植物源性抗真菌肽研究进展
2021-03-10王小咪吕欣然白凤翎
王小咪 吕欣然 白凤翎
(1. 渤海大学食品科学与工程学院,辽宁 锦州 121013;2. 生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心,辽宁 锦州 121013;3. 国家鱼糜及鱼糜制品加工技术研发分中心,辽宁 锦州 121013)
真菌是一类可引起植物、动物和人类疾病的真核微生物。根据美国农业部[1]估计,世界25%粮食作物在生长或贮藏期间受到霉菌污染。最常用的控制真菌技术是热处理和酸基防腐剂处理,控制烘焙产品中霉菌生长常采用紫外线或红外辐射[2]。一些果汁和茶饮料常采用巴氏杀菌技术和通过添加丙酸钙、脱氢乙酸钠等控制霉菌污染。传统抗真菌剂被认为是导致真菌耐受性的主要原因,因此,新型生物类抗真菌剂的研发与制备备受关注。
抗菌肽是一类广泛存在于自然界具有抗菌活性的小肽类物质,许多抗菌肽具有很强的抗真菌活性,即抗真菌肽(Antifungal peptides, AFPs)。根据结构将AFPs分为4类,第1类是在疏水环境中形成α-螺旋的线性肽,简称α-螺旋肽;第2类是β-片状结构的肽,简称β-片状肽;第3类是半胱氨酸稳定的α-螺旋和β-片层折叠肽,简称CSαβ肽;第4类是包含大量的特定残基和随机结构小肽的线性肽,简称线性肽[3]。依据来源将其分为植物源性、动物源性和微生物源性AFPs。植物源性AFPs来源植物的块茎、种子等植物组织中,种类丰富,具有广谱性、高效性、安全性等特点。
1 植物源性抗真菌肽
目前,学者们已从不同植物中分离纯化出一些具有活性的AFPs,表1概括了近20年有关植物源AFPs结构、抑制对象及作用机制等内容。
1.1 α-螺旋肽
α-螺旋肽主要包括脂质转移蛋白、α-发夹蛋白和蜕皮素类抗菌肽。脂质转移蛋白(Lipid transfer protein, LTP) 是一类由4个α-螺旋组成的小分子量碱性蛋白质。LTP来源广泛,已从玉米、大麦、菠菜、拟南芥、洋葱、豇豆、水稻等多种植物中获得了具有抗真菌活性LTP。Ace-AMP1是一种分离于洋葱种子的LTP,对番茄早疫病菌、棉花枯萎病菌和棉花黄萎病菌等多种植物真菌具有高效的拮抗活性[26]。Gonorazky等[27]从向日葵种子中分离出一种非特异性LTP (nsLTPs),命名为Ha-AP10,具有抗茄病镰刀菌(Fusariumsolanum)的活性,可通过静电吸附与真菌膜上的磷脂相互作用,渗透进入真菌孢子,达到抗真菌作用。
α-发夹蛋白是一类含有特征性半胱氨酸的环螺旋肽。Nolde等[8]从稗草中分离出一种α-发夹蛋白,命名为EcAMP1,其可进入茄病镰刀菌细胞内并积累在分生孢子(液泡或脂质体)囊泡中,从而抑制镰刀菌的生长。此外,Slavokhotova等[7]从金缕草种子中得到的Sm-AMP-X是一种有效的抗真菌α-发夹蛋白,其在微摩尔范围内抑制了曲霉(Aspergillus)或尖孢菌霉(Fusariumoxysporium)等植物病原真菌的分生孢子萌发和菌丝伸长。
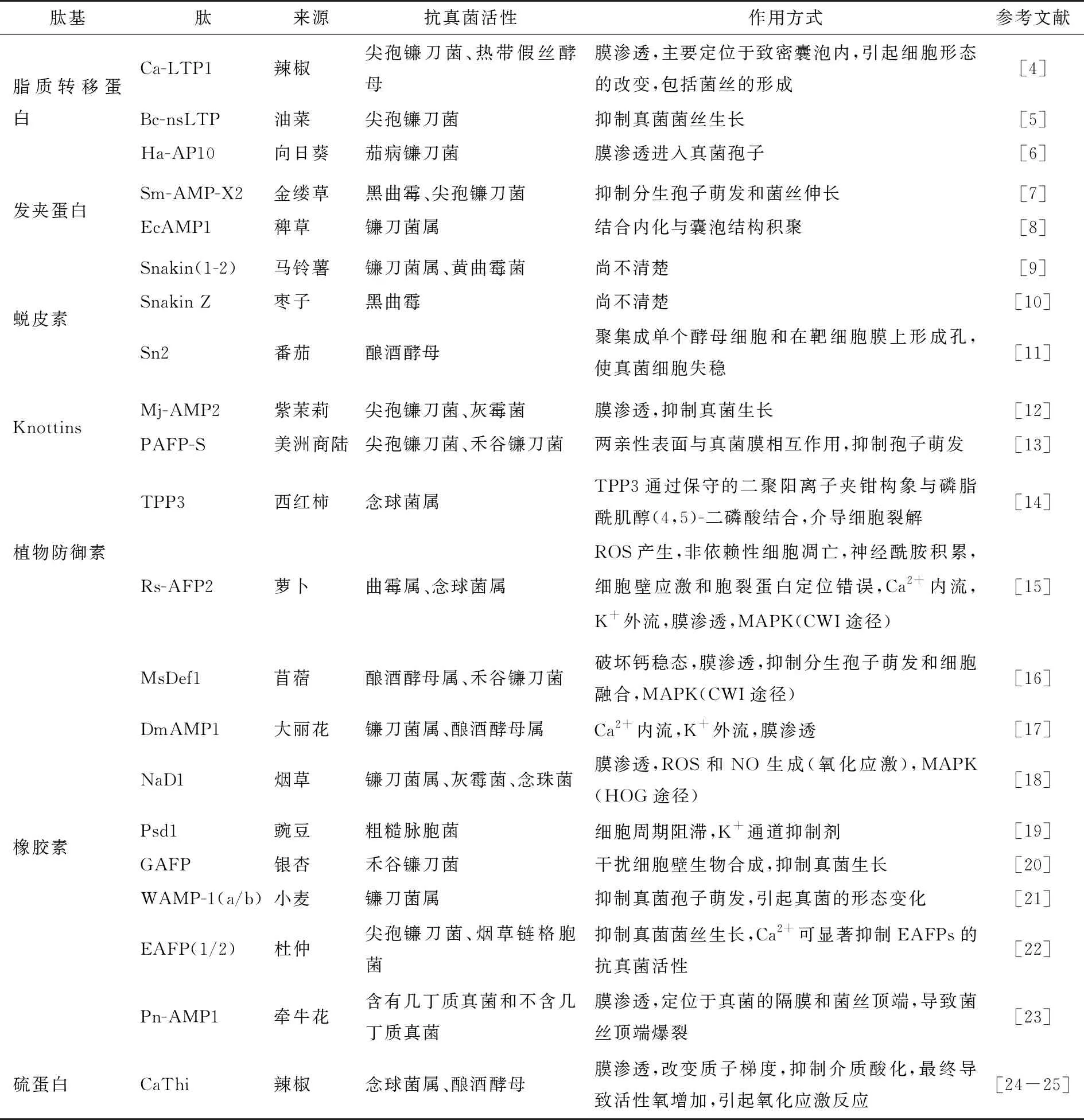
表1 植物源AFPs的抗菌活性和作用方式
蜕皮素是一类由1个双α-螺旋结构、6个稳定的二硫键和1个必需的C-末端(可确保肽的生物活性和稳定性)组成的碱性蛋白,分子量约7 kDa。Segura等[9]从马铃薯块茎中首次分离出一种蜕皮素,命名为Snakin-1,该肽含有63个氨基酸残基,其在低浓度(<10 μmol/L)时对马铃薯和其他植物中的细菌和真菌病原体(如茄病镰刀菌)均具有拮抗活性。此外,马铃薯块茎组织发生降解或受到侵袭时,其能表达一种肽Snakin-2,与Snakin-1序列同源性仅为38%。虽然Snakin-2和Snakin-1序列具有显著性差异,但其抑菌谱相似,对镰刀菌属、黑曲霉(Aspergillusniger)和酿酒酵母(Saccharomycescerevisiae)等几种植物病原菌均具有拮抗活性,Snakin-2的作用靶点是在细胞膜上形成孔洞。其他植物中还发现了蜕皮素Snakin-Z[10]和Sn2[11]。
1.2 β-片状肽
β-片状肽是由多个二硫键组成的稳定结构肽,具有两亲性,可与真菌细胞膜相互作用。Knottin肽是一种β-片状肽,含有30个半胱氨酸残基,具有稳定的“打结”拓扑结构。Gao等[13]从美洲商陆种子中分离并克隆了一种新的抗真菌肽PAFP-S,序列分析表明PAFP-S属于Knottin型抗菌肽,具有广谱的抗真菌活性,如对尖孢镰刀菌(Fusariumoxysporum)、禾谷镰刀菌(Fusariumgraminearum)和稻瘟病菌(Magnaporthegrisea)均具有较强的抑制作用。此外,分离自紫茉莉种子中的抗真菌肽Mj-AMP1和Mj-AMP2与PAFP-S具有相同的半胱氨酸结构[12],具有57%的序列同源性,并分别含有36,37个氨基酸残基,其中4个氨基酸残基不同,3个差异发生在C-末端发夹结构域,此结构域代表了1个β-发夹序列,称为γ-核心,常被认为是肽活性的必要结构。
1.3 CSαβ肽
植物防御素是一类CSαβ肽,富含二硫键,分子量在5 kDa 左右,由45~54个氨基酸组成,其中含有8个保守的半胱氨酸残基,其三维结构呈小球形,包括1个α-螺旋和3个反平行的β-折叠片层结构,存在于多种植物组织(种子)中,是生物免疫系统中的重要调节因子。
胡萝卜种子的植物防御素Rs-AFP2是最具特征性的防御素之一,对丝状真菌具有较强的抑制活性[28]。Thevissen等[15]研究发现,Rs-AFP2可作用于白色念珠菌(CandidaAlbicans)细胞壁葡萄糖神经酰胺(GlcCer),增大细胞壁压力,使胞裂蛋白(septin)错误定位和神经酰胺积累,抑制其菌丝生长。苜蓿防御素MsDef1也有类似GlcCer抑制禾谷镰刀菌生长的作用[16]。此外,大丽花防御素Dm-AMP1与真菌膜上甘露糖苷二肌醇磷脂酰神经酰胺[M(IP)2C]结合,可增加敏感真菌膜的通透性,导致钙离子和钾离子泄露,达到抑制真菌生长的作用效果[17]。植物防御素不但可以作用于真菌细胞壁和细胞膜,还可作用于胞内的一些靶点。
1.4 线性肽
线性肽主要包括具有特定氨基酸残基的小分子橡胶素和硫蛋白,其具有α和β随机结构。橡胶素以橡胶蛋白命名,通常由29~45个氨基酸残基组成,分子量约为4.7 kDa,含有6,8,10个半胱氨酸,其中8个半胱氨酸是最常见的形式。橡胶素具有几丁质结合特性,因此其作用方式是干扰细胞壁生物合成。Huang等[20]从银杏叶中分离的一种新型橡胶素银杏内酯GAFP与几丁质相互作用后可抑制多种镰刀菌生长,如禾谷镰刀菌和串珠镰刀菌等。Koo等[23]研究发现,源自日本牵牛花的Pn-AMP1是一种4.3 kDa半胱氨酸/甘氨酸丰富的抗菌肽,对细胞壁中含有几丁质和缺乏几丁质的真菌均具有较强的抑制活性。其能定位于真菌的隔膜和菌丝顶端,并快速渗透进入真菌菌丝中,使菌丝顶端爆裂,内容物泄露,导致真菌死亡。此外,来自芥末的Wj-AMP1[29]、苏铁的Cy-AMP1[30]、纺锤树的Ee-CBP[31]、杜仲的EAFP2[22]和小麦种子中的WAMP-1(a/b)[21]等橡胶素也具有抗真菌活性。
硫蛋白由两个反平行α-螺旋和一个反平行双链β-片组成,由3个或4个二硫键桥连接,形成一个特征稳定的γ折叠,分子量约5 kDa,含有高比例的精氨酸、赖氨酸和半胱氨酸,且在中性pH下带正电荷,广泛分布在大麦、豇豆等多种植物组织中。硫蛋白是疏水性的,通过疏水性和/或极性残基与膜元件的相互作用显示其抗真菌活性,导致真菌细胞膜的溶解[32]。Taveira等[24-25]从辣椒中分离出对念珠菌和酿酒酵母具有抑制作用的抗菌肽CaThi与酿酒酵母质膜成分相互作用,改变质子梯度,抑制介质酸化,增加活性氧,引起氧化应激反应。
综上,目前分离纯化得到的植物源AFPs主要对镰刀菌属、酿酒酵母属、念球菌属及曲霉属等真菌具有拮抗活性,其作用方式主要通过膜渗透作用,其中植物防御素类抗菌肽研究得较为深入。
2 植物源性抗真菌肽的作用机理
目前,已从多种植物中获得具有抗真菌作用的活性多肽。由图1可知,根据作用位点的不同,植物源AFPs作用机理大致可以分为以下3类。
2.1 对真菌细胞壁的作用
真菌细胞壁主要由糖基化蛋白、β-葡聚糖及几丁质等成分组成,是AFPs识别真菌细胞的靶点。植物源AFPs作用方式:① 通过与真菌细胞壁最外层的糖基化蛋白相互作用以抑制细胞壁蛋白质的合成,导致细胞壁不能维持正常的细胞形态,如结构多样化甘露糖蛋白是AFPs准确识别病原菌的靶点。Ibeas等[34]研究表明,烟草抗菌肽osmotin可与细胞壁甘露聚糖相互作用,以促进跨细胞壁的扩散,诱导信号级联并激活酿酒酵母中的信息素反应MAPK和应激反应途径,致使真菌细胞受到损伤。② 通过抑制真菌细胞壁中葡聚糖及几丁质的合成来破坏真菌细胞壁的合成,达到杀菌目的。α-硫蛋白是源自大麦和小麦的一种抗菌肽,可与细胞壁几丁质和β-1,3-葡聚糖结合,使真菌生长受到抑制。一些植物类thaumatin型肽被证明具有β-1,3-葡聚糖酶活性,可以水解细胞壁中的葡聚糖成分,使细胞壁合成受阻,导致真菌细胞溶解和死亡。橡胶蛋白中包含几丁质结合域,可与含有几丁质的真菌相互作用,抑制几丁质的合成,从而破坏细胞壁的完整性。与橡胶蛋白类似的hevein型肽也能抑制含有几丁质的真菌生长。
2.2 对真菌细胞膜的作用
细胞膜是真菌细胞的第二道防线,也是AFPs普遍的作用靶点。大多数阳离子型AFPs都可作用于真菌细胞膜,从而达到抑菌作用。细胞质膜带有负电荷,有助于AFPs通过静电作用吸附于细胞膜上,使细胞膜发生去极化反应,增大细胞膜的通透性,使其形成孔洞,导致钾离子以及细胞内容物外渗而死亡。结合图1可知,AFPs可通过“桶式模型”“环状孔道模型”“毯式模型”作用于真菌细胞膜。但在实际抗真菌时,植物源AFPs可能同时采用几种不同机制或者各种机制渐次发生的方式破坏细胞膜的结构,从而实现杀菌的目的[35]。

图1 植物源AFPs的作用机制[33]
2.2.1 桶式模型 AFPs与细胞膜通过静电作用吸附后,AFPs积聚形成多聚体,垂直插入细胞膜磷脂层内,形成类似于木桶的离子孔道,导致细胞内容物和钾离子外流,细胞膜分布不均,引起细胞裂解[36]。许多植物防御素的作用方式是桶式模型,其抗菌作用可分为3个阶段:① 通过静电作用与带负电荷的真菌膜脂层结合;② 作用于膜上形成稳定的多个通道,带正电荷的防御素分子或其多聚体则与真菌膜上带负电荷的磷脂头部和水分子相互作用,显著增加生物膜的通透性。③ 通道形成后,防御素进入细胞内的同时,其他胞外分子也伴随进入(如肽、蛋白质或无机离子),而靶细胞的重要物质(如盐离子和大分子)渗出,细胞内容物外泄致使靶细胞发生不可逆损伤而死亡。
2.2.2 环状孔道模型 环状孔道模型也称虫孔模型,是指AFPs的疏水部分与磷脂膜疏水头部相结合,随后AFPs移动引导细胞膜磷脂移动。AFPs疏水端和磷脂头部一直相结合,使细胞膜表面疏水区域产生裂缝,磷脂单分子层逐渐向内流动弯曲,形成孔道,改变细胞膜稳定性,使细胞无法维持正常的渗透压,进而凋亡。与桶式模型不同的是,AFPs始终与磷脂膜头部相结合,两者共同形成跨膜孔道。如源自番茄中的一种抗菌肽Sn2,其对酿酒酵母具有拮抗活性,主要作用方式是通过在靶细胞膜上形成孔,使真菌细胞失稳,进而凋亡[11]。
2.2.3 毯式模型 毯式模型是AFPs平行排布于细胞膜表面,在电荷作用下以地毯形式覆盖在磷脂层表面,随着AFPs积聚增多,被覆盖区域细胞膜稳定性下降,逐渐变形弯曲,最后导致细胞膜瓦解。Alfred等[37]研究发现,puroindolines是13 kDa的小麦胚乳碱性蛋白,具有丰富的色氨酸结构域和5个二硫键。其在真菌膜上产生孢子,可能是通过地毯状的机制抑制真菌活性[37]。由于毯式模型主要是在电荷作用下吸附AFPs与真菌膜相互作用,而大部分AFPs都是阳离子性多肽,所以该模型基本上都是与其他作用机制一起作用。
2.3 与真菌细胞内靶点的作用
一些AFPs与真菌细胞壁和细胞膜相互作用后,还可与胞内多种靶点作用,包括启动胁迫基因、絮凝细胞内含物、抑制DNA或蛋白质合成、调控线粒体等细胞器代谢等方式来发挥抗菌活性。通常内化过程发生前,AFPs先与细胞壁和细胞质膜相互作用,因此,AFPs作用于细胞内靶点时,也与真菌细胞壁或细胞膜发生作用。烟草防御素NaD1和豌豆防御素PsD1可进入真菌细胞并与细胞内靶点相互作用。NaD1是烟草花组织的一种高水平表达防御素,主要作用于真菌细胞质,使其呈颗粒状,抑制尖孢镰刀杆菌、灰霉菌和念珠菌等真菌生长[18]。PsD1的抗真菌活性基于阻断细胞核上的周期蛋白合成,终止真菌细胞周期,导致真菌细胞凋亡[19]。
综上,AFPs可通过作用位点的不同使真菌细胞生物活性降低或直接导致细胞裂解死亡,以达到抑菌作用。然而,目前关于植物源AFPs的作用机制研究主要集中于细胞膜上,其他靶点的作用机制还有待深入研究。
3 植物源性抗真菌肽的应用
目前,一些植物源AFPs已被应用于食品和农作物等领域,冯林慧[38]将对黑曲霉具有拮抗活性的大豆多肽GAP以食品添加剂的形式应用到海绵蛋糕中,发现随着GAP添加量的增加,海绵蛋糕的感官特性和理化特性有一定程度的下降,但抗菌特性显著增加。此外,更多的学者利用转基因技术将AFPs转入到小麦、玉米和水稻等农作物中,以提高农作物对病原真菌的抗性。该策略不仅减少了农药的大量使用,且比传统的育种方法效率更高。蒋雯等[39]将源自马铃薯块茎的抗菌肽SN1的基因编码序列克隆至载体pGEX-4T-1上,发现克隆表达的抗菌肽SN1可显著抑制禾谷丝核菌(Rhizoctoniacerealis)和平脐蠕孢菌(Bipolaris)菌丝的生长,其可作为小麦病害抗性育种的潜在基因。Zhang等[40]在玉米泛素启动子的控制下,将源自小麦胚乳的puroindoline蛋白PINA和PINB导入玉米基因中,可使玉米对异旋孢腔菌(Cochliobolusheterostrophus)产生一定的耐受性,并将症状的发生率降低42%。Chen等[41]利用农杆菌介导的基因转移技术,将苜蓿防御素基因(alfAFP)、水稻几丁质酶基因(CHI)及其结合基因(CHI-AFP)导入番茄微生态系统,发现转基因植株对病原菌感染的抗性高于非转基因植株,增强了番茄对灰霉病的抗菌性。
4 结论
植物源抗真菌肽具有来源广泛、结构多样、作用机制复杂等特点,是控制食品和农作物等产品感染真菌的重要资源。目前,不同来源的植物源抗真菌肽研究主要集中于抗真菌肽的分离、表达、纯化及活性分析方面,有关抗真菌肽的作用机制方面的研究还不够深入,同时造成其在食品和农作物等领域的应用产生了局限性。对于存在的这些问题,期望未来更多的研究集中于植物源抗真菌肽的作用机制和应用等方面,使新型、高效、低毒、广谱的植物源性抗真菌肽在农业、食品、医药等领域发挥重要作用。
