枯草芽孢杆菌信号肽依赖分泌表达系统研究进展
2021-03-11段春燕井明博
段春燕,井明博,宋 曦
(1.陇东学院 生命科学与技术学院,甘肃 庆阳 745000;2.甘肃省高校陇东生物资源保护与利用省级重点实验室,甘肃 庆阳 745000;3.陇东学院 农林科技学院,甘肃 庆阳 745000)
大肠杆菌是目前掌握最为成熟的基因克隆表达系统,经常被用作蛋白质分泌和表达的宿主菌,它遗传背景清晰、转化效率高、表达周期短、操作简单,因此使用范围很广。但是大肠杆菌也有一些不足之处:大肠杆菌表达的蛋白质形成包涵体,给下游的纯化处理带来不便,同时有些表达蛋白无法分泌至胞外,不利于测定酶的活性和直接使用[1]。枯草芽孢杆菌是基因工程中又一个被广泛使用的表达系统,常作为表达酶和外源蛋白的宿主菌使用。对于枯草芽孢杆菌的研究可以追溯到100年前,初期侧重于形态观察等方面的研究,1958年Spizizen在实验中采用了化学试剂,进而制备了枯草芽孢杆菌的感受态细胞,极大地促进了其在基因工程研究方面的进展[2]。1997年,枯草芽孢杆菌模式菌株168的基因组完成测序,全基因组共4.2Mb,含有4214810个碱基,200个调控蛋白参与1500个操纵子的调控,蛋白编码序列占87%的基因组。在4106个蛋白质编码基因中,已经可以明确或预测约58%的蛋白质编码基因功能[3]和约30%的产物功能[4],为研究枯草芽孢杆菌蛋白质的分泌机理提供了重要理论依据,推动了相关研究的进行[5]。现在枯草芽孢杆菌已经成为一种成熟的基因工程表达系统,饲料、食品等行业大多利用枯草芽孢杆菌高效表达系统来生产内源生化产物,其应用之广仅次于大肠杆菌[6]。
1 枯草芽孢杆菌基因表达系统的优势
1.1 容易获得
枯草芽孢杆菌广泛分布在土壤、腐败的有机物以及水环境中,容易在枯草浸汁中繁殖。
1.2 遗传操作简单
枯草芽孢杆菌的基因组测序已于1997年完成,DNA遗传背景清晰,便于操作[7]。质粒载体如pUB110、pC194、HCMC和大肠杆菌-枯草芽孢杆菌穿梭质粒如pHT43、pSP10等的研究应用,为枯草芽孢杆菌在基因工程中的发展应用提供了便利[8]。
1.3 蛋白分泌系统功能完善
枯草芽孢杆菌蛋白质分泌能力强,可以实现对多种蛋白质的有效分泌,分泌的时候,并不存在包涵体的形成,可以直接透过细胞膜向培养基释放,回收纯化目的蛋白质方法简单,
1.4 发酵工艺成熟
枯草芽孢杆菌嗜温好氧,培养基配方简单,代谢产物中含有蛋白酶、淀粉酶等酶类,在工业上应用成熟。
1.5 非致病性
生理生化背景清晰,细胞壁组成简单,只含有肽聚糖和磷壁酸,不含内毒素,可以在动物肠道中繁殖,为正常的肠道微生物,被美国食品药品管理局认为是生物安全级别的微生物,在饲用酶和益生菌的生产上具有极大的优势[9]。
2 枯草芽孢杆菌信号肽依赖分泌胞外蛋白途径
在外源基因的克隆表达过程中,重组蛋白一般以3种形式表达,包括细胞外的分泌表达、细胞内的可溶性表达和在细胞内形成不溶性的胞涵体。枯草芽孢杆菌为格兰氏阳性菌,只有一层细胞膜,蛋白可直接分泌至胞外而不形成胞涵体,其表达产物多数具有天然构象和生物活性,产物较易纯化,极大地方便了外源蛋白的下游加工和酶活测定,经枯草芽孢杆菌分泌的很多外源蛋白质已经应用于益生菌、酶制剂等工业生产领域[10],占有很大的市场份额。根据SubtiList数据库提供的SignalP算法软件,共294种蛋白质被预测为分泌蛋白质[11]。根据蛋白质分泌是否涉及到信号肽,可以将分泌途径分为两种:信号肽依赖以及不依赖的分泌途径[12]。其中枯草芽孢杆菌分泌的蛋白质大部分是依托于信号肽依赖的分泌途径分泌至胞外,因此这种途径也称为典型性途径。
2.1 信号肽的研究进展
20世纪70年代初,美国的细胞生物学教授Gunter Blobel提出信号肽假说。该假说理论认为信号肽位于蛋白的N端,可以结合并引导核糖体附着在内质网的极性通道内,肽链不断延伸并穿过内质网膜,进而延伸到腔内,信号肽酶水解信号肽,正确折叠肽链,进而实现向胞外的转运[13]。信号肽是一段连续的氨基酸序列,用于指导蛋白质跨膜运输,它一般由10~40个左右的氨基酸残基构成,包括三个区域:氨基端区(N端)、疏水端区(H端)和羧基端区(C区)。其中N端由赖氨酸和精氨酸等带正电荷的氨基酸组成,氨基酸残基与细胞膜带负电荷的磷酸基团相互作用。H端的长度以及疏水性在蛋白分泌中起着重要的作用,主要由异亮氨酸等20个或超过20个的中性氨基酸构成,疏水氨基酸残基接触到膜脂,形成α螺旋结构,在蛋白质跨膜转位起始阶段发挥重要作用。C区又称为加工区,存在丝氨酸等小分子氨基酸,在蛋白转位中或转位后,信号多肽在C区被信号多肽酶识别并切割,随后信号多肽被降解,蛋白质被释放至胞外[14]。
细胞外分泌表达是外源基因克隆表达中最为理想的表达形式,但是大部分外源蛋白都会在细胞内形成包涵体,只有少数外源蛋白可以在细胞外分泌表达。为了让许多原本不能分泌至胞外的外源蛋白进行细胞外分泌表达,目前解决方法多为直接利用表达宿主的信号肽序列或对载体的启动子进行表达后插入编码信号肽序列来引导外源蛋白胞外分泌[15]。有研究表明,信号肽连接后,在大肠杆菌、芽孢杆菌和乳酸杆菌等原核表达系统中分泌和表达了多种外源基因。同时信号肽在毕赤酵母等真核表达系统中也得到了广泛的应用。如魏蔷等将信号肽插入猪瘟病毒E2蛋白基因,实现了该基因在杆状病毒表达系统中的分泌表达,其中gp67信号肽可使E2基因分泌表达量、纯度、产率都得到较大提高[16]。祝发明等构建了枯草芽孢杆菌Tat信号肽,实现了青霉素G酰化酶(PGA)在枯草芽孢杆菌中的分泌表达,其中最佳信号肽可引导PGA的表达酶活高达0.83U/mL[17]。但是信号肽与目的蛋白质之间具有适配性,如Brockmeier等人以角质酶和脂解酶为例,通过高通量筛选的方法建立了信号肽文库,以期有效筛选枯草芽孢杆菌sec途径中的148个信号肽。结果显示,两者间并不存在对应关系,148个信号肽能引导角质酶基因在枯草芽孢杆菌中表达,但最有效促进角质酶分泌的信号肽引导脂解酶分泌的水平很低,仅为最高分泌量的5%[18],即使是同一种蛋白,用类型不同的信号肽进行引导,多数均可以表达,但分泌效率显著不同。因此选择合适的信号肽是影响蛋白质产量的重要因素。
2.2 枯草芽孢杆菌分泌胞外蛋白途径
枯草芽孢杆菌中至少存在4种信号肽依赖分泌途径,具体为Sec途径、Tat途径、ABC转运子途径和假菌丝蛋白输出途径[19]。ABC转运子途径和假菌丝蛋白输出途径只输出特定蛋白质,且分子量相对较小,目前研究较为清楚的是Sec途径和Tat途径,其中Sec途径广泛应用于枯草杆菌分泌蛋白质,信号肽分为两类:I型普通型信号肽、C区含有脂蛋白盒子的Ⅱ型信号肽。Tat途径通常情况下仅对组装成熟的蛋白质进行分泌,信号肽中一般包括两个相邻氨酸残基[20]。
Sec(general secretory pathway)转运途径。Sec途径是主要的蛋白分泌途径,很多文献中外源表达蛋白的分泌主要依托于Sec途径来实现[8],对于枯草芽孢杆菌来讲,大约存在着三百种分泌蛋白[21],很多分泌蛋白向胞外的转移主要依托于Sec途径来实现,Sec转运系统在转运过程中,发挥着重要作用的编码蛋白为SecA、SecD、SecE、SecF、SecY、ffh、ftsY。SecA本质上属于一种ATP酶,依托于ATP的水解驱动分泌蛋白前体穿膜,而SecYEG的构成主要包括了SecE、SecY和SecG,从蛋白转移酶的角度来说,SecD7和SecF作为其重要的辅助成分[22]。在Sec途径中,核糖体会先合成有着信号肽的分泌蛋白前体,此时蛋白为未折叠状态。Sec蛋白的ATP酶对ATP进行水解,信号肽在这种情况下将会引导分泌蛋白前体,向细胞膜外的转移主要依托于极性通道来实现,通道蛋白的构成主要包括了一组膜蛋白,其主要的职责是对分泌蛋白的细胞质进行定位和转移[23],分泌蛋白以肽链形式在转移阶段存在。在穿过细胞膜后,信号肽酶在C区切除信号肽,蛋白质被正确折叠,并穿过细胞壁,最终作为成熟蛋白分泌到培养基中[24],详见图1[25]。
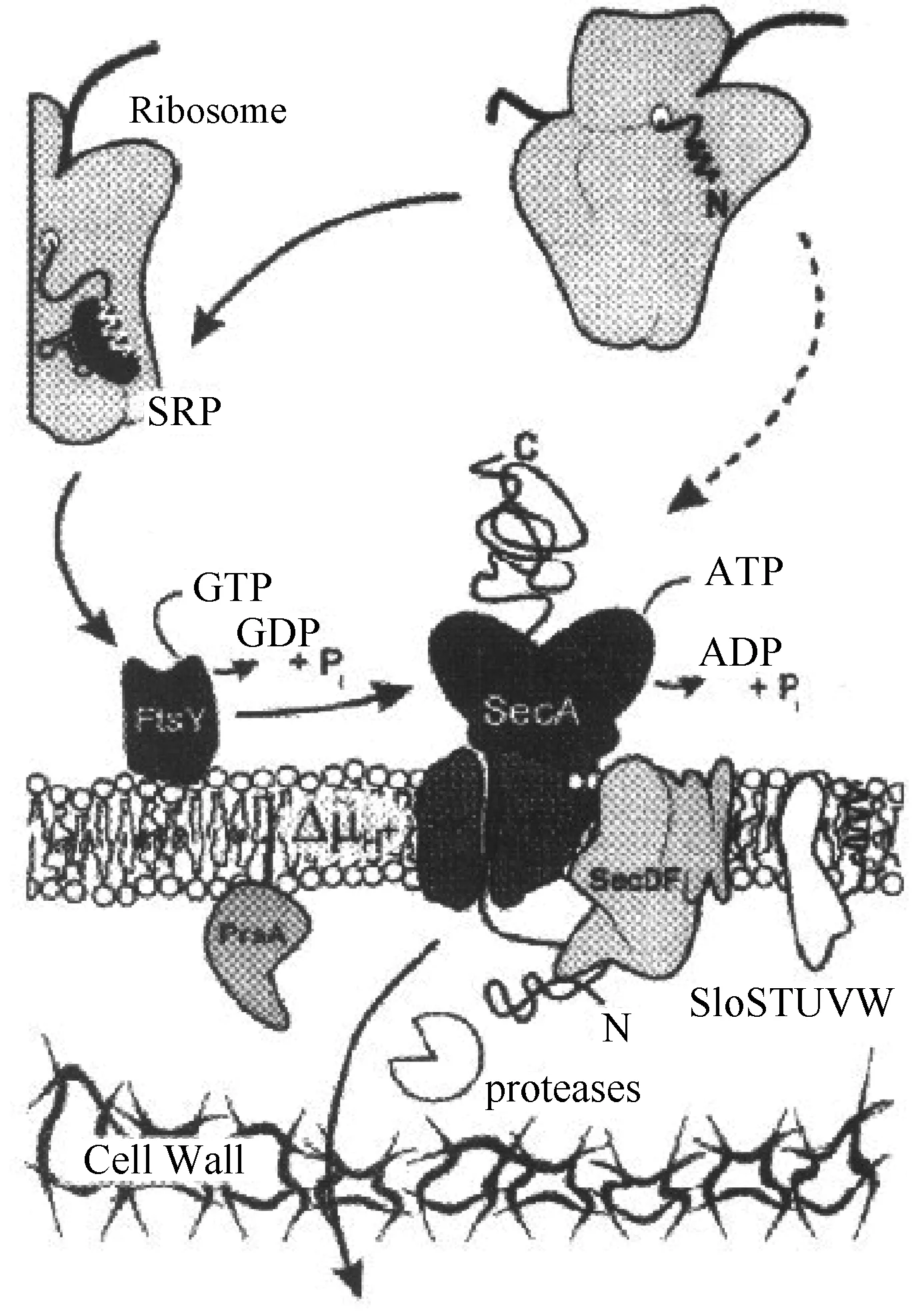
图1 枯草芽孢杆菌蛋白质Sec途径转移机制
Sec途径是枯草芽孢杆菌分泌胞外蛋白质的主要途径,也是酶制剂工业化生产的主要途径,但经Sec途径可成功分泌的外源蛋白较少,而且表达量并不高[26]。主要原因如下:
(1)效率较低。Sec 转运途径只能将以非折叠状态即多肽链形式存在的分泌蛋白前体从细胞质转运到细胞膜外,但在细胞质中已经折叠完全的蛋白质不能通过该途径转移;
(2)蛋白降解。经sec途径转运的外源蛋白在转移到膜外后才进行折叠,但这一过程比较慢,有时来不及折叠就会被蛋白酶降解,降低了外源蛋白的表达量。
(3)错误折叠。王光强等尝试用不同的Sec信号肽来引导分泌表达耐热乳糖酶BgaB,均分泌失败,利用python语言编写的程序分析表明,细胞质蛋白质的N端较容易折叠成高级结构,导致外源蛋白无法进入分泌途径而失败。
(4)膜定位能力差。
Tat(twin-arginine translocation)转运途径。2000年Jongbloed等通过功能基因组的分析发现Tat转运途径,这也是研究较为清楚的另一种蛋白分泌途径,与Sec转运途径有很大区别的是,该途径可以将折叠紧密的蛋白或者多亚基酶复合物直接分泌到胞外,而Sec转运途径只能转移多肽链型式的蛋白。除枯草芽孢杆菌外,如一些植物叶绿体和细菌如大肠杆菌等均存在该途径[27]。
Tat途径的典型特征为分泌蛋白信号肽中两个精氨酸残基相邻,经此途径分沁的蛋白大多数都带有联精氨酸信号肽。Tat途径由TatA(3个TatA/B/E的同源基因)和TatC(2个TatC的同源基因)组成[28],其中3个TatA/B/E同源基因的跨膜和两亲性双螺旋在蛋白分泌过程中起着关键性的作用。首先,蛋白信号肽双精氨酸前体与TatC相连,随后TatC和TatB再相连,在具有跨膜PH梯度下,再连入TatA,最后,TatB调节转移折叠好的基质从TatC转移到Tat孔,具体途径如图2[29]。
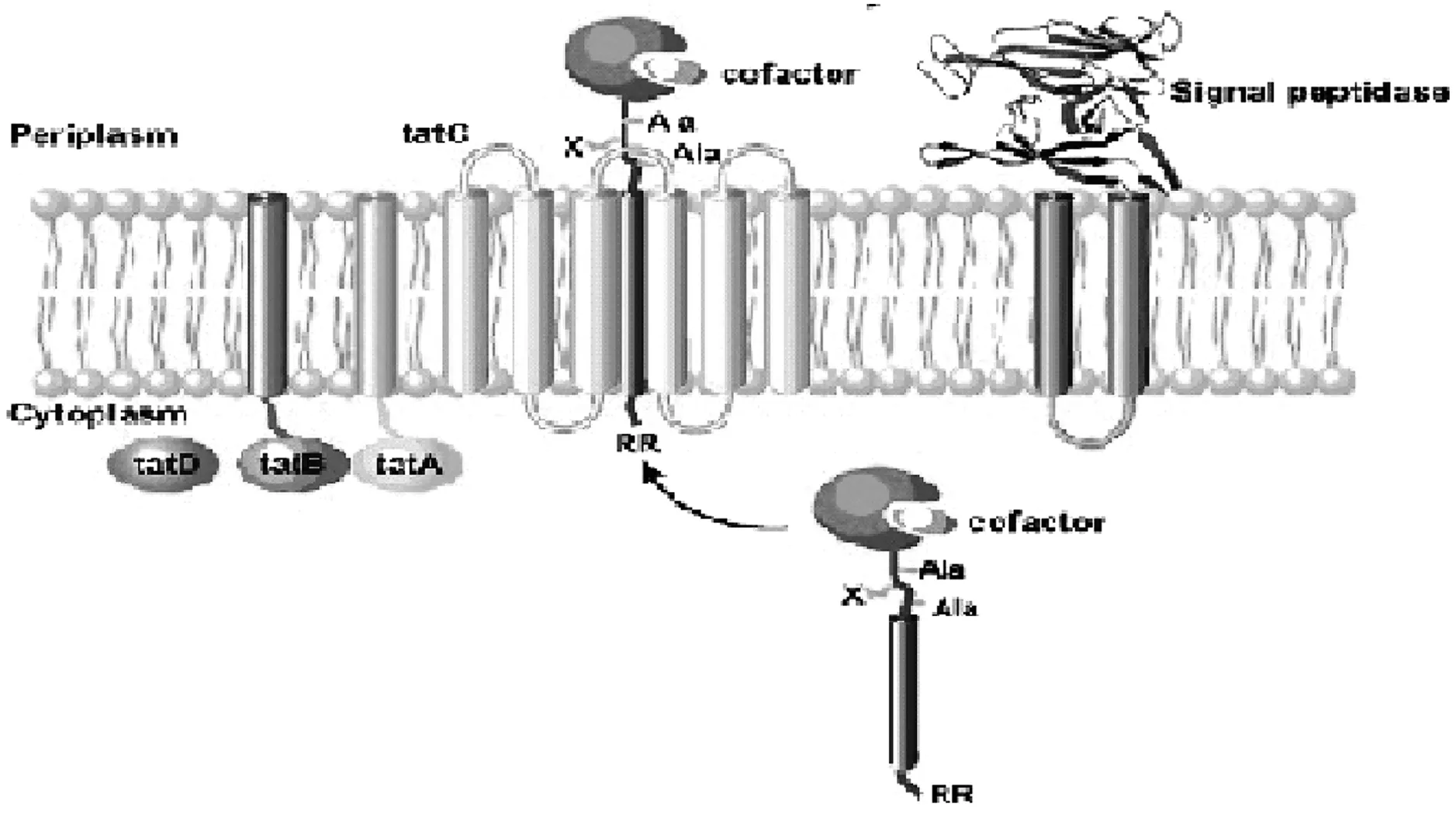
图2 枯草芽孢杆菌蛋白质Tat途径转移机制
Tat途径的研究多集中于大肠杆菌上,berk等发现大肠杆菌中Tat基质在辅助因子作用下可以转运折叠蛋白,Santini和Thomas分别证明绿色荧光蛋白可以在折叠状态下通过Tat途径在大肠杆菌中转移至胞外。对枯草芽孢Tat途径的研究较少,主要集中在与大肠杆菌的Tat途径的对比上,只有很少的蛋白质可以通过Tat途径分泌,目前仅磷酸二酯酶(PhoD)和未知功能蛋白质YwbN是枯草芽孢杆菌Tat途径分泌至胞外的蛋白质[30]。
3 枯草芽孢杆菌表达系统的研究方向
近年来,枯草芽孢杆菌表达系统不断地发展,在外源基因克隆表达领域和工业生产上都得到了广泛应用,很多外源基因经枯草芽孢杆菌表达系统得到高效表达[31],但同时枯草芽孢杆菌表达系统仍存在以下问题,影响了其进一步的应用发展。
(1)分子克隆效率低。枯草芽孢杆菌中很少有可以自发形成感受态的菌株,而且形成感受态持续的时间也很短,现有的质粒载体不稳定,经常会发生结构或者分离方面的问题,使质粒结构改变甚至丢失。
(2)蛋白酶降解。枯草杆菌中存在大量表达和分泌蛋白酶,比如碱性蛋白酶、中性蛋白酶、金属蛋白酶等。这些蛋白酶没有底物专一性,外源蛋白分泌到胞外后,会被这些蛋白酶降解,从而降低产率。现在通过研究已经构建了许多蛋白酶失活的突变菌株,如WB600和WB700,可以在一定程度上解决蛋白酶降解的问题,提高产率。
(3)枯草芽孢杆菌表达系统中有大量的信号肽(如Sec途径有148个,Tat途径有27个),这些信号肽对不同的蛋白有不同的引导分泌效率,目前研究中一般通过高通量筛选的方法来确定目的蛋白的适配信号肽,无法进行有效预测。
(4)真核蛋白表达量低[32]。因此在今后的研究中,一方面需要优化胞内蛋白酶质量控制体系,提高目的蛋白产率,另一方面要加强信号肽分泌机制研究,并构建目的蛋白的信号肽筛选系统,提高表达效率。
