北京综合性社区鸟类群落构成及其季节变化∗
——以中国林业科学研究院社区为例
2021-03-16孙宝强赵伊琳
徐 诗 王 成 韩 丹 孙宝强,3 赵伊琳
1 中国林业科学研究院林业研究所 国家林业和草原局林木培育重点实验室 北京 100091
2 国家林业和草原局城市森林研究中心 北京 100091
3 天津财经大学艺术学院 天津 300222
随着全球城市化进程的加快, 城市中能够为野生动物提供栖息的自然绿地日益减少, 城市生物多样性受到严重破坏, 这引起了部分生态学家和城市自然保护者的关注[1-2]。 城市鸟类多样性是城市生物多样性的重要组成部分[3],一个地区的城市鸟类群落已经成为评价该地区生态环境质量的重要指标[4]。 国外早在20 世纪30、 40 年代就开始了对城市鸟类的关注[5], 近年来在鸟类群落生态学的研究发展迅速, 并伴产生大量的新理论、 新方法。 我国城市鸟类群落生态学研究的起点较早, 但发展缓慢[6]。 近年来国内对于鸟类栖息地的研究日益增多, 研究内容主要涉及鸟类区系, 鸟类生理、 栖息地选择, 繁殖和人为干扰等领域[7]。
北京地区鸟类资源丰富, 2016 年北京鸟类协会提供的《北京鸟类名录2016》 显示, 北京地区共有鸟类21 目74 科469 种。 自20 世纪60年代开始, 我国很多学者陆续对北京地区的鸟类群落展开了基础调查研究。 郑光美[8-9]对北京及其附近地区不同季节的鸟类进行了长达9 年的调查, 记录了北京地区140 种鸟类的群落构成、 空间分布和动态变化; 赵欣如等[10]于1984—1986 年对北京的6 个典型公园进行了鸟类调查, 记录到134 种鸟类; 陈志强等[11]于2002—2008 年在圆明园遗址公园调查记录到鸟类217 种; 滑荣等[12]于2012—2019 年在颐和园调查记录到鸟类168 种。 目前, 研究者对城市鸟类的研究范围多数集中在城市公园绿地和自然保护区内, 鲜有在居民社区中的研究。 然而,在城市区域范围内, 各类社区已经成为城市的构成主体, 同时也是城市鸟类活动的重要区域,社区环境直接关系到整个城市生态系统的健康。因此, 提高城市生物多样性需要加大对城市社区生物多样性的重视。 本文以中国林业科学研究院社区为例, 对其鸟类群落结构及其季节变化展开调查研究, 旨在为北京社区的建设及鸟类保护提供理论依据。
1 研究区概况
中国林业科学研究院社区(以下简称林科院社区(CAFC) ) 位于北京西北郊, 北依大昭山, 南望玉泉山, 东南为颐和园, 其海拔58 m,占地总面积26.3 hm2, 其中绿地面积为10.3 hm2。该社区涵盖了办公、 教育、 科研、 居住等功能,人口集中且固定, 社区林木覆盖率高达48%,绿地类型丰富多样, 植物种类繁多, 是一个复合型的综合社区, 在北京非常具有典型性。
2 研究方法
2.1 样地设置
根据林科院社区的空间形态、 土地利用及绿地分布状况, 设置了可能完整涵盖整个区域和绿地类型的调查路线, 全长约3.4km。 根据研究区内不同的植被类型和植物生境特点, 将其划分为28 个调查板块(图1), 每个板块设置1 条局部样线, 确保样线覆盖调查区域内的全部生境类型。
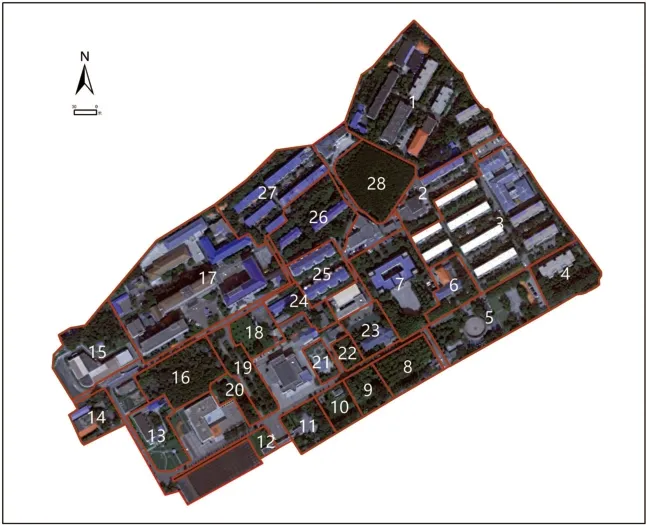
图1 林科院社区28 个板块划分图
2.2 鸟类调查
鸟类调查采用样线法, 于2018 年4 月—2019年3 月, 每两周选择晴朗无风天气调查1 次, 每季度6 次。 调查中使用双筒望远镜、 数码相机和录音笔, 记录拍到的、 听到的、 看到的鸟类基本情况, 包括观测的时间、 地点和鸟类的数量、 种类、 栖息树种、 活动状态和生境概况等。
2.3 数据处理
本研究采用以下6 个指数描述林科院社区鸟类群落的基本特征。
1) 鸟类丰富度, 即采用Patrick 指数反映样地中鸟类种数的多少。 计算公式为R=S, 其中R表示鸟类丰富度,S表示单位样方中的鸟类种数。
2) 鸟类相对多度, 即反映某个鸟类的个体数量在全部鸟类总数量的比例。 计算公式为Pi=,其中Pi表示鸟类i 的相对多度,Ni表示第i种鸟的个体数,N0表示样地内鸟类的总个体数。
3) 鸟类多样性指数,即采用Shannon-Wiener指数反映样地群落中鸟类的多样性程度。 计算公式为,其中H′表示鸟类多样性,Pi表示第i 种鸟的个体数占总鸟类数量的比例。
4) 鸟类均匀度指数, 即鸟类个体数在群落中分配的均匀程度。 采用Pielou 指数, 其计算公式为。 其中J表示物种均匀度,S表示物种数量,Pi表示第i 种的个体数占总物种数量的比例。
5) 鸟类数量等级。 划分方法参考了陈媛的频率指数估计法[13], 将每种鸟遇见的百分率(R) 与平均每天遇见该种鸟数量(B) 的乘积(RB) 作为指数进行数量等级划分:

当RB≥40 为优势, 5≤RB<40 为常见种,RB<5 为稀有种[13]。
6) 鸟类平均多度, 即反映样地的鸟类数量在持续观测时间内的平均水平。 鸟类平均多度M=每样地n次调查总数量的平均数。
3 结果与分析
3.1 林科院社区全年鸟类群落构成
3.1.1 群落组成
调查共记录到鸟类31 种, 分属于5 目17科, 其中雀形目鸟类种类最多, 共13 科25 种,占全部鸟类种数的80%, 而非雀形目鸟类4 科6种, 占总数的20% (表1), 这表明林科院社区鸟类的主要类群是以雀形目为主。
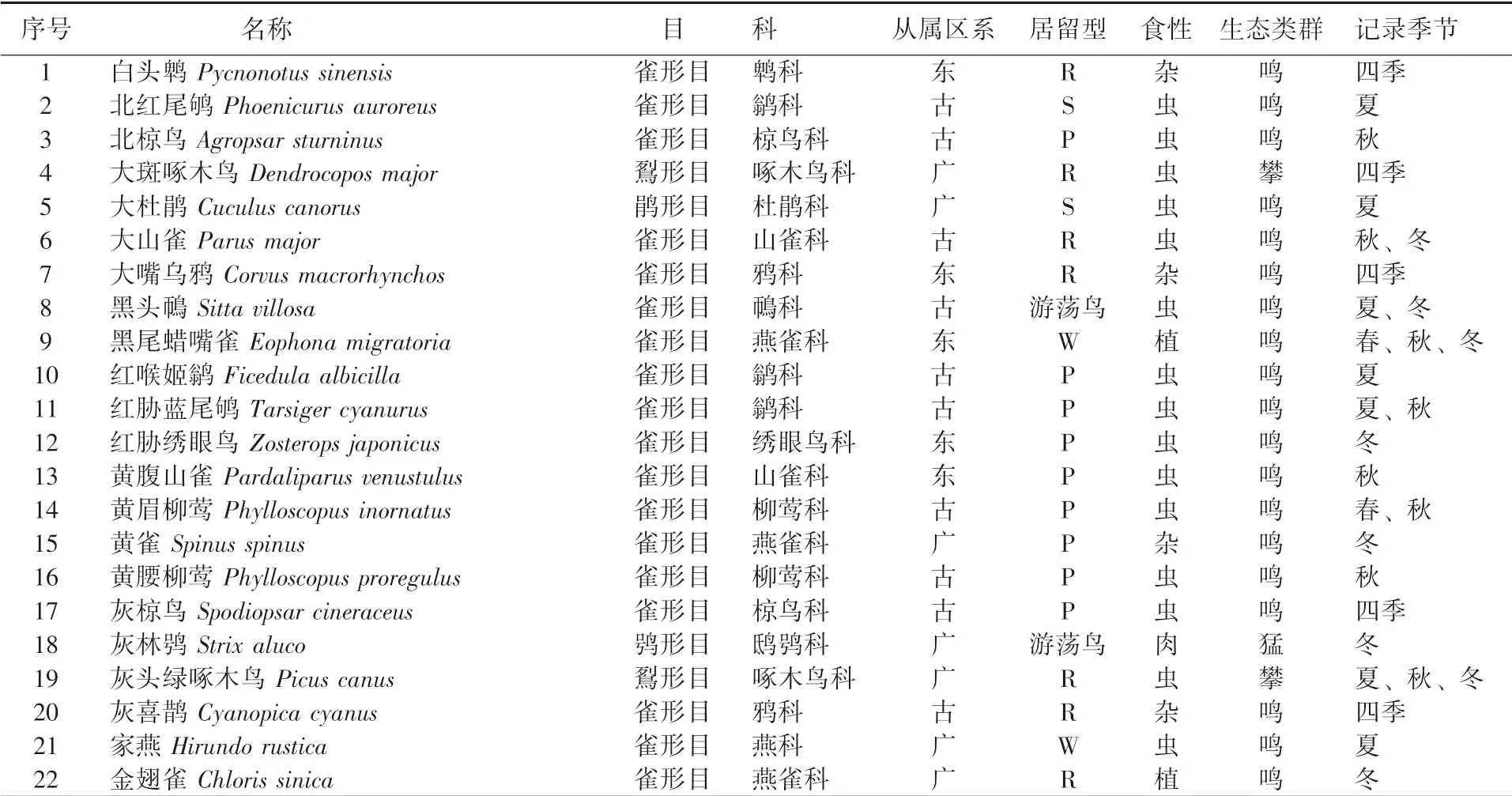
表1 林科院社区鸟类名录
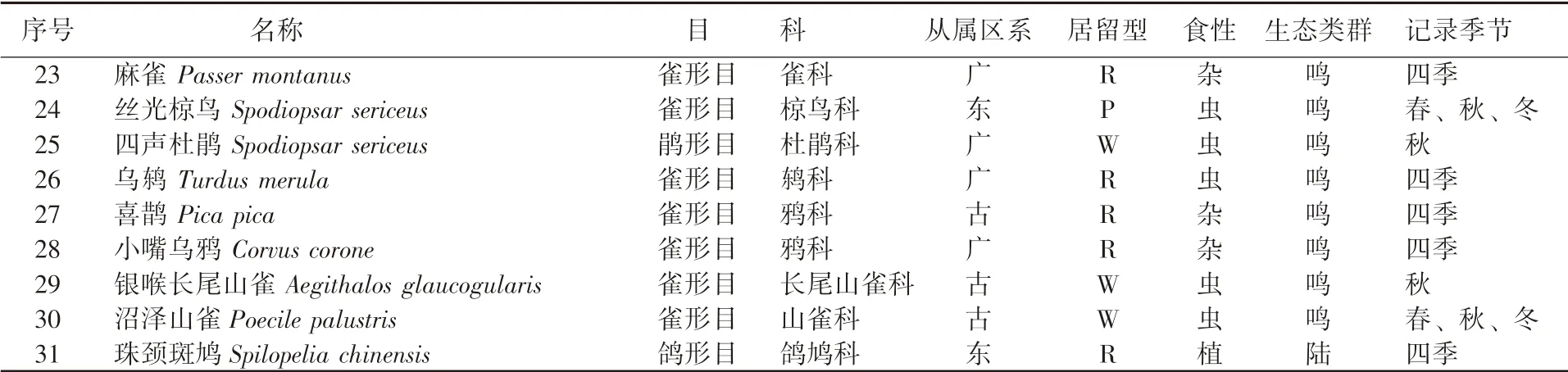
表1 (续)
3.1.2 类型特征
从林科院社区鸟类区系组成看, 古北种共计17 种, 约占全部记录种类的54%; 而广布种和东洋种各为7 种, 均占记录的23%。 从鸟类的居留型构成看, 留鸟12 种、 旅鸟10 种、 夏候鸟4 种、 冬候鸟3 种, 留鸟和旅鸟占比较大。 从鸟类的生态类群看, 鸣禽27 种、 攀禽2 种、 陆禽1 种、 猛禽1 种, 鸣禽种数占比87%。 从鸟类的取食类型看, 虫食性鸟类20 种、 杂食性鸟类7 种、 植食性鸟类3 种、 肉食性鸟类1 种, 虫食性鸟类占比达64%。
3.1.3 优势种分析
频率指数估计的结果表明,RB≥40(优势种)有2 种,分别是麻雀和灰喜鹊;5≤RB<40(常见种)有2 种,分别是喜鹊和白头鹎;RB <5(稀有种)有27 种,包括乌鸫、珠颈斑鸠等。 从表2 可以看出,在频率指数数值排序上,麻雀RB 值远大于其他鸟类,是林科院社区鸟类的绝对优势种。
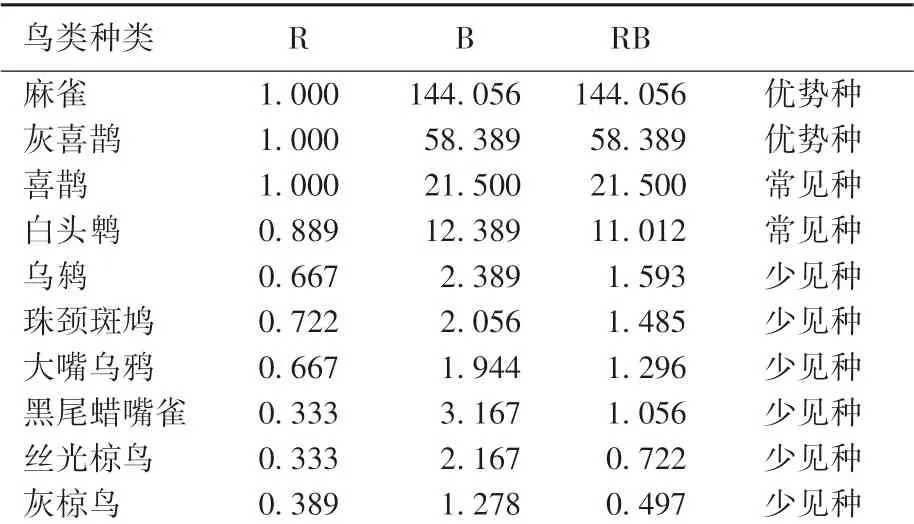
表2 林科院社区鸟类频率指数
3.2 林科院社区不同季节的鸟类群落构成
3.2.1 种类构成
林科院社区全年鸟类群落在种类构成上呈现出一定的季节性变化, 表现为秋季最多, 春季次之, 夏季最少。 其中春季调查到鸟类3 目14 科19 种(占比25.68%), 以留鸟为主, 还包括部分的旅鸟、 冬候鸟和夏候鸟; 夏季鸟类有4目14 科16 种(21.62%), 包括留鸟、 夏候鸟和部分 旅 鸟; 秋 季 鸟 类 4 目 14 科 21 种(28.38%), 以留鸟和旅鸟为主, 还观察到部分候鸟; 冬季3 目11 科18 种, 包括留鸟、 冬候鸟和部分旅鸟(24.32%)。
3.2.2 数量差异
林科院社区不同季节的鸟类数量变化差异较大, 表现为冬季最多, 秋季次之, 夏季最少(图2)。 一年之中, 冬季鸟类记录数量最多, 这与麻雀、 灰喜鹊等北京比较典型的留鸟于冬季在社区内的集群性活动有关。
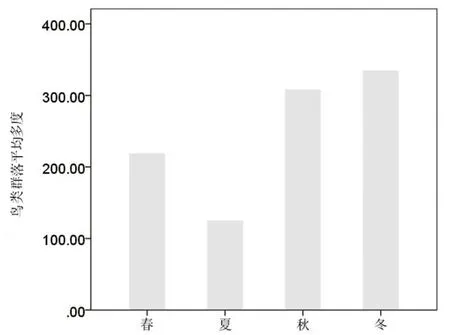
图2 林科院社区鸟类群落平均多度季节变化
3.3 不同月份鸟类多样性变化
林科院社区全年鸟类群落的丰富度指数总体波动变化略大, 10 月份达到了峰值, 3 月份达到第2 个峰值, 其他月份呈波动状态(图3)。春秋两季是一年中鸟类的迁徙高峰期, 北京地区作为鸟类迁徙路线(东亚-澳洲迁徙路线) 中的必经驿站之一, 成为很多旅鸟迁徙中途落脚、休息的重要通道, 因此这时候记录到的鸟类物种数量明显增加, 10 月达到全年最大值。 这是因为社区内存在一定比例的食源树种, 10 月份大部分植物果实成熟, 鸟类拥有了更大的捕食空间, 因此吸引了更多种类的迁徙鸟来此驻足。
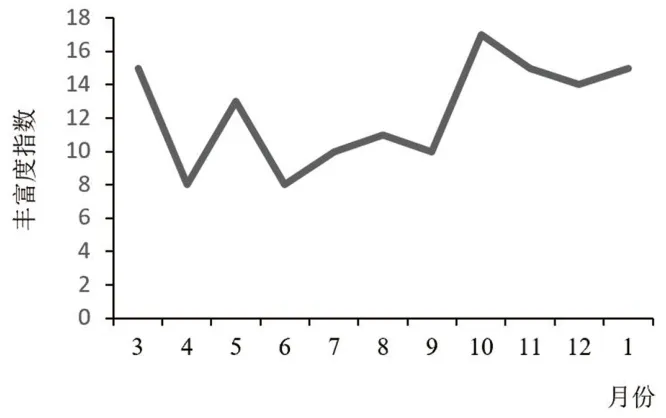
图3 林科院社区鸟类群落丰富度指数的月变化
鸟类的平均多度变化相对最为显著, 从3—8 月, 呈下降趋势, 并于8 月达到最低点, 8—11 月呈急剧上升趋势, 11 月达到峰值, 而后从11 月到次年3 月又开始缓慢下降(图4)。 春末夏初, 社区中的鸟类进入繁殖期, 观察到的鸟类逐渐变少, 于8 月数量降到最低。 11 月鸟类记录数量最多, 这可能与麻雀、 灰喜鹊等鸟类在冬季的集群性活动有关, 这对社区内鸟类总数量的影响较大。
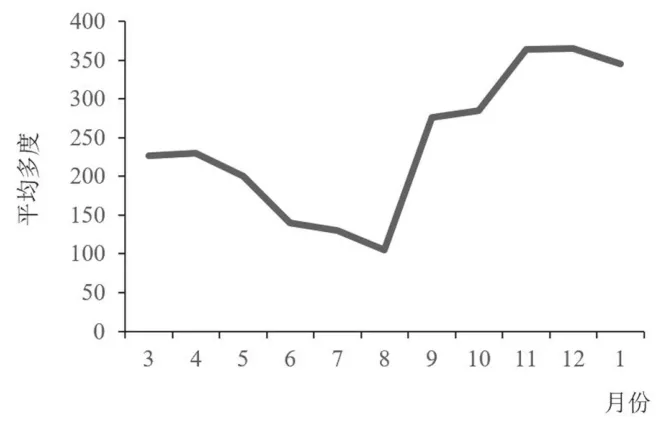
图4 林科院社区鸟类群落平均多度月变化
鸟类的多样性指数和均匀度指数变化趋势相似, 一年中均出现了两个峰值和两个谷值,两个谷值均在7 月和9 月, 多样性指数最高值出现在10 月, 均匀度最高值出现在8 月, 其他月份变化平缓(图5、 图6)。 由于林科院社区中的优势种和常见种的数量要远大于其他少见种的数量, 并且其所包含的4 种鸟类均为留鸟,这就保证了社区内全年鸟类群落结构处于一个相当稳定的水平。 因此, 每当到了春秋鸟类迁徙季, 随着社区内候鸟和旅鸟的增多,H′值和J值升高, 到10 月和3 月达到两个峰值, 当迁徙期结束后,H′值和J值回落到原来的水平。 在5—9 月, 又出现了一次波动, 这可能是由于春末夏初, 鸟类进入繁殖、 育雏阶段, 一些鸟类主要围绕巢穴活动, 总体观测到的鸟类种类和数量都减少,H′值呈缓慢下降趋势; 而后随着大量优势种幼鸟数量的增加,H′值和J值持续下降(6—7 月), 然而, 由于优势种(主要是麻雀) 幼鸟离巢后的大量死亡[14],H′值和J值升高, 随着存活下来的幼鸟不断长大, 群落逐渐趋于稳定,H′值和J值又回落到之前的水平。
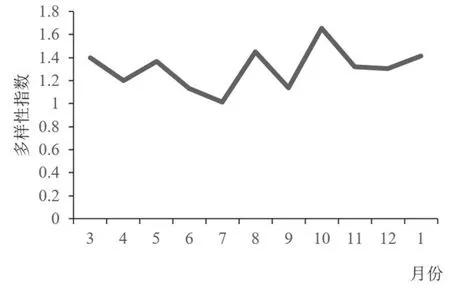
图5 林科院社区鸟类群落多样性月变化
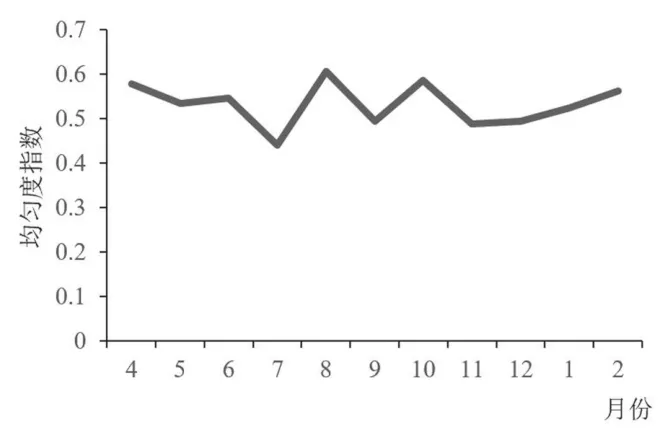
图6 林科院社区鸟类群落均匀度月变化
4 结论与讨论
4.1 结论
本次在北京中国林科院社区的调查研究共记录到鸟类5 目17 科31 种, 占北京地区记录鸟类物种数量的6.61%, 其中, 留鸟和旅鸟占比较大。鸟类区系以古北界为主, 生态类群以鸣禽为主,取食方式以虫食性为主。 全年鸟类优势种为麻雀和灰喜鹊, 常见种为喜鹊和白头鹎, 其余27 种均为少见种。 全年鸟类群落在种类构成上呈现出一定的季节性变化, 表现为秋季最多、 春季次之、夏季最少, 并于10 月达到峰值。 全年鸟类数量变化显著, 表现为冬季最多、 秋季次之、 夏季最少,并于8 月达到最小值, 11 月达到最大值。
4.2 讨论
中国林科院社区位于北京西北部, 是背靠山地森林和拥有较高林木覆盖的一个综合性社区[15]。 林科院社区鸟类调查结果与贾丽丽[16]在2013—2015 年对北京地坛公园记录到的43 种和张晥清[17]在2013—2015 年对北京奥林匹克森林公园调查记录到的66 种相比结果偏低, 这可能是因为公园的绿化条件一般优于居民社区, 并且公园内有较大的水域面积, 可以为鸟类提供更为理想的栖息地环境。 从优势种来看, 林科院社区的全年优势种为麻雀和灰喜鹊, 与北京圆明园遗址公园记录到的优势种一致[11], 北京颐和园[12]和北京奥森公园[17]的优势种中也包含这两个物种, 表现出在优势种上的总体相似性。
林科院社区鸟类群落结构呈现出很强的季节性, 引起这种变化的原因主要来自候鸟的季节性迁徙和留鸟的季节性繁殖。 物种数在春秋鸟类迁徙季出现两个高峰, 这与北京玉渊潭公园[18]、 北京奥林匹克森林公园[17]的调查结果一致。 林科院社区夏季物种数低于冬季, 而奥林匹克森林公园[17]和圆明园遗址公园[11]夏季物种数均高于冬季, 这与园内丰富的水生态环境有较大的关系, 夏季公园内会记录到更多种类的水鸟, 而社区内水资源缺乏, 导致夏季记录到的物种数量整体偏低。 鸟类数量受优势种集群性活动的影响比较大, 北京其他地区记录到的鸟类数量大多为春秋迁徙季最多, 而在林科院社区记录到的鸟类数量表现为冬季最多, 这可能是因为冬季社区内有较多的挂果树种, 这些挂果树种为当地留鸟提供了越冬生存所必需的食物, 从而保障了更多留鸟的生存。
研究鸟类群落结构变化规律, 观测鸟类季节动态, 一方面可以客观掌握社区生态环境基础资源; 另一方面可以为建设和优化现有的社区环境提供建议。 在社区绿地建设中, 为提高鸟类生物多样性, 需要加大对鸟类生境的关注, 适当增加水景观(尤其是在夏季), 营造多种类型的鸟类微生境, 增加一定数量和不同类型的冬季挂果树种。 考虑到食虫性鸟类占比高达64%, 在绿地中应尽量减少农药喷洒, 采用生物防治, 这也为社区植物的后期管护提供了新思路。
