减少水稻生产中的损失[24, 25]。本研究结果表明,低温胁迫影响了植株的正常生长发育,与低温相比,低温淹水处理3 d后不同耐性品种的苗高、鲜质量、干质量和倒三叶叶长显著增加,淹水可减轻低温对直播早稻秧苗农艺性状的影响,起到深水护苗的作用。这与前人关于低温及淹水处理对水稻生长规律研究结果类似[19, 26]。低温胁迫主要通过减小秧苗叶片的栅栏组织厚度,造成植株生物量减少,从而影响植株器官的形态建成、物质合成与积累,而淹水营造的缺氧环境诱导植株体内乙烯含量增加,刺激了细胞的分裂和伸长,从而促进水稻株高的增长,使得水稻在低温胁迫下表现出较好的适应性[27]。此外,低温和低温淹水后苗高均出现显著降低的现象,这可能是由于人工气候室的光照及密闭的环境与大田环境不同,加速了秧苗苗高的快速增长,使得秧苗变得细长,而低温和低温淹水处理期间由于夜间温度较低,造成秧苗叶片失水干枯卷曲,导致秧苗矮小。耐冷品种湘早籼6号秧苗农艺性状表现要好于冷敏感品种中嘉早17,这可能与品种的遗传特性有关;同时低温抑制了冷敏感品种叶片和节间伸长。
3.2 淹水对苗期低温冷害直播早籼稻抗氧化酶系统的影响
植物受低温胁迫时,细胞内氧代谢的平衡被破坏,导致活性氧(ROS)的产生,加剧膜脂过氧化作用,造成细胞膜系统损伤[28]。为了应对低温冷害对其生长造成的影响,水稻启动自身应对低温冷害的保护机制以减少低温对水稻的损伤[28]。抗氧化酶是植物活性氧清除系统中的重要酶类,可以平衡活性氧自由基的产生与清除[29, 30]。而适度淹水可进一步增强植物的抗氧化能力,抑制膜脂的过氧化作用,减轻叶片过氧化损伤[31]。本研究表明,低温和低温淹水处理3 d后,直播早籼稻幼苗叶片的SOD、POD、CAT活性和可溶性蛋白含量增加,而低温淹水后POD和CAT的增幅要显著低于低温处理,低温淹水受到的冷害程度明显小于低温处理,这与Minami等[32]研究结果一致。浅层淹水可以改善直播稻幼苗在低温胁迫下的抗氧化酶保护系统,不同耐冷性品种间存在差异。与低温相比,低温淹水处理的SOD活性均显著降低,但冷敏感品种中嘉早17的降幅大于耐冷品种湘早籼6号,表明耐冷品种湘早籼6号受低温胁迫影响小,抵抗低温和淹水的能力较强,在生产中可作为直播稻抗逆栽培的首选品种。
此外,当细胞面临低温胁迫时,需要进行渗透调节以维持细胞内外的渗透平衡,因此必然会使机体内参与渗透调节的物质发生变化,从而避免冻害,提高植物的抗寒能力[33]。MDA含量可在一定程度上反映植物细胞膜脂过氧化水平和对细胞膜结构的伤害程度[34]。本研究表明,低温淹水3 d后,MDA和脯氨酸含量较低温处理显著降低,表明低温引起了植物体内活性氧的积累和膜脂过氧化作用的加剧,而低温淹水有助于减少膜脂过氧化酶在膜脂过氧化中的参与,减少对细胞膜系统的损伤,降低细胞内渗透压力。同时,低温淹水处理的可溶性蛋白含量显著低于低温处理,进一步说明淹水可减少低温胁迫对叶片生理活性的损伤,保持叶片生理结构的完整性。
3.3 淹水对直播早籼稻苗期低温冷害光合作用和内源激素的影响
水稻光合作用和呼吸作用对低温十分敏感,在营养生长期,低温可抑制叶面积的形成,降低冠层的光合作用能力,使干物质积累量和产量减少[35]。Rubisco和PEPCK是植物光合作用中的一个关键酶,既控制着CO2的固定,同时又制约着碳素向Calvin循环和光呼吸循环分流,其活性直接影响着光合速率[36]。Sperotto等[37]研究发现,淹水胁迫降低水稻光合速率、减弱蒸腾作用,从而使得产量下降。本研究结果也表明,低温和低温淹水处理降低光合酶(Rubisco、PEPCK)活性,同时降低叶绿素和ATP含量,而低温淹水处理降幅小于低温处理,说明淹水可缓解低温对水稻光合作用的影响。低温胁迫使类囊体膜上PSⅡ的光能传递效率和光能转换效率降低,从而导致CO2同化能力降低,同时使得Rubisco和PEPCK酶活性降低[38]。而浅层淹水条件下仍有部分叶片露出水面,露出的叶片可以正常进行光合作用,同时根部可以通过其发达的通气系统获得部分氧气,其积累的光合产物可用作抵御低温及短期淹水的能量消耗[39]。耐冷性品种的自身反应机制有利于抵抗低温胁迫对叶片光合能量和光合作用造成的伤害。
在低温胁迫下,水稻还通过改变体内激素含量来调节生理过程。有研究指出,在低温处理下水稻叶片ABA含量总体上升,GA3含量下降[40]。本研究也得出类似的结果。低温胁迫下,细胞中内源生长促进类激素GA3含量降低,抑制水稻植株生长,促进气孔关闭,降低蒸腾作用以适应低温环境[41];同时,细胞中ABA迅速积累,低温增加了叶绿体膜对ABA的通透性,并加快由根系合成的ABA向叶片运输及积累[23]。刘春风等[42]研究表明,随着淹水时间的延长,银杏、黄连叶片的ABA含量呈下降趋势。本研究表明,相比低温处理,低温淹水处理3 d后ABA含量显著降低,而GA3含量显著增加。说明淹水处理对低温冷害下叶片内源激素调节具有较好的缓解恢复效应,这与刘春风等[42]的研究结果较为相似。但潘向艳等[43]研究结果表明,淹水4 d内,叶片体内ABA含量先升后降,IAA含量先降后升。这表明不同植物对淹水胁迫的响应存在差别,尤其在激素水平动态平衡层面,其抵御胁迫的内在机制尚需进一步研究。
3.4 淹水缓解直播早籼稻苗期低温冷害途径
一般来说,强抗性的优质品种、适宜的播期及合理的水分管理等方法是直播稻抗逆稳产栽培的重要措施。本研究湘早籼6号对低温冷害淹水的敏感性要低于中嘉早17,说明其可作为耐淹耐寒性的品种直播。在直播稻生产中,当低温来临时,农户往往通过灌溉一定的水层来减轻低温对秧苗的伤害,从而达到深水护苗的作用。我们前期通过田间试验观察发现,适度淹水可防止早籼稻幼苗苗尖因气候因素造成生理失水;而两叶一心期的幼苗缺水若再遭遇低温伤害更严重,会导致苗黄干尖,叶片卷曲。本研究表明,直播早籼稻幼苗在两叶一心期保持5~6 cm淹水层3 d可减轻低温冷害对不同耐冷性品种苗高、鲜质量、根长等农艺性状的伤害。因为适当的淹水有利于提高植株节间基部及根系温度,防止夜间低温对早籼稻幼苗的直接伤害,且缺氧环境刺激了细胞的分裂和伸长,促进了水稻苗高的生长[24, 25]。同时,叶片生理活性结果表明,淹水可缓解低温条件下植物体内活性氧物质的积累和膜脂过氧化作用的加剧,改善了植物体内源激素的积累。因此,直播稻生产时,在选用耐寒耐淹品种的基础上,可通过采用深水护苗方法,减轻低温对幼苗造成的损伤;且播种前需做到精细化整地,减少因淹水后水层深浅不一形成淹涝冷害的双重胁迫,造成烂秧烂苗的后果。此外,通过适当增施氮肥[44]、磷肥[45]也可起到对低温冷害的缓解作用。
4 结论
相比低温处理,低温淹水处理3d后显著增加不同耐性品种的苗高、鲜质量、干质量和顶三叶叶长,根数、根长、顶一叶叶长、顶二叶叶长均不同程度的增加,淹水可缓解低温冷害胁迫对直播早籼稻秧苗农艺性状的伤害,冷敏感品种中嘉早17较耐冷品种湘早籼6号受到的伤害程度大;同时,低温淹水处理显著降低了不同耐性品种的抗氧化酶活性(SOD、POD、CAT)、可溶性蛋白、渗透调节物质(MDA、Pro含量)和ABA含量,但显著增加GA3含量;此外,低温和低温淹水处理降低不同耐性品种叶片的叶绿素和ATP含量,导致光合酶(Rubisco、PEPCK)活性降低,但低温淹水的降幅低于低温处理。总体而言,浅层淹水有助于缓解低温胁迫对直播早籼稻秧苗造成的有关生理伤害。
[1] 李刚华. 特高产水稻产量形成机理及定量栽培技术研究[D]. 南京: 南京农业大学, 2010.
Li G H. Study on yield formation mechanism and quantitative cultivation technology of special high-yield rice[D]. Nanjing: Nanjing Agricultural University, 2010. (in Chinese with English abstract)
[2] Ohno H, Banayo N P M C, Bueno C, Kashiwagi J I, Nakashima T, Lwama K, Corales A M, Garcia R, Kato Y. On-farm assessment of a new early-maturing drought-tolerant rice cultivar for dry direct seeding in rainfed lowlands[J]., 2018, 219: 222-228.
[3] 吴杏春, 王茵, 林文雄. 水稻苗期耐冷性状的QTL分析[J]. 中国生态农业学报, 2008, 16(4): 1067-1069.
Wu X C, Wang Y, Lin W X. QTL mapping in controlling seedling cold tolerance in rice[J]., 2008, 16(4): 1067-1069. (in Chinese with English abstract)
[4] 王慰亲. 种子引发促进直播早稻低温胁迫下萌发出苗的机理研究[D]. 武汉: 华中农业大学, 2019.
Wang W Q. Mechanisms underlying the effects of seed priming on the establishment of direct-seeded early season rice under chilling stress[D]. Wuhan: Huazhong Agricultural University, 2019. (in Chinese with English abstract)
[5] 王亚男, 范思静. 低温胁迫对水稻幼苗叶片生理生化特性的影响[J]. 安徽农业科学, 2017, 45(5): 8-9.
Wang Y N, Fang S J. Effects of low-temperature stress on the physiological and biochemical characteristics of rice seedling leaves[J]., 2017, 45(5): 8-9. (in Chinese with English abstract)
[6] 易子豪. 水分亏缺对水稻秧苗生长的影响及调控[D]. 北京: 中国农业科学院, 2020.
Yi Z H. Effect of water deficit on rice seedling growth and its regulation[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020. (in Chinese with English abstract)
[7] 唐双勤. 芽期低温、淹水胁迫对早籼稻生长特性与产量的影响[D]. 南昌: 江西农业大学, 2019.
Tang S Q. Effects of low temperature and water-logging stress at bud stage on growth characteristics and yield of early indica rice[D]. Nanchang: Jiangxi Agricultural University, 2019. (in Chinese with English abstract)
[8] Singh A, Septiningsih E M, Balyan H S, Singh N K, Rai V. Genetics, physiological mechanisms and breeding of flood-tolerant rice (L.)[J]., 2017, 58(2): 185-197.
[9] 张耗, 余超, 陈可伟, 孔祥胜, 刘海浪, 陈俊义, 顾骏飞, 刘立军, 王志琴, 杨建昌. 直播方式对水稻生理性状和产量的影响及其成本分析[J]. 农业工程学报, 2017, 33(13): 58-64.
Zhang H, Yu C, Chen K W, Kong X S ,Liu H L, Chen J Y, Gu J F , Liu L J, Wang Z Q, Yang J C. Effect of direct-seeding methods on physiological characteristics and grain yield of rice and its cost analysis[J]., 2017, 33(13): 58-64. (in Chinese with English abstract)
[10] 宣守丽, 石春林, 张建华, 魏秀芳, 曹宏鑫, 薛昌颖. 分蘖期淹水胁迫对水稻地上部物质分配及产量构成的影响[J]. 江苏农业学报, 2013, 29(6): 1199-1204.
Xuan S L, Shi C L, Zhang J H, Wei X F, Cao H X, Xue C Y. Effects of submergence stress on aboveground matter distribution and yield components of rice at tillering stage[J]., 2013, 29(6): 1199-1204. (in Chinese with English abstract)
[11] 向镜, 陈惠哲, 张玉屏, 张义凯, 朱德峰. 淹涝条件下水温对水稻幼苗形态和生理的影响[J]. 中国水稻科学, 2016, 30(5): 525-531.
Xiang J, Chen H Z, Zhang Y P, Zhang Y K, Zhu D F. Effects of on morphological and physiological response of rice seedlings to water temperature under complete submergence[J]., 2016, 30(5): 525-531. (in Chinese with English abstract)
[12] 杨知建, 成诗瑜. 低温下土壤水分对籼稻及粳稻根系生长的影响[J]. 湖南农业大学学报, 2002, 28(5): 369-372.
Yang Z J, Cheng S Y. Effects of the soil moisture at low temperature on the growth roots[J]., 2002, 28(5): 369-372. (in Chinese with English abstract)
[13] 杜彦修, 孙红正, 张静, 李俊周, 彭廷, 赵全志. 不同生育时期淹水对水稻耐淹基因导入系产量的影响[J]. 中国稻米, 2016, 22(4): 28-34.
Du Y X, Sui H Z, Zhang J, Li J Z, Peng T, Zhao Q Z. Effects of waterflooding on yield of rice submergence tolerance geneintrogression line at different growth stage[J]., 2016, 22(4): 28-34. (in Chinese with English abstract)
[14] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2006.
Li H S. Principle and Technology of plant physiology and biochemistry experiment[M]. Beijing: Higher Education Press, 2006. (in Chinese)
[15] 张志良. 植物生理学实验指导[M]. 北京: 高等教育出版社, 1990.
Zhang Z L. Experimental Guidance on Plant Physiology[M]. Beijing: Higher Education Press, 1990. (in Chinese)
[16] Chen G, Zhang Y, Ruan B P, Guo L B, Zeng D L, Gao Z Y, Zhu L, Hu J, Ren D Y, Yu L, Xu G H, Qian Q.controls the vegetative growth and panicle fertility of rice by its effect on potassium-mediated sugar metabolism[J]., 2018, 274: 261-270.
[17] Wu K, Li J X, Luo J P, Liu Y K, Song Y C, Liu N L, Rafiq M T, Li T Q. Effects of elevated CO2and endophytic bacterium on photosynthetic characteristics and cadmium accumulation in[J]., 2018, 643: 357-366.
[18] Teng Y B, Cui H Q, Wang M Y, Liu X Y. Nitrate reductase is regulated by circadian clock-associated1 in[J]., 2017, 416(1): 1-9.
[19] 周伟江, 吴旺嫔, 唐才宝, 肖志芳, 陈光辉, 王悦. 外源油菜素内酯对低温胁迫下水稻幼苗生长及生理特性的影响[J]. 西北农业学报, 2020, 29(9): 1-7.
Zhou W J, Wu W P, Tang C B, Xiao Z F, Chen G H, Wang Y. Effects of exogenous 2,4-epibrassinolide on germination and physiological characteristics of rice seedlings under chilling stress[J]., 2020, 29(9): 1-7. (in Chinese with English abstract)
[20] 李响珍. 水稻发芽期亚干旱对苗期、孕穗期与开花期耐冷性的影响[D]. 长沙: 湖南师范大学, 2019.
Li X Z. Effects of subdrought at germination stage on cold tolerance at seedling stage, booting stage and flowering stage in rice[D]. Changsha: Hunan Normal University, 2019. (in Chinese with English abstract)
[21] 胡涛. 低温对水稻根系生理特性及其基因表达的影响[D]. 沈阳: 沈阳农业大学, 2019.
Hu T. Effects of low temperature on physiological characteristics and gene expression of rice roots[D]. Shenyang: Shenyang Agricultural University, 2019. (in Chinese with English abstract)
[22] 吴立群, 蔡志欢, 张桂莲, 刘逸童, 赵瑞. 低温对不同耐冷性水稻品种秧苗生理特性及根尖解剖结构的影响[J]. 中国农业气象, 2018, 39(12): 805-813.
Wu L Q, Cai Z H, Zhang G L, Liu Y T, Zhao R. Effects of low temperature on physiological characteristics of rice seedlings with different cold tolerance and anatomical structure of root tip[J]., 2018, 39(12): 805-813. (in Chinese with English abstract)
[23] 向丹. 水稻苗期低温耐性差异及其调控研究[D]. 北京: 中国农业科学院, 2013.
Xiang D. The difference in low temperature tolerance of Rice Seedlings and its regulation[D]. Beijing: Chinese Academy of Agricultural Sciences, 2013. (in Chinese with English abstract)
[24] Farrell T C, Fox K M, Williams R L, Fukai S. Genotypic variation for cold tolerance during reproductive develop ment in rice: screening with cold air and cold water[J]., 2006, 98(2): 178-194.
[25] Gunawardena T A, Fukai B. The interaction of nitrogen application and temperature during reproductive stage on spikelet sterility infield-grown rice[J]., 2005, 56: 625-636.
[26] 于艳梅, 李芳花, 姜丽霞, 孟岩. 不同淹水处理对寒地水稻生长规律及产量影响的研究[J]. 水资源与水工程学报, 2018, 29(3): 249-253.
Yu Y M, Li F H, Jiang L X, Meng Y. Effects of different flooding treatments on growth law and yield of rice in cold region[J]., 2018, 29(3): 249-253. (in Chinese with English abstract)
[27] 杨生龙, 贾志英. 不同水分处理对水稻和旱稻产量及产量构成因子的影响[J]. 安徽农业科学, 2010, 38(31): 17410-17412.
Yang S L, Jia Z Y. Effect of different water treatments on yield and yield components of lowland rice and aerobic rice[J]., 2010, 38(31): 17410-17412. (in Chinese with English abstract)
[28] Sanghera S G, Wani S H, Hussain W, Singh N B. Engineering cold stress tolerance in crop plants[J]., 2011, 12(1): 30-43.
[29] 许培磊. 低温对不同基因型黄瓜叶片抗氧化酶与蛋白质组的影响[D]. 泰安: 山东农业大学, 2009.
Xu P L. Effects of low temperature on antioxidant enzymes and leaf proteome fromof different genotypes[D]. Taian: Shandong Agricultural University, 2009. (in Chinese with English abstract)
[30] 喻方圆, 徐锡增. 植物逆境生理研究进展[J]. 世界林业研究, 2003, 16(5): 6-11.
Yu F Y, Xu X Z. A review on plant stress physiology[J]., 2003, 16(5): 6-11. (in Chinese with English abstract)
[31] Li Y S, Ou S L, Yang C Y. The seedlings of different japonica rice varieties exhibit differ physiological properties to modulate plant survival rates under submergence stress[J]., 2020, 9(8): 982.
[32] Minami A, Yano K, Gamuyao R, Nagai K, Kuroha T, Ayano M, Nakamori M, Koike M, Kondo Y, Niimi Y, Kuwata K, Suzuki T, Higashiyama T, Takebayashi Y, Kojima M, Sakakibara H, Toyoda A, Fujiyama A, Kurata N, Ashikari M, Reuscher S. Time-course transcriptomics analysis reveals key responses of submerged deepwater rice to flooding[J]., 2018, 176(4): 3081-3102.
[33] 何云, 李贤伟, 李西, 龚伟. 2 种野生岩生植物叶片可溶性蛋白含量对低温胁迫的响应[J]. 安徽农业科学, 2008, 36(18): 7552-7553.
He Y, Li X W, Li X, Gong W. Response of two kinds of wild rock plant leaf soluble protein content to low temperature stress[J]., 2008, 36(18): 7552-7553. (in Chinese with English abstract)
[34] 齐光, 佟伟霜, 杨雨华, 陈明辉, 吴秀菊. ABA 对低温胁迫下水稻幼苗抗氧化酶活性的影响[J]. 湖北农业科学, 2016, 55(23): 6079-6082.
Qi G, Tong W S, Yang L H, Chen M H, Wu Z J. Effects of aba on antioxidase activity in rice seedlings under low temperature stress[J]., 2016, 55(23): 6079-6082. (in Chinese)
[35] Bilska A, Sowinski P. Closure of plasmodesmata in maize () at low temperature: A new mechanism for inhibition of photosynthesis[J]., 2010, 106(5): 675-686.
[36] 何亚飞, 李霞, 谢寅峰. 核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)与Rubisco活化酶的分子机理研究进展[J]. 分子植物育种, 2017, 15(8): 399-405.
He Y F, Li X, Xie Y F. Advances in molecular mechanism of[J]., 2017, 15(8): 399-405. (in Chinese with English abstract)
[37] Sperotto R A, Cruz R P D, Cargnelutti D, Adamski J M, Fett J P. Avoiding damage and achieving cold tolerance in rice plants[J]., 2013, 2(2): 96-119.
[38] 姜籽竹, 朱恒光, 张倩, 宋北光, 孟丽君, 杨德光. 低温胁迫下植物光合作用的研究进展[J]. 作物杂志, 2015(3): 23-28.
Jiang Z Z, Zhu H G, Zhang Q, Song B G, Meng L J, Yang D G. Progress of influence of low temperature on plant photosynthesis[J]., 2015(3): 23-28. (in Chinese)
[39] Kato Y, Collard B C Y, Septiningsih E M, Ismail A M. Increasing flooding tolerance in rice: combining tolerance of submergence and of stagnant flooding [J]., 124(7): 1199-1210.
[40] 柏斌, 吴俊, 盛文涛, 庄文, 李莺歌, 邓启云. 育性敏感期低温胁迫对水稻光温敏不育系叶片内源激素的影响[J].杂交水稻, 2016, 31(1): 57-61.
Bai B, Wu J, Sheng W T, Zhuang W, Li Y G, Deng Q Y. Effects of cold stress at fertility sensitive stage on endogenous hormones of rice ptgms lines[J]., 2016, 31(1): 57-61. (in Chinese with English abstract)
[41] 曾研华, 张玉屏, 潘晓华, 朱德峰, 向镜, 陈惠哲, 张义凯. 花后低温对水稻籽粒灌浆与内源激素含量的影响[J]. 作物学报, 2016, 42(10): 1551-1559.
Zeng Y H, Zhang Y P, Pan X H, Zhu D F, Xiang J, Chen H Z, Zhang Y K. Effect of low temperature after flowering on grain filling and plant hormones contents in rice[J]., 2016, 42(10): 1551-1559. (in Chinese with English abstract)
[42] 刘春风. 淹水对15个树种苗木生长和形态特征的影响[D]. 南京: 南京林业大学, 2009.
Liu C F. Effects of artificial flooding on the growth and morphological characteristics of 15 tree species seedlings[D]. Nanjing: Nanjing Forestry University, 2009. (in Chinese with English abstract)
[43] 潘向艳, 季孔庶, 方彦. 淹水胁迫下杂交鹅掌楸无性系叶片内源激素含量的变化[J]. 南京林业大学学报, 2008, 32(1): 29-32.
Pan X Y, Ji K S, Fang Y. Changes in contents of endogenous hormones in different clones of×under flooding stress[J]., 2008, 32(1): 29-32. (in Chinese with English abstract).
[44] Zhou Y J, Li X X, Cao J, Li Y, Huang J L, Peng S B. High nitrogen input reduces yield loss from low temperature during the seedling stage in early-season rice[J]., 2018, 228: 68-75.
[45] Si L L, Xie Y N, Ma Q X, Wu L H. The short-term effects of rice straw biochar, nitrogen and phosphorus fertilizer on rice yield and soil properties in a cold waterlogged paddy field[J]., 2018, 10(2): 537.
Study on Physiological Characteristics Behind Mitigative Effects of Flooding on Low Temperature-caused Chilling Damage to Direct Seeded EarlyRice at the Seedling Stage
WANG Wenxia, CHEN Liming, WANG Haixia, LIU Youqing, WU Ziming, ZENG Yongjun, TAN Xueming,PAN Xiaohua, SHI Qinghua, ZENG Yanhua*
(,,,,;,:)
【】It is important to explore the effects of flooding on the growth of direct seeded earlyrice seedlings under low-temperature stress, and lay a physiological basis for direct seeded rice production and stress-resistant cultivation in South China.【】The cold-tolerant cultivar Xiangzaoxian 6 and cold-sensitive cultivar Zhongjiazao 17 were used as materials in this experiment, and three treatments each lasting three days including low temperature treatment (8ºC), low temperature & flooding treatment (8ºC plus flooding) and ambient temperature as control treatment (25ºC) were designed to analyze the physiological characteristics including agronomic characters, antioxidant enzyme activities, osmotic regulation substances contents, photosynthesis enzyme activities and endogenous hormone contents of rice seedlings.【】Compared with low temperature (LT), the seedling height, fresh weight, dry weight and T3 (the 3rd leaf from top) leaf length of different cultivars significantly increased after three days at low temperature under flooding (LTF), and the root number, root length, T1 (the first leaf from top) leaf length and T2 (the 2nd leaf from top) leaf length also increased. Flooding could reduce the damage induced by low temperature to the agronomic characters of rice seedlings with the cold-tolerant cultivar Xiangzaoxian 6 less affected as compared with cold-sensitive cultivar Zhongjiazao 17. In addition, compared with LT, LTF significantly decreased the antioxidant enzymes activities (SOD, POD, CAT), soluble protein contents, osmotic regulatory substances contents (MDA, proline) and endogenous growth inhibitory hormone contents, ABA content, whereas increased the content of endogenous growth promoting hormone GA3under low temperature; Flooding could alleviate the accumulation of reactive oxygen species (ROS) and the aggravation of membrane lipid peroxidation in plants, and improve the regulation of endogenous hormones in plants. Meanwhile, LT and LTF significantly decreased the contents of chlorophyll and ATP in leaves, resulting in the decrease of photosynthetic enzyme activities (Rubisco, PEPCK); but LTF had lower damage than that of LT, and LTF could play a mitigating role. After the recovery treatment, the physiological activity indexes of rice subjected to LTF were close to the level of CK, whereas the self-response mechanism of cold-tolerant cultivar could reduce the damage caused by low temperature stress.【】Low temperature treatment affected the growth characteristics and photosynthetic enzyme activity of direct seeded earlyrice seedlings, and increased the antioxidant protective enzyme activity and osmotic regulation potential of leaves, but flooding treatment help to alleviate the physiological damage of leaves caused by low-temperature stress.
earlyrice; direct seeded rice; low temperature; flooding; physiological characteristics
10.16819/j.1001-7216.2021.0914
2020-09-23;
2020-12-19。
国家自然科学基金资助项目(31760366);国家重点研发计划资助项目(2017YFND0301605);江西省杰出青年自然科学基金资助项目(20202ACBL215004);江西省自然科学基金资助项目(2016BAB214171);江西省水稻产业技术体系专项( JXARS-02-03)。
