游离氨对活性污泥系统中硝化性能和硝化群落结构的影响
2021-03-11姜婷婷王光杰杨浩吴常峰孙洪伟陈桐生
姜婷婷 王光杰 杨浩 吴常峰 孙洪伟 陈桐生


摘 要:建立了4个平行的SBR处理合成废水,游离氨(FA)浓度分别为0.5、5、10、15 mg/L,命名为S0.5、S5、S10和S15,4个系统的脱氮性能在整个实验过程中均很好(平均值为98.7%),利用FA对亚硝酸氧化细菌(NOB)的抑制作用,结合过程控制,成功在S10和S15系统中实现短程硝化。在建立短程硝化途径的过程中,S10的NAR在第79天迅速达到90.3%,S15的NAR在139天迅速达到90.5%。在S10的80~250 d和S15的140~250 d中,平均NAR分别稳定在98.8%和98.2%左右。用16S rRNA基因的高通量测序技术分析样本中硝化细菌的相对丰度和结构,结果表明,AOB和NOB丰度的变化与试验结果一致。FA不仅可以显著影响AOB和NOB的相对丰度,而且还可以抑制NOB活性。此外,还发现较低的AOB含量在FA浓度为15 mg/L时具有较高的氨底物微生物利用能力。
关鍵词:游离氨;短程硝化;高通量测序;氨氧化细菌;亚硝酸氧化细菌
Free ammonia (FA), a form of ammonia nitrogen (NH+4—N), is widely present in municipal/industrial wastewater. Commonly, Eq. (1) is used to calculate the concentration of FA[1].
There are positive relationships between the concentration of FA and NH+4—N, and the pH and T (temperature) based on Eq.(1).
FA has a significant effect on the nitrogen removal performance in the biological nutrient removal (BNR) pathway, because FA can affect the activity of nitrifying bacteria (including ammonia- oxidizing bacteria (AOB) and nitrite-oxidizing bacteria (NOB))[1-4]. The earliest inhibition threshold levels were reported by Anthonisen et al.[1], who demonstrated that AOB and NOB are inhibited by 10~150 mgFA/L and 0.1~1.0 mgFA/L, respectively. Considerable research has focused on the inhibition of AOB and NOB activity [5-8]. These studies mainly explored how to achieve effective partial nitrification during aerobic nitrification using the gap in the inhibitory concentration of FA between AOB and NOB. Nitrification, a two-step microbial process, is the oxidation of ammonia to nitrite by AOB and of nitrite to nitrate by NOB[9-12]. Partial nitrification is usually achieved by using the difference in activity between AOB and NOB, so ammonia is only oxidized to nitrite[13-15]. Over several decades, much research has been performed on stabilizing partial nitrification in BNR systems to remove nitrogen from high ammonia nitrogen wastewater by adjusting the concentration of FA to a specific level that inhibits NOB rather than AOB[16-18].
It is widely acknowledged that the nitrogen removal performance in BNR systems is affected by the activity of nitrifying bacteria[19-20]. From the perspective of the biological inhibition mechanism, the relative abundance and structure of nitrifying bacteria in the sewage treatment system is significantly influenced by the concentration of FA. Recently, Illumina high-throughput sequencing (HTS) technology was used to gain an in-depth understanding of characteristics of the microbial community in the water treatment system. During past years, much literature focused on the relative abundance and the structure of the nitrifying bacteria caused by FA in the BNR process[21-23]. These studies revealed that the relative abundance and structure of nitrifying bacteria are significantly affected at 2.9~50.1 mgFA/L.
Although exploration of the effects of FA on nitrifying bacteria during aerobic nitrification is increasingly successful, there is still a technical problem in that there is no consistent threshold level at which FA inhibits AOB and NOB. Thus, it is essential and urgent to investigate a consistent level at which FA inhibits AOB and NOB in biological systems. Most reported studies focused on the nitrifying bacteria in wastewater treatment bioreactors with a random FA concentration range (the concentration of FA depends on the quality of the wastewater). Few studies investigated nitrifying bacteria in the bioreactor under the precisely controlled concentration of FA. In addition, although previous studies revealed that FA has adverse effects on the microbial activity and stability of sludge, detailed information on the mechanisms by which it accomplished this was limited to various sludge characteristics and the structure of the microbial community caused by the concentration of FA. To eliminate these drawbacks, it is indispensable to systematically assess the variety and structure of nitrifying bacteria of relative abundance under different concentrations of FA during aerobic nitrification.
During our experiment, we filled four parallel sequencing batch reactors (SBRs) with precisely controlled FA concentrations of 0.5, 5, 10 and 15 mg/L. Activated sludge (AS) was used to tame microorganisms exposed to a specific concentration of FA in the SBRs. First, the long-term nitrogen removal performance of the SBRs was investigated. A method was explored to successfully achieve rapid and stable partial nitrification in the SBR at a high concentration of FA. The variety and structure of AOB and NOB of relative abundance in the SBRs with different concentrations of FA and the related mechanisms were examined in detail using high-throughput sequencing of the 16S rRNA gene. The information obtained has implications for the biological mechanism of nitrogen removal and the mechanism by which FA inhibits nitrifying bacteria.
1 Material and methods
1.1 Batch experiments design and operationFour 4 L parallel SBRs with 0.5, 5, 10 and 15 mg/L, concentrations of FA were operated to enrich the microbial community. The concentration of ammonia, the temperature and the pH were adjusted to obtain different concentrations of FA. Every cycle of the four SBRs consisted of 5 min filling, aerobic reaction, anoxic reaction, settling, 5 min decanting, and an idling period. The aerobic reaction, anoxic reaction, settling and idling period were flexible because of the different initial concentrations of FA.
The nitrification and denitrification were performed by adjusting the ORP, pH and DO. The pH was adjusted by the addition of 0.1 mol/L HCl and 0.4 mol/L NaOH. The temperature control system was used to control the temperature in the SBR. The DO concentrations in the four SBRs were maintained at 1.0~2.5 mg/L by an air compressor during aeration and by continuous stirring at 150 r/min by a mechanical stirrer rotating during the anoxic reaction. Table 1 summarizes the reactor operation.
Activated sludge with a mixed liquor suspended solid (MLSS) of 3 000 mg/L was collected at a local domestic sewage treatment plant (WWTP) in Lanzhou, Gansu to start up the batch reactors.
The four SBRs were operated for 250 days under the above mentioned concentrations of FA. After sustained long-term steady treatment by the reactors, four acclimated activated sludge samples, S0.5, S5, S10 and S15, were obtained. On day 240, 12 samples were collected from the four SBRs (3 samples from each SBR). The collected samples were immediately mixed with absolute ethanol at a 1∶1 volume ratio, and placed in a refrigerator, where they were maintained at -20 ℃ for DNA extraction.
1.2 Synthetic media
Nutrients and trace elements in the synthetic media support microbial growth in the reactors. The synthetic media contains (adapted from Kuai et al.[24]). 115 mg/L NH4Cl, 385 mg/L CH3COONa, 26 mg/L of K2HPO4 and KH2PO4, and 2 mL of trace elements solution. 5.07 mg MgSO4·7H2O, 1.26 mg Na2MoO·2H2O, 2.49 mg FeSO4·7H2O, 0.41 mg CoCl2·6H2O, 0.44 mg ZnSO4·7H2O, 0.31 mg MnSO4·4H2O, 0.43 mg CaSO4·2H2O, 0.25 mg CuSO4, 1.88 mg EDTA and 0.25 mg NaCl were contained per liter in the trace elements solution.
1.3 Analytical measurements
The concentrations of NH+4—N, NO-2—N, NO-3—N and COD were measured every 30 minutes to determine the performance of the reactor. These parameters were monitored simultaneously in both influent and effluent throughout the experiment. Standard methods (APHA[25])were used to measure NH+4—N, NO-3—N, NO-2—N, COD and MLVSS. The pH/oxi 340 analyzer (WTW, Germany) was used to measure the pH, DO and temperature.
1.4 Calculations
The concentration of FA was calculated by Eq.(1).
The ammonia oxidation rate (AOR) was calculated by Eq.(2).
The nitrite oxidation rate (NOR) was calculated by Eq. (3).
The nitrite accumulation rate (NAR) was calculated by Eq. (4).
Where NH+4—Ninf and NH+4—Neff are the ammonia concentrations (mg/L) in the influent and the effluent, respectively; NO-2—Ninf and NO-2—Neff are the nitrite concentrations (mg/L) in the influent and the effluent, respectively; NO-3—Neff is the nitrate concentration in the effluent (mg/L); T is the temperature (℃); t is the running time (min); and MLVSS is the mixed liquid volatile suspended solids (mg/L).
The biological reaction rate is strongly affected by the concentration of FA. Past studies proved that AOR and NOR show a sharp downward trend when the concentration of FA is higher than 25 mg/L[26-28]. In our study, we also found that variation in the concentration of FA significantly affects the AOR and NOR during nitrification throughout the experiments. A considerable increase in AOR occurred from 0.038 mgN/gVSS·min at 0.5 mg/L (S0.5) to a maximum of 0.093 mgN/gVSS·min at 10 mg/L (S10), and then slightly declined to 0.073 mgN/gVSS·min at 15 mgFA/L (S15). However, the NOR was maintained at 0.038±0.009 in S0.5 and S5 and 0.012±0.004 in S10 and S15 (Fig.2). The AOR was still maintained at a high level of 0.073-0.093 mgN/gVSS·min even though the concentration of FA was higher than 10 mg/L, indicating that stable and efficient NH+4—N removal was achieved.
A high level of NH+4—N removal efficiency (above 98%) was observed in the four SBRs, demonstrating that NH+4—N is removed, to a great extent, during nitrification and denitrification in the SBR. Therefore, the SBR is a useful reactor to remove NH+4—N from high nitrogen wastewater.
2.2 Achieving a nitrate pathway in S0.5 and S5 and a nitrate pathway in S10 and S15 for nitrogen removal
2.2.1 Achieving a nitrate pathway in S0.5 and S5 at a lower concentration of FA
In S0.5 and S5, the SBRs were operated for 250 days by adjusting the pH, DO and temperature to switch between aerobic and anaerobic. In the nitrified effluent, the nitrite concentrations were 0.38 mg/L and 0.51 mg/L, the nitrate concentrations were 28.2 mg/L and 66.5 mg/L, and NAR and the accumulation rate of nitrate were 0.57% and 0.50%, 98.8% and 99.1% in the S0.5 and S5, respectively (Fig. 3). Extremely low NAR and nitrate accumulation indicated that complete nitrification was obtained in S0.5 and S5. Simultaneously, the results indicated that the NH+4—N removal efficiency was not affected by the rise of FA concentration (0.5 to 5 mg/L) and full nitrification was achieved in both S0.5 and S5. The results showed that nitrifying bacteria could quickly adapt to specific concentrations of FA.
The concentrations of FA in S0.5 and S5 were 0.5 and 5 mg/L, that were both within and also much higher than the inhibition threshold of FA on NOB (begin at 0.1~1.0 mg/L) reported by Anthonisen et al.[1]. However, obvious nitrite accumulation was not found in S0.5 and S5 (Fig. 3(a)), showing that NOB activity could not be inhibited significantly when the FA is lower than 5 mg/L. Our research fully confirmed the conclusion of Vadivivelu et al.[2], who also found no significant decrease of NOB activity from 0 to 4 mgFA/L, indicating that a low concentration of FA has a small inhibitory effect on NOB.
In addition, nitrite accumulation did not occur in S0.5 and S5 even though we strictly controlled the process in each SBR cycle within 250 days. Our finding was consistent with Cao et al.[22]. There was no obvious nitrite accumulation by applying process control after 29 cycles at 2.9 and 5.6 mgFA/L, but they also achieved high total nitrogen removal efficiency averaging over 99%. However, Vlaeminck et al.[13] obtained stable nitrite accumulation and high nitrogen removal efficiency when the concentration of FA was higher than 3 mg/L, meaning that NOB suffered from strong suppression.
2.2.2 Initiating the nitrite pathway at a high concentration of FA in S10 and S15
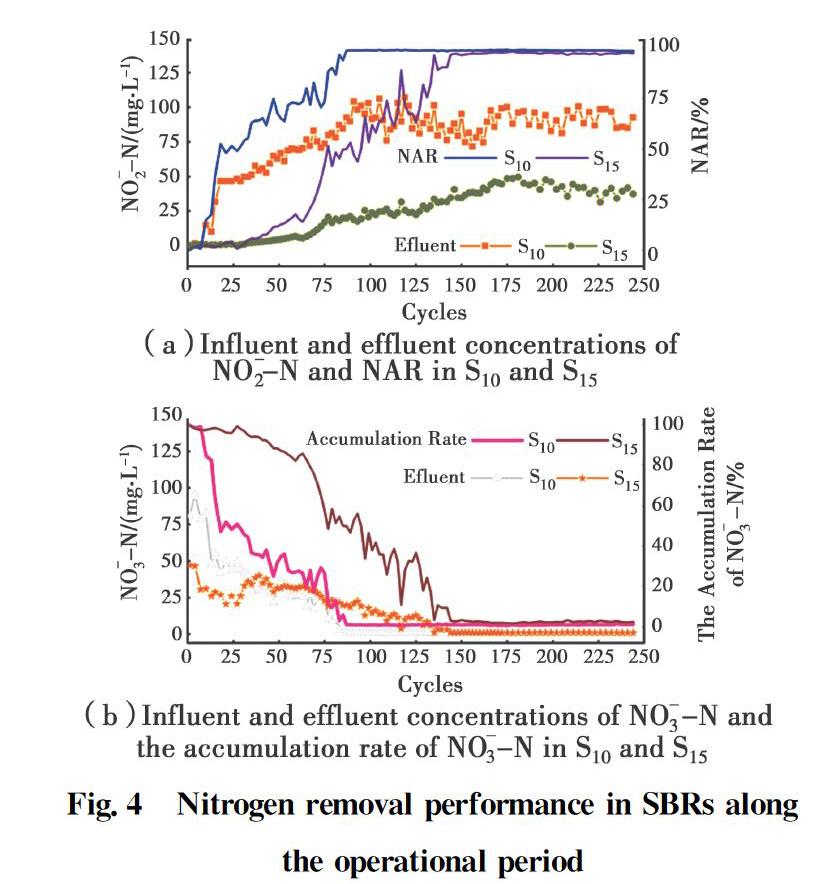
In the nitrified effluent, a remarkable decline took place in the NO-3-N concentration from 80.8 to 1.0 mg/L and 47.2 to 1.1 mg/L in S10 and S15, respectively (Fig. 4). However, the NO-2-N concentration increased considerably, from 0.07 to 81.2 mg/L and 0.18 to 48.3 mg/L in S10 and S15, respectively, indicating the accumulation of nitrite in S10 and S15. The NAR dramatically went up to 90.3% on day 79 in S10 and 90.5% on day 139 in S15, and the nitrate accumulation rate decreased to 1.1% on day 79 in S10 and 2.3% on day 139 in S15, respectively (Fig. 4). The increase of NAR with the decline of nitrate accumulation showed that NOB activity was greatly inhibited and partial nitrification was successfully achieved. AOB gradually became the dominant bacteria, because NOB was inhibited, forming a new nitrifying bacteria structure. S10 and S15 successfully achieved partial nitrification at a high concentration of FA.
The nitrite pathway was successfully established at 10 mgFA/L and 15 mgFA/L, and S10 achieved the nitrite pathway faster than S15 (Fig. 4). NOB activity was strongly inhibited in S15 and S10, while AOB activity was reduced more in S15 than S10, leading to the decline of the oxidation rate of the ammonia, and increasing the realization time of the nitrite pathway. Both the continuous ammonia oxidation process and the nitrite accumulation indicated that AOB activity wasn't greatly affected by 10 and 15 mgFA/L, which confirmed that AOB is more tolerant of FA than NOB.
Although several authors analyzed the inhibitory thresholds of FA on AOB and NOB, a consistent inhibitory threshold was not obtained. For example, the first inhibition threshold level of FA on AOB and NOB reported by Anthonisen et al.[1] was 10~15 mg/L and 0.1~1.0 mg/L, respectively. Kim et al.[4] suggested that only NOB was inhibited, while AOB could still oxidize ammonium to nitrite at 14~17 mgFA/L. Van Hulle et al.[27] reported that AOB activity was not inhibited in SHARON reactors at 70-300 mgFA/L.
Vadivelu et al.[2] found that the inhibitory effect of FA on NOB started from 1 mg/L and stopped growing when it was higher than 6 mg/L. In comparison, Vadivelu et al.[3] revealed that the inhibitory effect of FA on AOB started at 16 mg/L. Our research and previous studies showed that FA concentrations at S10 and S15 can strongly inhibit NOB activity, but have little effect on AOB activity.
2.2.3 Maintaining the nitrite pathway at a high concentration of FA in S10 and S15
During the period the nitrite pathway was maintained (S10 on day 80~250 and S15 on day 140~250), the main product in the nitrification was nitrite (81.2 and 48.3 mg/L), the nitrate concentration was still maintained at the bottom level (1.0 and 1.1 mg/L) and the NARs remained stable at 98.8% for 170 days and 98.2% for 110 days in S10 and S15, respectively (Fig. 4). Therefore, we can conclude that 10 mgFA/L and 15 mgFA/L can continue to inhibit NOB activity, and the nitrite conversion process was strongly inhibited.
In this study, the selective inhibition at high concentration of FA combined with process control is a basic method to achieve and stabilize the nitrite pathway in the treatment of synthetic wastewater in the SBR.
2.3 Confirmation of the dominant nitrifying bacterial population
Nitrifying bacteria plays a crucial role in nitrogen removal in the SBR. Therefore, we analyzed the relative abundance and structure of the nitrifying bacteria to better understand the microbial role in nitrification using high-throughput sequencing. Considerable studies have reported that five kinds of AOB (Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosolobus and Nitrosovibrio) and four kinds of NOB (Nitrococcus, Nitrospira, Nitrobacter and Nitrospira) were widely found in the sewage treatment system[29-32]. In this work, we found two types of nitrifying bacteria (Nitrosomonas (AOB) and Nitrospira (NOB)), which were regarded as the dominant nitrifying bacteria in WWTPs, as reported. A widespread and reliable consensus that NOB is even more sensitive to FA inhibition than AOB was reached and applied in numerous studies[6, 13, 23].
As depicted in Fig.5(a), the relative abundance of AOB and NOB suffered from a significant change by the variation in the concentration of FA, influencing the nitrogen removal. Surprisingly, we found that AOB activity was not inhibited, but enhanced at S10 with the increasing concentration of FA. AOB first increased sharply from 0.13% in S0.5 to 3.17% in S10 and then decreased to 0.60 in S15, but NOB linearly decreased from 6.14% in S0.5 to 0.96% in S15 and had a significant negative relationship with concentrations of FA (y=-0.3x+5.3, R2=0.72). The reason for the entirely different trend could be that FA can serve as a matrix for AOB to increase its relative abundance when the concentration of FA is lower than 10 mg/L, but FA reached the threshold for inhibiting AOB and made its abundance decrease when the concentration of FA was higher than 10 mg/L. However, the inhibition threshold of NOB was 0.5 mgFA/L. Thus, there was a gradual decline in the abundance of NOB with the increasing concentration of FA. The results were in agreement with AOR and NAR under the four FA treatments and confirmed that NOB was even more sensitive to FA inhibition than AOB.
Furthermore,it was clear that that NOB Nitrospira was the dominant nitrifier (AOB/NOB<1) at FA below 5 mg/L, and led to complete oxidation of ammonia to nitrate. But AOB Nitrosomonas was the predominant nitrifier (AOB/NOB=2.03) at 10 mg FA/L, and resulted in sufficient oxidation of ammonia to nitrite. A stable nitrite pathway was maintained at 15 mg FA/L with higher AOR (0.073 mgN/gVSS·min) than NOR (0.017 mgN/gVSS·min) although the relative abundance of NOB Nitrospira (0.95%) was higher than AOB Nitrosomonas (0.6%) (AOB/NOB=0.63) (Fig. 5(a) and (b)). In other words, a higher abundance of NOB Nitrospira had lower activity of utilizing the nitrite substrate, indicating that the NOB activity was strongly inhibited in S15. However, AOB Nitrosomonas exhibited the opposite trend. For example, the lower abundance had a higher microbial utilization capacity of the ammonia substrate. Thus, it was still able to make the AOR higher than the
NOR when the AOB abundance was greater than that of the NOB and remain stable during the ammoxidation process. Similar inhibitory threshold levels of FA on NOB were reported by Sun et al.[32], who obtained a nitrite pathway of over 90% at 16.3 mgFA/L in the UASB-SBR system treating landfill leachate and intense oppression occurred of FA on NOB activity.
Based on the results, we can draw the conclusion that FA can affect not only the relative abundance of AOB and NOB, but also the NOB activity. However, AOR decreases with AOB abundance. Thus, we cannot conclude that AOB activity is inhibited at 15 mgFA/L. Moreover, by adjusting the concentration of FA, promoting AOB but suppressing NOB, stable short-range nitrification was successfully achieved.
3 Conclusions
The SBR is an efficient and stable reactor to remove ammonia from synthetic wastewater. Stable partial nitrification was successfully achieved in the SBR at high concentrations of FA (10 and 15 mgFA/L). Although AOB and NOB coexist in the four systems, AOB is still the main nitrifying bacteria. This finding emphasizes the importance of cultivating the appropriate bacteria to achieve short-range nitrification. FA can affect not only the relative abundance of AOB and NOB but also the activity of NOB. Furthermore, we found that a lower abundance of AOB had a higher microbial utilization capacity of ammonia substrate at 15 mgFA/L.
Acknowledgements
The authors would like to acknowledge the financial support from the National Natural Science Foundation of China (Grant No. 51668031).
References:
[1] ANTHONISEN A, LOEHR R, PRAKASAM T, et al. Inhibition of nitrification by ammonia and nitrous acid [J]. Journal of Water Pollution Control Federation, 1976, 48(5): 835-852.
[2] VADIVELU V M, KELLER J, YUAN Z G. Effect of free ammonia on the respiration and growth processes of an enriched nitrobacter culture [J]. Water Research, 2007, 41(4): 826-834.
[3] VADIVELU V M, KELLER J, YUAN Z G. Effect of free ammonia and free nitrous acid concentration on the anabolic and catabolic processes of an enriched nitrosomonas culture [J]. Biotechnology andBioengineering, 2006, 95(5): 830-839.
[4] KIM D J, LEE D I, KELLER J. Effect of temperature and free ammonia on nitrification and nitrite accumulation in landfill leachate and analysis of its nitrifying bacterial community by FISH [J]. Bioresource Technology, 2006, 97(3): 459-468.
[5] JIANG Y S, POH L S, LIM C P, et al. Effect of free ammonia inhibition on process recovery of partial nitritation in a membrane bioreactor [J]. Bioresource Technology Reports, 2019, 6: 152-158.
[6] QIAN W T, PENG Y Z, LI X Y, et al. The inhibitory effects of free ammonia on ammonia oxidizing bacteria and nitrite oxidizing bacteria under anaerobic condition [J]. Bioresource Technology, 2017, 243: 1247-1250.
[7] AHN J H, YU R, CHANDRAN K. Distinctive microbial ecology and biokinetics of autotrophic ammonia and nitrite oxidation in a partial nitrification bioreactor [J]. Biotechnology and Bioengineering, 2008, 100(6): 1078-1087.
[8] PAMBRUN V, PAUL E, SPRANDIO M. Modeling the partial nitrification in sequencing batch reactor for biomass adapted to high ammonia concentrations [J]. Biotechnology and Bioengineering, 2006, 95(1): 120-131.
[9] YE Y Y, NGO H H, GUO W S, et al. A critical review on ammonium recovery from wastewater for sustainable wastewater management [J]. Bioresource Technology, 2018, 268: 749-758.
[10] WANG Q, NI B J, LEMAIRE R, et al. Modeling of nitrous oxide production from nitritation reactors treating real anaerobic digestion liquor [J]. Scientific Reports, 2016, 62: 53-36.
[11] WALTERS E, HILLE A, HE M, et al. Simultaneous nitrification/denitrification in a biofilm airlift suspension (BAS) reactor with biodegradable carrier material [J]. Water Research, 2009, 43(18): 4461-4468.
[12] JETTEN M S M, HORN S J, VAN LOOSDRECHT M C M. Towards a more sustainable municipal wastewater treatment system [J]. Water Science and Technology, 1997, 35(9): 171-180.
[13] VLAEMINCK S E, TERADA A, SMETS B F, et al. Nitrogen removal from digested black water by one-stage partial nitritation and anammox [J]. Environmental Science & Technology, 2009, 43(13): 5035-5041.
[14] CHUNG J, SHIM H, PARK S J, et al. Optimization of free ammonia concentration for nitrite accumulation in shortcut biological nitrogen removal process [J]. Bioprocess and Biosystems Engineering, 2006, 28(4): 275-282.
[15] CHUNG J, SHIM H, LEE Y W, et al. Comparison of influence of free ammonia and dissolved oxygen on nitrite accumulation between suspended and attached cells [J]. Environmental Technology, 2005, 26(1): 21-33.
[16] WEI D, ZHANG K, NGO H H, et al. Nitrogen removal via nitrite in a partial nitrification sequencing batch biofilm reactor treating high strength ammonia wastewater and its greenhouse gas emission [J]. Bioresource Technology, 2017, 230: 49-55.
[17] CHEN J W, ZHENG P, YU Y, et al. Enrichment of high activity nitrifers to enhance partial nitrification process [J]. Bioresource Technology, 2010, 101(19): 7293-7298.
[18] NISHIMURA F, HIDAKA T, NAKAGAWA A, et al. Removal of high concentration ammonia from wastewater by a combination of partial nitrification and anammox treatment [J]. Environmental Technology, 2012, 33(13): 1485-1489.
[19] ZHANG S J, PENG Y Z, WANG S Y, et al. Organic matter and concentrated nitrogen removal by shortcut nitrification and denitrification from mature municipal landfill leachate [J]. Journal of Environmental Sciences, 2007, 19(6): 647-651.
[20] HE Q L, GAO S X, ZHANG S L, et al. Chronic responses of aerobic granules to zinc oxide nanoparticles in a sequencing batch reactor performing simultaneous nitrification, denitrification and phosphorus removal [J]. Bioresource Technology, 2017, 238: 95-101.
[21] ZHANG C, QIN Y G, XU Q X, et al. Free ammonia-based pretreatment promotes short-chain fatty acid production from waste activated sludge [J]. ACS Sustainable Chemistry & Engineering, 2018, 6(7): 9120-9129.
[22] CAO J S, YU Y X, XIE K, et al. Characterizing the free ammonia exposure to the nutrients removal in activated sludge systems [J]. RSC Advances, 2017, 7(87): 55088-55097.
[23] SUI Q W, LIU C, ZHANG J Y, et al. Response of nitrite accumulation and microbial community to free ammonia and dissolved oxygen treatment of high ammonium wastewater [J]. Applied Microbiology and Biotechnology, 2016, 100(9): 4177-4187.
[24] KUAI L P, VERSTRAETE W. Ammonium removal by the oxygen-limited autotrophic nitrification-denitrification system [J]. Applied and Environmental Microbiology, 1998, 64:4500-4506.
[25] Anon Coliphage Detection (proposed). In Standard Methods For The Examination of Water and Wastewater[S]. APHA, Washington, 1995.
[26] WANG Q L, DUAN H R, WEI W, et al. Achieving stable main stream nitrogen removal via the nitrite pathway by sludge treatment using free ammonia [J]. Environmental Science & Technology, 2017, 51(17): 9800-9807.
[27] VAN HULLE S W, VOLCKE E I, TERUEL J L, et al. Influence of temperature and pH on the kinetics of the Sharon nitritation process [J]. Journal of Chemical Technology & Biotechnology, 2007, 82(5): 471-480.
[28] BALMELLE B, NGUYEN K M, CAPDEVILLE B, et al. Study of factors controlling nitrite build-up in biological processes for water nitrification [J]. Water Science and Technology, 1992, 26(5/6): 1017-1025.
[29] BOCK E. Growth of nitrobacter in the presence of organic matter. II. Chemoorganotrophic growth of nitrobacter agilis [J]. Archives of Microbiology, 1976, 108: 305-312.
[30] EHRICH S, BEHRENS D, LEBEDEVA E, et al. A new obligately chemolithoautotrophic, nitrite-oxidizing bacterium, Nitrospira Moscoviensis sp. nov. and its phylogenetic relationship [J]. Archives of Microbiology, 1995, 164(1): 16-23.
[31] DAIMS H, PURKHOLD U, BJERRUM L, et al. Nitrification in sequencing biofilm batch reactors: lessons from molecular approaches [J]. Water Science and Technology, 2001, 43(3): 9-18.
[32] SUN H W, PENG Y Z, SHI X N. Advanced treatment of landfill leachate using anaerobic-aerobic process: Organic removal by simultaneous denitritation and methanogenesis and nitrogen removal via nitrite [J]. Bioresource Technology, 2015, 177: 337-345.
(編辑 胡英奎)
