Global-Scale Diversity and Distribution Characteristics of Reef-Associated Symbiodiniaceae via the Cluster-Based Parsimony of Internal Transcribed Spacer 2 Sequences
2021-03-05GONGSanqiangCHAIGuangjunSUNWeiZHANGFengliYUKefuandLIZhiyong
GONG Sanqiang, CHAI Guangjun, SUN Wei, ZHANG Fengli, YU Kefu, and LI Zhiyong, *
Global-Scale Diversity and Distribution Characteristics of Reef-Associated Symbiodiniaceaethe Cluster-Based Parsimony of Internal Transcribed Spacer 2 Sequences
GONG Sanqiang1), 2), 3), CHAI Guangjun1), SUN Wei1), ZHANG Fengli1), YU Kefu4), and LI Zhiyong1), *
1),,,200240,2),,510301,3)(),511458,4),,530004,
Photosynthetic species belonging to Symbiodiniaceae play important ecological roles in coral reef ecosystems. Nine phylogenetically supportedSymbiodiniaceae clades (each containing multiple genetically distinct molecular types) have been detected in reef-associated animals and in seawater or sediments near coral reef areas. Although major advances in research on Symbiodiniaceae diversityhave been made, information on the global-scale diversity and distribution characteristics of Symbiodiniaceae clades/types is still limited or must be expanded because most studies on Symbiodiniaceae have mainly focused on a limited number of hosts associated with Symbiodiniaceae or coral reef areas. Here, the diversity and distribution characteristics of Symbiodiniaceae data derived from reef-associated animals and seawater or sediments near coral reef areas were analyzed using 3899 qualifiedSymbiodiniaceae internal transcribed spacer 2 (ITS2) sequences retrieved from the GenBank database in May 2017. These sequences were annotated as Symbiodiniaceae ITS2 typessequence alignment against database of Symbiodiniaceae ITS2 types. Results showed that photosynthetic Symbiodiniaceae ITS2sequences could be annotated as 119 Symbiodiniaceae ITS2 types within nine clades (A–I), and Symbiodiniaceae ITS2 types mainly inhabited coral reefs between 30˚N and 30˚S. Symbiodiniaceae ITS2 types could be associated with multiple host taxa. Interestingly, these results implied that the distribution of Symbiodiniaceae ITS2 types was distinct in the Atlantic Ocean, the Red Sea, the Mediterranean Sea, and the Indo-Pacific Ocean. Although future field research should be performed to verify our results, this study provided a broad perspective of the diversity of Symbiodiniaceae clades/types on a global scale.
Symbiodiniaceae; diversity; reef-dwelling animals; coral reefs; global scale
1 Introduction
Photosynthetic Symbiodiniaceae plays important roles in the establishment of the most diverse and productive coral reef ecosystems in oligotrophic marine waters (Coffroth., 2006). Symbiodiniaceae is mainly associated with metazoans (, reef-building corals, non-reef-build- ing corals, flatworms, and sponges) and foraminifera by providing them nutrients. In turn, these reef-associated metazoans and foraminifera create a microenvironment and supply some metabolic substrates that allow Symbiodiniaceae to survive at high densities (Blackall., 2015). Previous studies revealed the environmental factors that influence the diversity, geographic distribution, and mutual association of Symbiodiniaceae with different hosts (Baird.,2009; Takabayashi., 2012; Yamashitaand Koike,2013). The breakdown of Symbiodiniaceae–coral associations contributes to remarkable declines in coral reef covers worldwide (Jones, 2008; De’Ath., 2012). Consequently, the diversity and geographic distribution of Symbiodiniaceae should be monitored to predict the fate of corals and coral reef ecosystems under marine environmental changes.
Initially, Symbiodiniaceae members were considered a single pandemic species based on morphological analyses (Freudenthal, 1962). Subsequently, our understanding of the diversity of Symbiodiniaceae was significantly changedbecause of sequence-based molecular analyses (Baker, 2003;Coffroth and Santos,2005). At present, Symbiodiniaceae taxa aredivided into nine phylogenetically supported clades (A–I), and several Symbiodiniaceae clades have been formally described as a genus based on their genetic and ecological characteristics ([clade A],[clade B],[clade C],[clade D],[clade E],[clade F], and[clade G]; Lajeunesse., 2018). Each clade or genus of Symbiodiniaceae includes a number of informally described molecular types (Pochon., 2014; Lajeunesse., 2018).
The diversity and geographic distribution of Symbiodiniaceae taxa and their associations with reef-associated metazoans and foraminifera have been mainly explored with different molecular markers,., nuclear large subunit (LSU), internal transcribed spacer regions (ITS2 and ITS1), chloroplast large subunit (), cytochrome oxi- dase b, and noncoding chloroplastregion (LaJeunesse., 2011). However, the ITS2 region currently remains the most commonly employed DNA marker for assessing the diversity of Symbiodiniaceae ITS2 types (Franklin., 2012; Tonk., 2013; Arif., 2014; Green., 2014; Prada., 2014; Quigley., 2014; Thomas., 2014; Cunning., 2015; Reimer., 2017; Ziegler., 2017).
Baker (2003) and van Oppen(2009) reviewed the global-scale diversity and geographic distribution of Sym-biodiniaceae among corals at the clade level and suggested that Symbiodiniaceae clades A–D are common to corals living in the Atlantic Ocean, whereas Symbiodiniaceae clad- es C and D are dominant in corals inhabiting the tropical Indo-Pacific Ocean.
Lajeunesse (2002, 2003), and Lajeunesse(2005) analyzed the geographic distributions of Symbiodiniaceae typeswithinclades C and B in cnidarians (especially reef- building corals) living between the Atlantic Ocean and the Indo-Pacific Ocean at the type level. Franklin(2012) built a global Symbiodiniaceae type data set with an online interface for users to explore and utilize global geospatial and ecological data onSymbiodiniaceae–host symbioses. Tonk(2013) reviewed studies on the di- versity of Symbiodiniaceae in invertebrates from the Great Barrier Reef. In addition, previous studies revealed that corals can change or exchange their symbiotic Symbiodiniaceae communities, and Symbiodiniaceae living in seawater or sediments near coral reef areas likely serve as an open pool from which corals can acquire algae when they are under environmental stress (Fautin and Buddemeier, 2004; Keshavmurthy., 2014; Hughes., 2017). By contrast, studies have reported stable Symbiodiniaceae communities in corals, suchas–Symbiodiniaceae associations, which are stable in the Indo-Pacific in broad geographical ranges and temperature gradients and through bleaching events or after transplantation (Baker, 2003).
Furthermore, physiological differences in Symbiodiniaceae clades or types have been described. For example, certain Symbiodiniaceae clades or types are exceptionally thermotolerant (., D1, D1–4, C15, and A3), whereas others are thermosensitive (., C3, C7, B17, and A13;Stat., 2013; Tonk, 2013; Hughes., 2017). These data suggest that the diversity and distribution of Symbiodiniaceae may depend on biotic (., host species) and abiotic (., temperature, light, nutrients, and geographic affinities) factors (Baker, 2003; Prada., 2014).
Although advances in the diversity and distribution of Symbiodiniaceae have been made, global-scale knowledge on Symbiodiniaceae clades/types is still limited because most previous studies focused on a limited number of hosts associated with Symbiodiniaceae or coral reef areas. In addition, Baker’s work (2003) should be expand- ed. Baker (2003) provided a global perspective of the di- versity and distribution of Symbiodiniaceae based on nuclear LSU gene sequences of reef-building corals. However, the diversity of Symbiodiniaceae has been analyzed in reef-building corals and other metazoans (., non-reef-building corals, flatworm sand sponges), foraminifera, and seawater or sediments near coral reef areas mainly based on the Symbiodiniaceae ITS2 region (Franklin., 2012; Tonk., 2013; Arif., 2014; Green., 2014; Prada., 2014; Quigley., 2014; Thomas., 2014; Cunning., 2015; Reimer.,2017; Ziegler., 2017).
This study aimed to provide a broad global perspective of the diversity and distribution of Symbiodiniaceae by annotating the Symbiodiniaceae ITS2 sequences derived from reef-associated metazoans and foraminifera and from seawater or sediments over 28 coral reef areas worldwide in the GenBank database as Symbiodiniaceae ITS2 typessequence alignment against database of Symbiodiniaceae ITS2 types in accordance with previously described methods (Arif., 2014).
2 Materials and Methods
2.1 Retrieval of Symbiodiniaceae ITS2 Sequences from GenBank
We carried out a systematic search in the NCBI nucleotide database in order to obtain ITS2 sequences of Sy- mbiodiniaceae ITS2 in May 2017 using the string ‘(Symbiodiniaceae and ITS2) and (Symbiodiniaceae and internal transcribed spacer 2)’. The GenBank-formatted ITS2 sequences that matched the string were downloaded from the NCBI nucleotide database and saved as ITS2.GB, which was used as raw data source for the subsequent analyses. Only the Symbiodiniaceae ITS2 sequences derived from the samples of different hosts, seawater, or sediment (including reef sand) were included. In addition, the term ‘host-associated Symbiodiniaceae data’ referred to Symbiodiniaceae ITS2 sequences derived from hosts (host in- formation from GenBank), and the term ‘free-living Symbiodiniaceae data’ corresponded to Symbiodiniaceae ITS2 sequences obtained from seawater or sediment samples.
2.2 Quality Control and Bioinformatic Analysis of Symbiodiniaceae ITS2 Sequences
Symbiodiniaceae ITS2 sequences from the NCBI nucleotide database were subjected to quality control and bioinformatic analysis with modified pipeline and scripts provided by Arif(2014). In brief, Symbiodiniaceae ITS2 sequences were removed if they had a length of <150 nucleotides (nt), contained >6 ambiguous bases, or had a homopolymer length of >6. Chimeric sequences were iden-tified and removed using chimera.uchime and remove.seqs scripts against a database of Symbiodiniaceae ITS2 types (two databases supplied by Arif, 2014, and Cunning, 2017, were merged).
Symbiodiniaceae ITS2 sequences were first clustered into different clades by applying an 85% cutoff value withthe get.oturep script (Arif., 2014) and BLASTn (Altschul., 1990) against the custom database of Symbio- diniaceae ITS2 types because of difficulties in aligning these sequences. Symbiodiniaceae ITS2 sequences belong- ing to different clades were aligned separately with MUSCLE. The aligned Symbiodiniaceae ITS2 sequences were further clustered into operational taxonomic unit (OTU) at a 97% cutoff value. The OTU that represented fewer than three Symbiodiniaceae ITS2 sequences was removed. The most abundant sequence from each OTU was chosen as a representative sequence and annotated as a Symbiodiniaceae ITS2 type with BLASTn (Altschul., 1990) against the custom database of Symbiodiniaceae ITS2 types based on 97% similarity.
The number ofSymbiodiniaceae ITS2 sequences of dif- ferent Symbiodiniaceae ITS2 types was used to record the diversity of hosts or coral reef areas from where Symbiodiniaceae ITS2 sequences were derived.
Rarefaction curve analysis was performed using past3 software (http://folk.uio.no/ohammer/past/) to investigate whether the number ofSymbiodiniaceae ITS2 sequences exceeded the point where the rarefaction curve reached its asymptote.
The annotated ITS2 sequences of Symbiodiniaceae typeswere aligned with MUSCLE in MEGA 6.06 (Tamura., 2013) and trimmed to have an equal length (152bp). Phy- logenetic trees of ITS2 sequences were constructed with maximum likelihood (ML) and neighbor joining (NJ) me- thods in MEGA 6.06. The reliability of internal branches was assessed with the bootstrap method with 1000 replicates. The NJ trees were nearly identical to the corresponding ML trees; thus, only the ML trees were addressed further in this study.
The number ofSymbiodiniaceae ITS2 sequences in different Symbiodiniaceae ITS2 clades/types among different hosts or coral reef areas was used to perform a principal component analysis (PCA) of the community structureCanoco software for Windows 4.5 (Microcomputer Power, NY, USA). The PCA results were assessed with an MCPP test in Canoco for Windows 4.5 in accordance with the manufacturer’s instructions (Ter Braak and Smilauer, 2002).
3 Results
3.1 Diversity of Symbiodiniaceae Based on ITS2 Sequences
In this study, a total of 3899Symbiodiniaceae ITS2 sequences were obtained after they were subjected to qua- lity control and further annotated as 119 Symbiodiniaceae ITS2 types within nine clades (Fig.1). Most Symbiodiniaceae ITS2 sequences belonged to Symbiodiniaceae ITS2 types within clades A, B, C, D, and F, accounting for 96% of the ITS2 sequences and 90% of all Symbiodiniaceae ITS2 types. Specifically, Symbiodiniaceae ITS2 types within clade C showed the highest abundance (2305 ITS2 sequences) and diversity, representing 59% of the total ITS2 sequences and 53% of all the types. Clades A–I had 14, 9, 63, 5, 2, 16, 5, 3, and 2 Symbiodiniaceae ITS2 types, respectively. The diversity of Symbiodiniaceae ITS2types was further analyzed on the basis of rarefaction curveanalysis. The results showed that the number ofSymbiodiniaceae ITS2 sequences deposited in GenBank exceed- ed the point where the rarefaction curve reached its asym- ptote, but the rarefaction curves based on hosts or seawater samples did not reach their asymptotes.
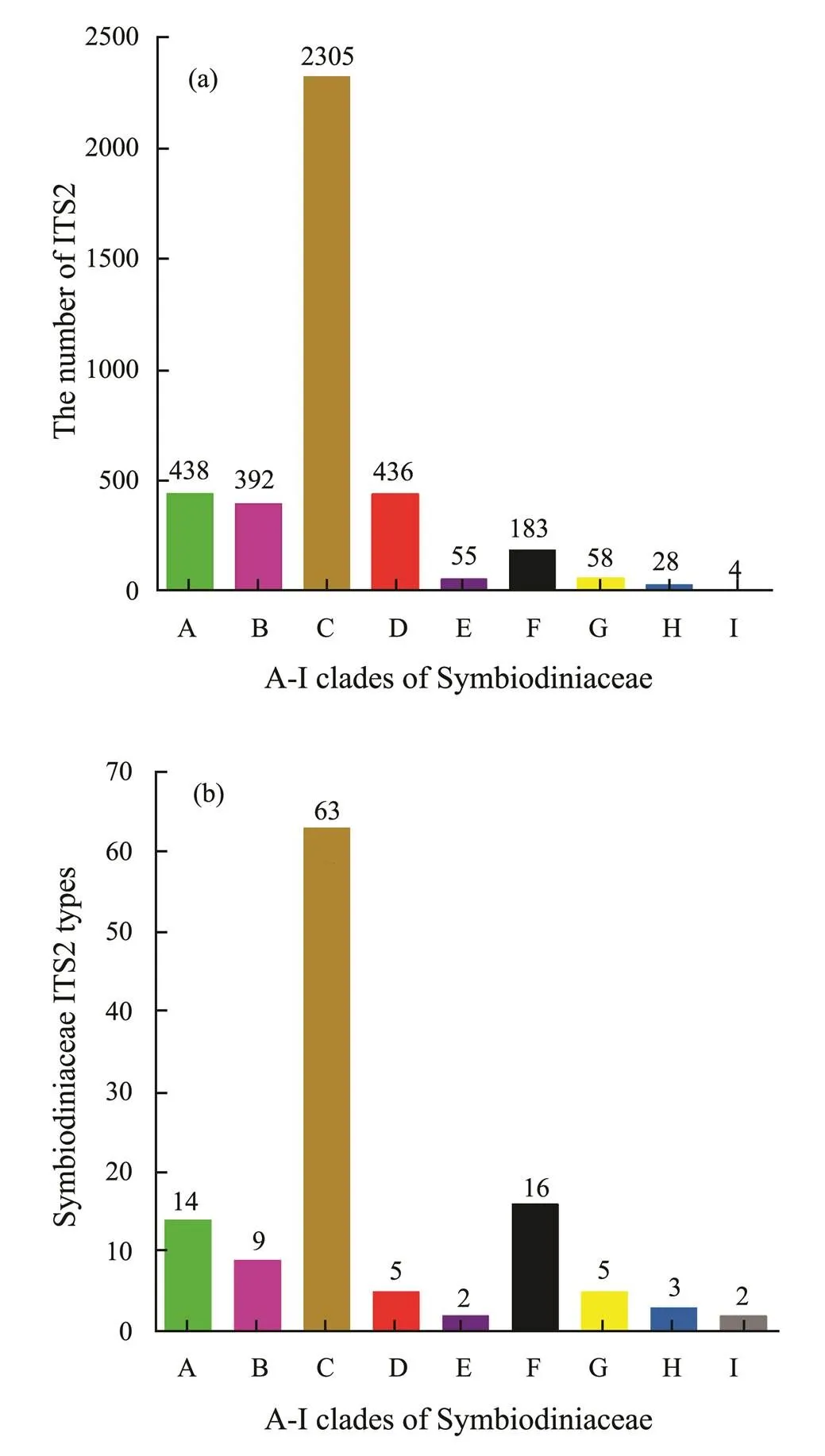
Fig.1 Number of Symbiodiniaceae ITS2 sequences (a) and number of Symbiodiniaceae ITS2 types (b) in different clades (A–I). The number of Symbiodiniaceae ITS2 se- quences was designated among Symbiodiniaceae ITS2 se- quences with different hosts and/or habitats information. The Symbiodiniaceae clades and types were clustered and annotated with an operational taxonomic unit-based method against the database of Symbiodiniaceae ITS2 types.
3.2 Diversity of Hosts Associated with Symbiodiniaceae
The number of Symbiodiniaceae ITS2 sequences and the host information corresponding to the sequences indicated that the majority (91.0%) of ITS2 sequences belonging to Symbiodiniaceae clades A–Iwere derived from metazoans and foraminifera (Fig.2), including Cnidaria (69.1%; reef-building corals, soft corals, jellyfishes, and anemones), Mollusca (11.0%;, bivalves, and nudibranchs), Porifera (0.4%; sponges), and Foraminifera (9.7%). Specifically, the ITS2 sequences belonging toSymbiodiniaceae cladesA, B, C, and D were mainly derived from reef-building corals; the ITS2 sequences belong- ing toSymbiodiniaceae cladesF, H, and I were mainly de- rived from protists(foraminifera); the ITS2 sequences be- longing to Symbiodiniaceae clades G and A were mainly derived from Porifera; and the ITS2 sequences belonging to Symbiodiniaceae clades A, B, C, D, and E were derived from anemones. These results suggested that the hosts as- sociated with Symbiodiniaceae were diverse
In addition, approximately 9.0% of the Symbiodiniaceae ITS2 sequences belonging to Symbiodiniaceae clades A, B, C, D, E, and G were derived from the seawater or se- diment samples collected near coral reef areas (Fig.2).
3.3 Global-Scale Geographic Distribution of Symbiodiniaceae
A broad global perspective of the diversity and distribution of the nine Symbiodiniaceae clades was obtained on the basis of the number of Symbiodiniaceae ITS2 sequences belonging to differentSymbiodiniaceae clades andthe corresponding reef area information (Fig.3). The results showed that the Symbiodiniaceae ITS2 sequences belong-ing to different clades were mainly derived from 28 coral reef areas across the tropical and subtropical oceans between 30˚N and 30˚S in the ocean.Symbiodiniaceae ITS2 sequences belonging to clades A, C, D, and F were widely distributed and derived from coral reef areas acrossthe Atlantic Ocean, the Red Sea, the Mediterranean Sea, and the Indo-Pacific Ocean. By contrast, the remaining Sym- biodiniaceae ITS2 sequences belonging to the other clades were derived from specific oceanic regions. Specifically, Symbiodiniaceae ITS2 sequences belonging to Symbiodiniaceae clade B were mainly derived from coral reef areas in the Atlantic Ocean,., the coral reefs of the Ca- ribbean (Fig.3).
In Figs.4–7,the broad global perspectives of the diver- sity and distribution of the 119 Symbiodiniaceae ITS2 ty- pes belonging to the nine Symbiodiniaceae clades were obtained on the basis of the number of Symbiodiniaceae ITS2 sequences belonging to differentSymbiodiniaceae ITS2 types and the corresponding reef area information, respectively. The results showed that the ITS2 sequences of Symbiodiniaceae ITS2 types A1.1, A1.2, and A1.3 were mainly derived from the hosts living in the Mediterranean Sea and the Rea Sea, the ITS2 sequences of Symbiodiniaceae ITS2 types A2.1 and A2.3 were mainly derived fromthe coral reef areas of the Indo-Pacific Ocean, and the ITS2 sequences of Symbiodiniaceae A6 was mainly derived from the coral reef areas of the Rea Sea (near the Mediterranean). The ITS2 sequences belonging to nine phy- logenetically related Symbiodiniaceae ITS2 types in clade B were mainly derived from coral reefs in the Atlantic Ocean,., the Caribbean (Fig.4). The ITS2 sequences of Symbiodiniaceae ITS2 types F1.1, F1.2, F1.3, and F1.4 were mainly derived from hosts in coral reef areas in the Indo-Pacific Ocean (near Hawaii and New Guinea). Con- versely, the ITS2 sequences ofSymbiodiniaceae ITS2 types F2.1, F2.2, and F2.4 were mainly derived from hosts in the Red Sea and the Mediterranean Sea. The ITS2 sequences ofSymbiodiniaceae ITS2 types F4.1, F4.2, F4.3, and F4.4 were mainly derived from coral reef areas in the Indo-Pacific Ocean (near Guam), and the ITS2 sequencesofSymbiodiniaceae ITS2 types F5.1 and F5.2 were mainly derived from coral reef areas in the Caribbean. The ITS2 sequences ofSymbiodiniaceae ITS2 types in clade I (., types I1 and I2) were mainly derived from foraminifera living near Hawaii (Fig.5). The ITS2 sequences of the Symbiodiniaceae ITS2 types in clade H (., H1.1, H1.2, and H2) were mainly derived from foraminifera in coral reefs in the Indo-Pacific Ocean (near Guam and New Gui- nea) and the Caribbean (H1.2; Fig.6). Notably, the ITS2 sequences ofSymbiodiniaceae C1, C2, and C3 within clade C were derived from numerous animal hosts in different coral reef areas in the Atlantic Ocean, the Red Sea, and the Indo-Pacific Ocean (Fig.7B). These results suggested that Symbiodiniaceae C1, C2, and C3 were globally distributed ITS2 types, whereas the other Symbiodi- niaceae ITS2 types within clade C had limited distributions (Fig.7C).

Fig.2 Symbiodiniaceae clades (A–I) associated hosts and Symbiodiniaceae detected in sediments or seawater. The relative abundance of Symbiodiniaceae clades was calculated on the basis of the number of Symbiodiniaceae ITS2 sequences derived from samples of different hosts and sediment or seawater samples.
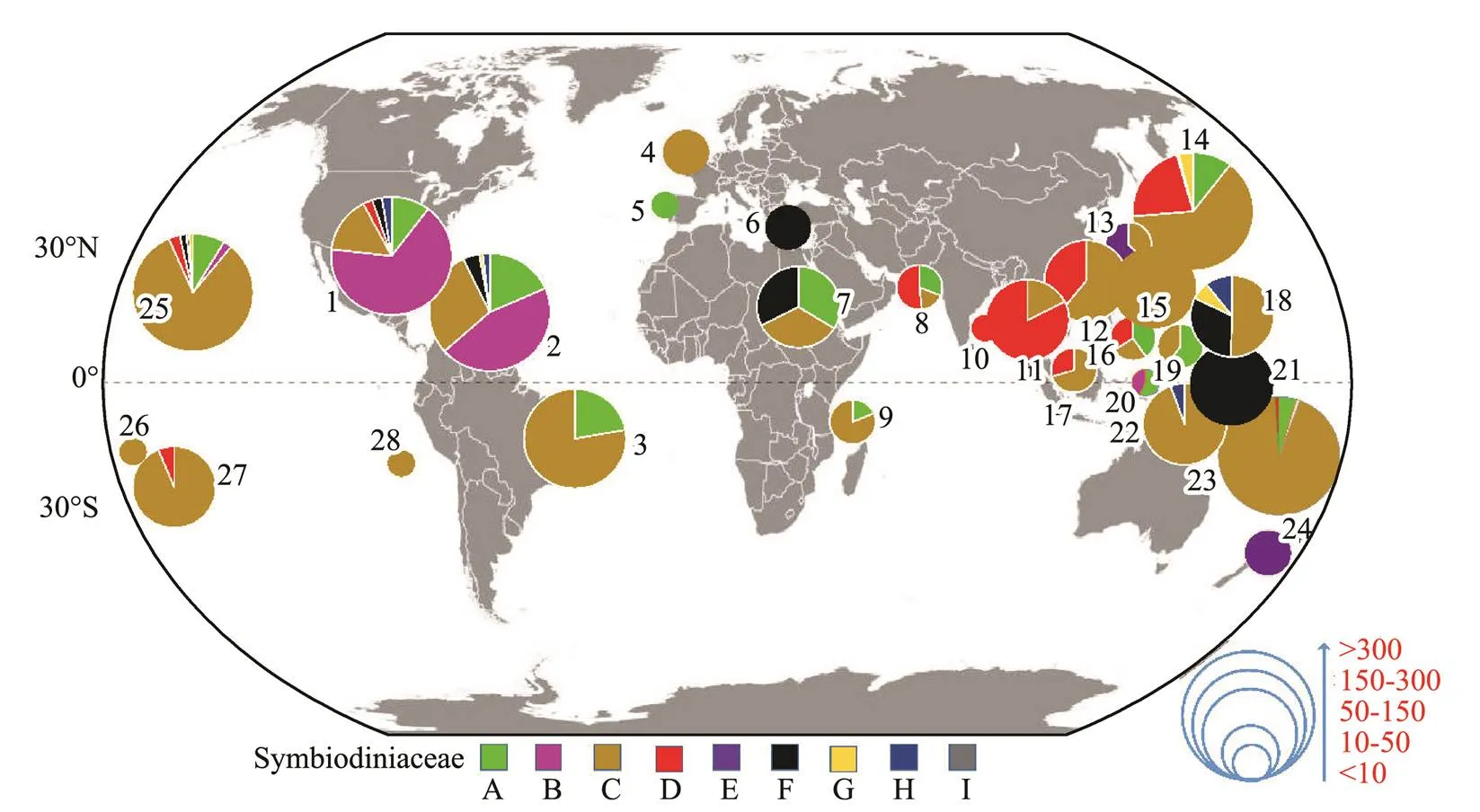
Fig.3 Global geographic distribution of Symbiodiniaceae at the clade level. Numbers (1–28) represent different coral reef areas in or near the following regions: 1, Gulf of Mexico; 2, Caribbean Sea; 3, Brazil; 4, Southampton; 5, Gulf of Alicante (Spain); 6, Mediterranean Sea; 7, Rea Sea; 8, Persian Gulf; 9, Tanzania; 10, Bay of Bengal; 11, Thailand; 12, South China Sea; 13, South Korea; 14, Japan; 15, Taiwan; 16, Philippines; 17, Singapore; 18, Guam; 19, Palau; 20, Indonesia; 21, Papua; 22, New Guinea; 23, Australia; 24, New Zealand; 25, Hawaii; 26, Samoa; 27, Polynesia; and 28, Ascension Island. Pie charts represent the relative abundances of different Symbiodiniaceae clades found in each coral reef area. The pie charts are presented in one of five sizes according to the occurrence records of Symbiodiniaceae in each coral reef area. The global map was drawn with tmap1.8-1 (https://github.com/mtennekes/tmap) in R.
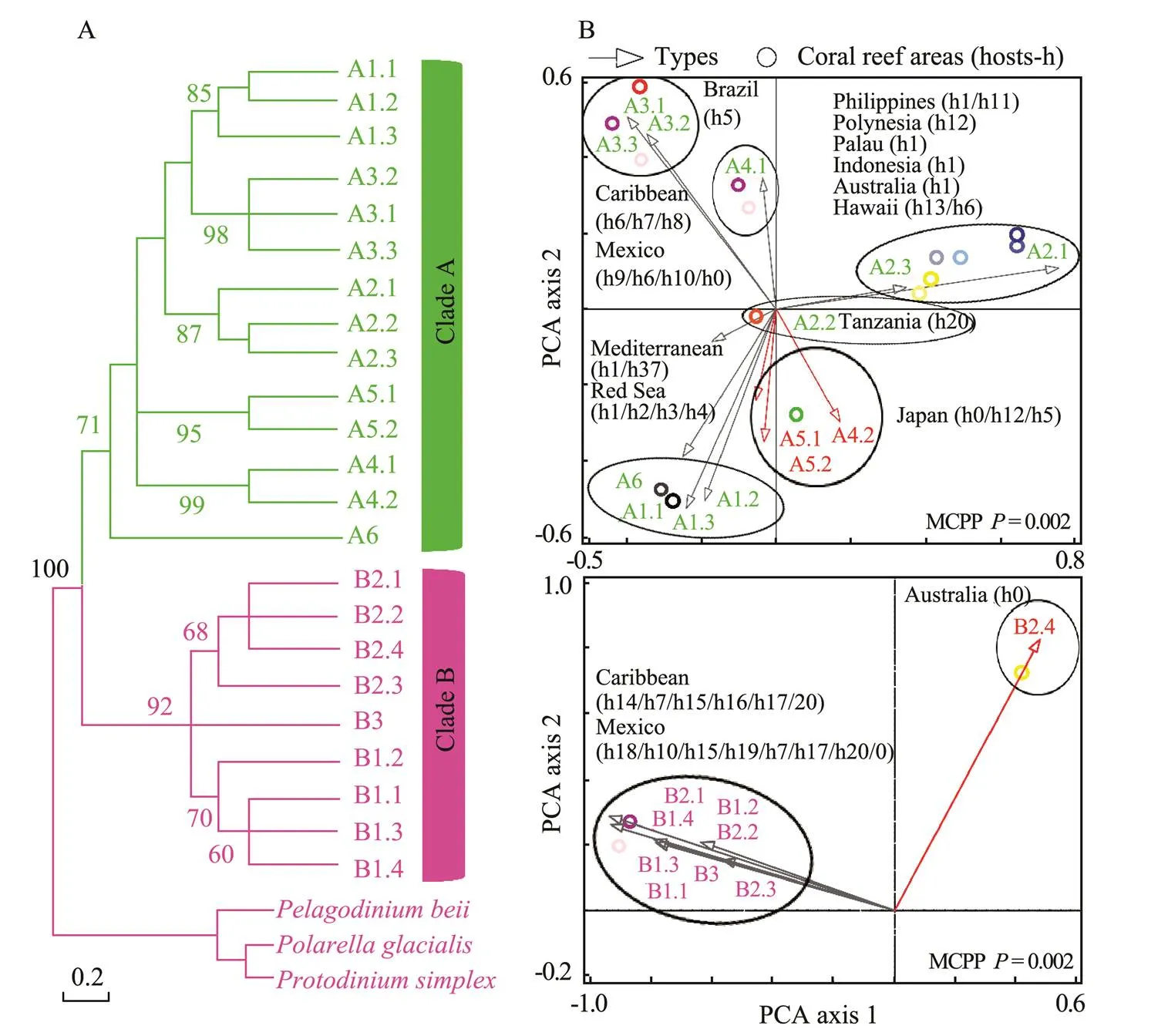
Fig.4 Phylogenetic tree (A) and principal component analysis (B) of Symbiodiniaceae ITS2 types in clades A and B. A phylogenetic tree (maximum likelihood) was constructed using Symbiodiniaceae ITS2 representative sequences in MEGA 6.06. Symbiodiniaceaewas subjected to principal component analysis based on the occurrence records of Symbiodiniaceae in different coral reef areas (empty arrows indicate the hosts associated with Symbiodiniaceae ITS2 types, red arrows refer to free-living Symbiodiniaceae ITS2 types, and circles represent coral reef areas). The host information (h) is presented within parentheses next to the name of each coral reef area. h0, free-living Symbiodiniaceae; h1, Tridacna spp.; h2, Stylophora spp.; h3, Pocillopora spp.; h4, Nephthea spp.; h5, Zoanthus spp.; h6, Porites spp.; h7, Plexaura spp.; h8, Cliona spp.; h9, Gorgonia spp.; h10, Orbicella spp.; h11, Hippopus spp.; h12, Acropora spp.; h13, Cassiopeia spp.; h14, Madracis spp.; h15, Eunicea spp.; h16, Plexaurella spp.; h17, Antillogorgiaspp.; h18, Montastraea spp.; h19, Actinia spp.; h20, Millepora spp. MCPP shows the P-value of the Monte Carlo permutation test.
Further rarefaction curve analysis of Symbiodiniaceae ITS2 sequences and observed Symbiodiniaceae ITS2 type in coral reef areas in the Atlantic Ocean, the Mediterranean Sea, the Rea Sea, the Indo-Pacific Ocean, and Australian waters show that the number ofSymbiodiniaceae ITS2 sequences from the four reef areas exceeded the point where the rarefaction curve reached its asymptote.
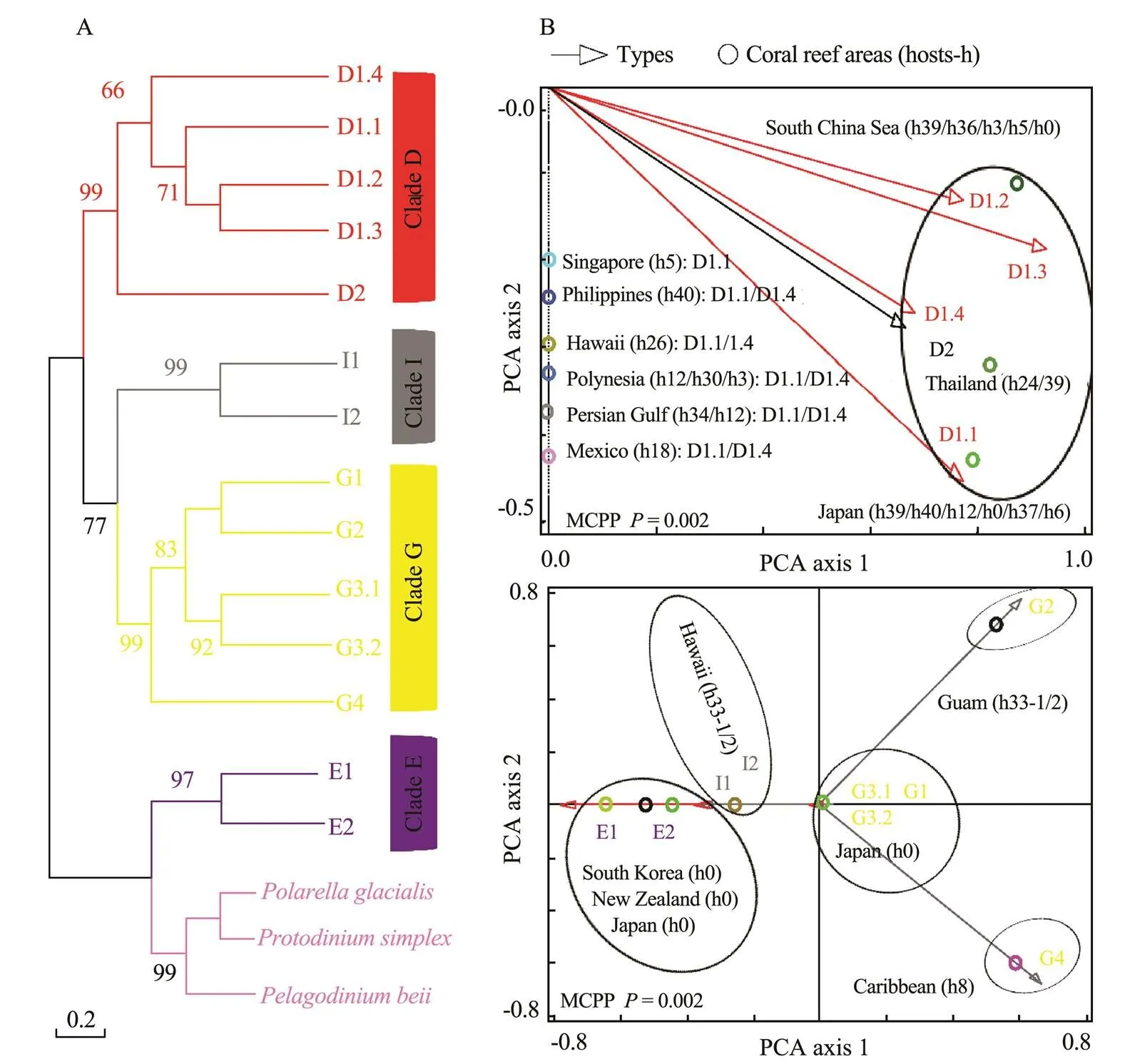
Fig.5 Same as Fig.4 but for clades D, E, G, and I. The host information (h) is presented within parentheses next to the name of each coral reef area. h0, free-living Symbiodiniaceae; h3, Pocillopora spp.; h5, Zoanthus spp.; h6, Porites spp.; h8, sponges (Cliona spp.); h12, Acropora spp.; h18, Montastraea spp.; h24, Goniastrea spp.; h26, Montipora spp.; h30, Leptoseris spp.; h33, foraminifera (h33-1, Sorites spp.; h33-2, Marginopora spp.); h34, Palythoa spp.; h36, Galaxea spp.; h37, anemone (unknown); h39, Oulastrea spp.; and h40, Pteraeolidia spp. MCPP shows the P-value of the Monte Carlo permutation test.

Fig.6 Same as Fig.4 but for clades F and H. The host information (h) is presented within parentheses next to the name of the coral reef area. h18, Montastraea spp.; h26, Montipora spp.; h33, foraminifera (Sorites spp.); h41, Meandrina spp.; and h42, Sinularia spp. MCPP shows the P-value of the Monte Carlo permutation test.
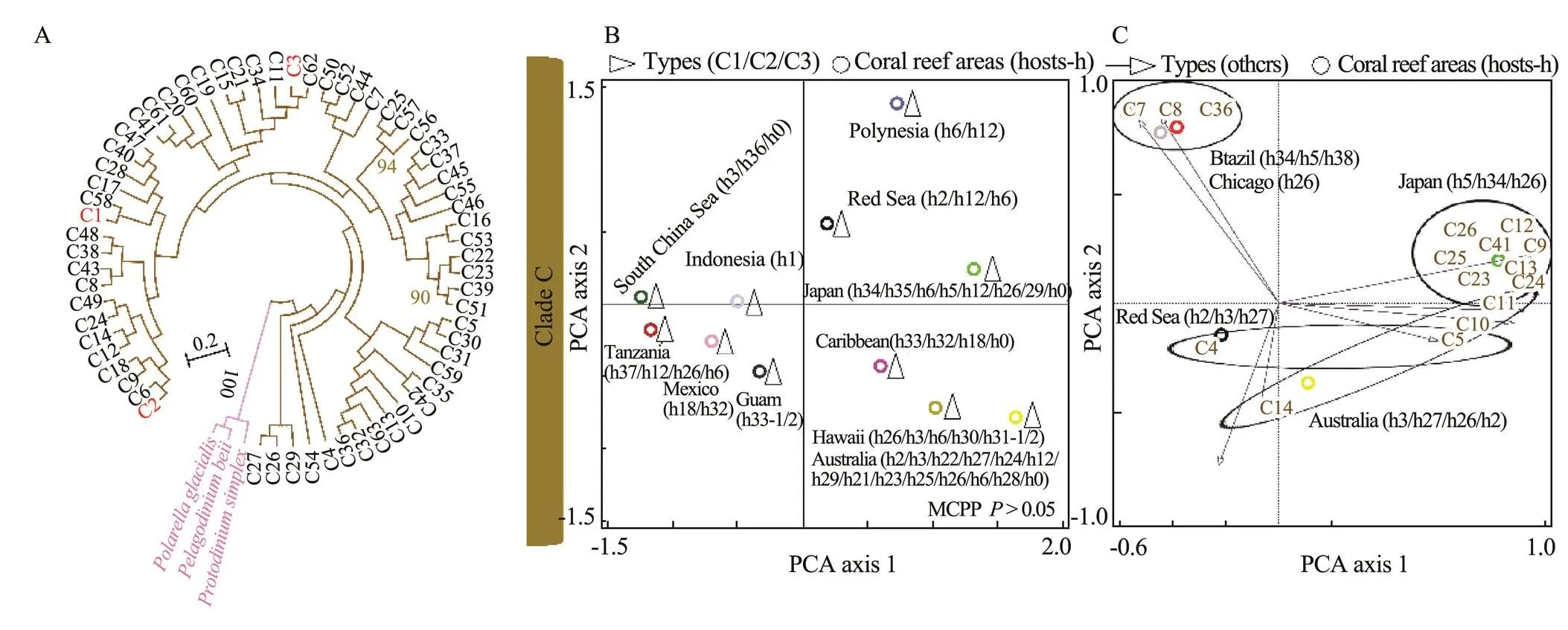
Fig.7 Phylogenetic tree (A) and principal component analysis (B) of Symbiodiniaceae ITS2 types C1, C2, and C3 (B) and other Symbiodiniaceae ITS2 types (C) in clade C. A phylogenetic tree (maximum likelihood) was constructed using Symbiodiniaceae ITS2 representative sequences in MEGA 6.06. Symbiodiniaceaewas subjected to principal component analysis based on the occurrence records of Symbiodiniaceae in different coral reef areas (empty triangles (B) indicate Symbiodiniaceae ITS2 types C1, C2, and C3; empty arrows (C) refer to the hosts associated with Symbiodiniaceae ITS2 types; and circles represent coral reef areas). The host information (h) is presented within parentheses next to the name of the coral reef area. h0, free-living Symbiodiniaceae; h1, Tridacna spp.; h2, Stylophora spp.; h3, Pocillopora spp.; h5, Zoanthus spp.; h6, Porites spp.; h12, Acropora spp.; h18, Montastraea spp.; h21, Coscinaraea spp.; h22, Entacmaea spp.; h23, Favites spp.; h24, Goniastrea spp.; h25, Marginopora spp.; h26, Montipora spp.; h27, Seriatopora spp.; h28, Symphyllia spp.; h29, Turbinaria spp.; h30, Leptoseris spp.; h31, black corals (h31-1, Antipathes spp.; h31-2, Aphanipathes spp.); h32, Agaricia spp.; h33, foraminifera (h33-1, Sorites spp.; h33-2, Marginopora spp.); h34, Palythoa spp.; h35, Plesiastrea spp.; h36, Galaxea spp.; h37, Condylactis spp.; and h38, Protopalythoa spp. MCPP shows the P-value of the Monte Carlo permutation test.
These results revealed the geographic segregation ofSym-biodiniaceae clades, especially Symbiodiniaceae ITS2 types, in the Atlantic Ocean, the Mediterranean Sea, the Red Sea, and the Indo-Pacific Ocean reef areas.
In summary, this study provided a broad overview of the diversity and distribution of Symbiodiniaceae clades/ typeson a global scale.
4 Discussion
Recognizing the diversity of Symbiodiniaceae is chal-lenging because neither sex differences nor significant morphological differences are readily observable among Symbiodiniaceae types (Santos and Coffroth, 2003). With the practical difficulties associated with analyzing the diversity of Symbiodiniaceae according to biological and morphological features, the phylogenetic species concept has been used to understand the diversity of Symbiodiniaceae (Correa and Baker, 2009), whose members are hy- pothesized to differentiate into potentially ecologically dis- tinct entities (Correa and Baker, 2009). In accordance with the phylogenetic species concept, nine phylogenetically supported Symbiodiniaceae clades, known as clades A–I, are classified on the basis of largely conserved gene markers(Pochon., 2014). Symbiodiniaceae clades are further subdivided into subclade types or putative ‘species’ by using less conserved gene markers (Lajeunesse and Thornhill, 2011). The less conserved gene ITS2 marker has been the most widely used to survey the diversity of Symbiodiniaceae in reef-associated metazoans and foraminifera and in environmental samples near coral reefs in different regions (Arif., 2014).
This study showed that 119 Symbiodiniaceae ITS2 types within nine clades (A–I) mainly inhabited coral reefs be- tween 30˚N and 30˚S. Franklin(2012) conducted a similar study, but they reported 409 Symbiodiniaceae sub- clade types. This difference between Franklin’s study and our study might be caused by the different methods used to collect and process the ITS2 sequences. Raw Symbiodiniaceae ITS2 sequences were collected by Franklin(2012) to represent types. In the present study, the qualified Symbiodiniaceae ITS2 sequences were clustered and annotated as Symbiodiniaceae ITS2types at a 97% simila- rity cutoff value (Arif.,2014). According to Arif(2014), ITS2 sequence similarities with >97% are consi- dered indicative of the same Symbiodiniaceae type because they likely represent nonspecific amplifications of closely related types or intragenomic sequence divergences. We acknowledged that some Symbiodiniaceae types could not be differentiated with a cutoff value of >97% similarity. For instance, the clade C, which includes many types, may require higher cutoff values to differentiate the diversity of Symbiodiniaceae at the type level (Arif., 2014). A minimum entropy decomposition- based approach (referred to as the metahaplotype approach) has been used to consolidate the high diversity to a smaller setof core sequence nodes based on biologically informative sequence positions rather than the fixed similarity thresholds of the OTU approach (Eren., 2015; Smith., 2017). Therefore, our results provided deflated ra- ther than inflated Symbiodiniaceae diversity estimates.
Symbiodiniaceae clades A and F were distributed glo- bally (Fig.3), whereas the Symbiodiniaceae ITS2 types with- in the two clades were geographically segregated (Figs.4 and 6). Even for the globally distributed Symbiodiniaceae clade C, the geographic segregation of certain Symbiodiniaceae ITS2 types within this clade was also suggested (Fig.7C). For example, Symbiodiniaceae ITS2 types C7, C8, and C36 were mainly associated with hosts living in coral reef areas near Brazil and a host from an aquarium in Chicago. Symbiodiniaceae ITS2 types C4 and C14 were mainly related to hosts in the Red Sea. Symbiodiniaceae ITS2 types C5, C9, C10, C11, C12, C13, C23, C24, C25, C26, and C41 were mainly linked to hosts living in the coral reef areas in the Indo-Pacific Ocean (near Japan). Our study, some previous studies (Lajeunesse, 2002, 2003; Lajeunesse., 2005; van Oppen., 2009), and one review (Baker, 2003) revealed that the Symbiodiniaceae ITS2types within clade B were mainly distributed in the Atlantic Ocean. This result implied that they are endemic Symbiodiniaceae ITS2typesLajeunesse. (2005) also suggested that the distinct geographic distributions of Symbiodiniaceae are likely related to the hypothesis that the closure of the Central American Seaway by the uplift of the Central American Isthmus occurred 3.1–3.5 millionyears ago. Consequently, a barrier formed between the Atlantic Ocean and the Indo-Pacific Ocean and likely fa- vored the growth of distinct communities of Symbiodiniaceae in the two oceans. The distinctness of the Symbiodiniaceae community in the Red Sea and the Mediterranean Sea may be due to specific physical geographic features in the region (creating landlocked marine ecosystems) and regional environmental conditions,., ‘higher sea surface temperatures (SST) and salinity than those in other ocean regions’ (Sawall., 2014).
The flexible associations of Symbiodiniaceae with different hosts were observed in this study. For example, the following Symbiodiniaceae types were associated with different hosts at the phylum or genus level: A3.1, A5.1, A5.2, A4.2, and B1.1 (Fig.4); D1.1, 1.2, D1.3, E1, E2, E3, G1, and G3.1 (Fig.5); and C1, C2, and C3 (Fig.7). Speci- fically, this study revealed foraminifera-hosted Symbiodiniaceae in clades C, F, G, H, and I, whereas Baker (2003) reported foraminifera-hosted Symbiodiniaceae in clades C, F, and G. In addition, as the main component of the coral reef ecosystem, sponges could be associated with multiple Symbiodiniaceae clades; for example, they were related to Symbiodiniaceae ITS2 types belonging to clades A and G. This finding might have been observed because the survival of sponges depends on food particles from seawater (Buddemeier and Fautin, 1993) and the nutrition acquired from Symbiodiniaceae.
Corals and their supported coral reefs have suffered unprecedented declines in species abundance and community degradation over the last few decades because of the effects of climate change and anthropogenic disturbances, and further declines and degradation are projected in the near future (Hughes., 2017). An emerging ‘nugget of hope’ for these threats to corals and their supported coral reefs is related to the interactions of corals with the ecological, physiological, and functional diversity of Symbiodiniaceae clades/types. For example, extremely high temperatures can lead to coral bleaching (a result of the breakdown of the interactions of coral hosts with Symbiodiniaceae); however, corals with more heat- tolerant Symbiodiniaceae (., heat-tolerant Symbiodiniaceae in clade D) less likely bleach than other corals, and bleached corals gain Symbiodiniaceae with a high heat tolerance after recovery (van Oppen., 2009). Our re- sults showed that corals are the most flexible hosts mainly associated with Symbiodiniaceae ITS2 types within clades A, B, C, D, and F. According to the ‘adaptive bleaching hypothesis’ proposed by Buddemeier. (1993), such flexible associations between Symbiodiniaceae and coral hosts may serve as an adaptive mechanism of corals under the effects of marine environmental changes (Putnam., 2012). Our study, previous studies, and reviews revealed that the Symbiodiniaceae belonging to clade B mainly existed in the Atlantic Ocean (Baker, 2003; Lajeunesse, 2003), implying that its members might be important for the local acclimation of coral–Symbiodiniaceae associations. A previous study suggested that Symbiodiniaceae clade D is typically observed as weedy or opportunistic algae that preferentially occupy recently stressed or marginal habitats, inshore regions, and bleached corals (Stat., 2013). These results showed that the Symbiodiniaceae belonging to clade D mainly occur in the Indo-Pacific Ocean (specifically, in corals living near Thailand, the South China Sea, and Japan). Therefore, corals living in the Indo-Pacific Ocean were likely threatened, but coral species mainly associated with clade D Symbiodiniaceae might have a more positive status, as previous studies suggested that corals composed of Symbiodiniaceae with a high heat tolerance (., heat-tolerant Symbiodiniaceae in clade D) were less likely to bleach.
The term ‘free-living Symbiodiniaceae’ is defined as fol- lows: A) Symbiodiniaceae living outside a host while re- taining the ability to form symbioses with hosts (Hirose., 2008), including ‘transiently free-living’ Symbiodiniaceae that has been recently expelled from their hosts and may not persist in a free-living state (Yamashita., 2013) or B) novel Symbiodiniaceae types not currently known to engage in symbioses permanently in a free-living state (Takabayashi., 2012). According to adaptive bleaching hypothesis, free-living Symbiodiniaceae may provide an open pool from which hosts under environmental stress can obtain symbionts (Johnson, 2011). In addition, free-living Symbiodiniaceae can affect the com- munity composition ofSymbiodiniaceae associated with different hosts (Manning and Gates, 2008; Takabayashi., 2012; Cunning., 2015) because Symbiodiniaceae may be routinely expelled by reef-associated metazoans,., jellyfish and reef-building corals (Kinzie., 2001; Baird., 2009; van Oppen., 2009). This family may also be exogenously acquired by juvenile and adult coralshorizontal transfer (Stat., 2008; Baird., 2009). Our study partially supported these ideas as most Symbiodiniaceae ITS2 types belonging to clades A, B, C, and D were observed in various reef-associated hosts and seawater or sediments near coral reef areas. Furthermore, some Symbiodiniaceae ITS2 types (., most Symbiodi- niaceae ITS2 types within clades E and G) mainly existed in a free-living state.
5 Conclusions
The diversity and geographic distribution of Symbiodi- niaceae were analyzed on the basis of the Symbiodiniaceae ITS2 sequences derived from metazoans and foraminifera associated with reefs and from seawater and sediments near coral reef areas in the GenBank database.In particular, the possible geographic segregation of Sym- biodiniaceae ITS2 types implied that different Symbiodi- niaceae ITS2 types might have different ecological functions. For example, the Symbiodiniaceae ITS2 types belonging to clade B were mainly from the Atlantic Ocean, implying that they were important for the local acclimation of host–Symbiodiniaceae associations. By contrast, the Symbiodiniaceae ITS2 types belonging to clade D were mainly found in the Indo-Pacific Ocean, suggesting that they might be essential for the maintenance of host–Sym- biodiniaceae associations under environmental stress. How- ever, the 3899 qualified Symbiodiniaceae sequences from GenBank were obtained from reef-associated metazoans and foraminifera and from seawater and sediment samples near coral reef areas by different researchers using various methods at different periods; consequently, sampling or method biases were unavoidable. Thus, any conclusions were premature at this stage. Nevertheless, our re-sults provided a broad overview of Symbiodiniaceae clade/ type diversity and distributionon a global scale. An international program of the global sampling and investigation of Symbiodiniaceaea consistent research stra- tegy should be developed to fully understand the diversity and distribution of Symbiodiniaceae clades/types on a glo- bal scale.
1) Multiple gene barcodes (., ITS2, ITS, ITS1, rbcL, and matK) should be assessed and used to survey Symbiodiniaceae typediversity, and 28S rDNA should be uti- lized as the benchmark barcode of different Symbiodiniaceae clades (Pochon., 2012, 2014).
2) High-resolution methods, including quantitative PCR and high-throughput DNA sequencing, should be considered, as recent studies revealed that rare Symbiodiniaceae clades/types are common in different hosts and environmental samples, which cannot be detected with traditional methods (., DGGE or RFLP; Arif., 2014).
3) Known and potential hosts associated with Symbiodiniaceae and seawater, sand, and sediments near reef areas should be uniformly sampled from different coral reefs areas.
4) Environmental factors (., temperature, pH, light, inorganic and organic pollutants, and sampling depth) in different coral reefs areas should be considered.
5) A standardized nomenclature system of Symbiodi- niaceae types and a related database should be constructed on the basis of the above aspects to monitor the dynamics of Symbiodiniaceae on a global scale and determine the effects of environmental factors on coral reefs.
Acknowledgements
This work was funded by the National Key Basic Research Program of China (No. 2013CB956103), the Key Ecological Processes and Health Regulation Principles of Marine Ecosystem in Guangdong-Hong Kong-Macao Grea-ter Bay Area (No. GML2019ZD0405), the CAS Pioneer Hundred Talents Program, and the South China Sea Insti- tute of Oceanography, CAS (Nos. Y8SL031001 and Y9YB021001).
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J., 1990. Basic local alignment search tool., 215: 403-410.
Arif, C., Daniels, C., Bayer, T., Hinestroza, E. B., Barbrook, A., Howe, C. J., Lajeunesse, T. C., and Voolstra, C. R., 2014. Assessingdiversity in scleractinian coralsnext- generation sequencing-based genotyping of the ITS2 rDNA region., 23: 4418-4433.
Baird, A. H., Guest, J. R., and Willis, B. L., 2009. Systematic and biogeographical patterns in the reproductive biology of scleractinian corals., 40: 551-571.
Baker, A. C., 2003. Flexibility and specificity in coral-algal sym- biosis: Diversity, ecology, and biogeography of., 34: 661-689.
Blackall, L. L., Wilson, B., and van Oppen, M. J., 2015. Coral–the world’s most diverse symbiotic ecosystem., 24: 5330-5347.
Buddemeier, R. W., and Fautin, D. G., 1993. Coral bleaching as an adaptive mechanism: A testable hypothesis., 43: 320-326.
Coffroth, M. A., and Santos, S. R., 2005. Genetic diversity of symbiotic dinoflagellates in the genus., 156: 19-34.
Coffroth, M. A., Lewis, C. F., Santos, S. R., and Weaver, J. L., 2006. Environmental populations of symbiotic dinoflagellates in the genuscan initiate symbioses with reef cnidarians., 16: 985-987.
Correa, A. M. S., and Baker, A. C., 2009. Understanding diversity in coral-algal symbiosis: A cluster-based approach to interpreting fine-scale genetic variation in the genus., 28: 81-93.
Cunning, R., Gates, R. D., and Edmunds, P. J., 2017. Using high- throughput sequencing of ITS2 to describemetacommunities in St. John, U.S. Virgin Islands., 5: e3472.
Cunning, R., Yost, D. M., Guarinello, M. L., Putnam, H. M., and Gates, R. D., 2015. Variability ofcommunities in waters, sediments, and corals of thermally distinct reef pools in American Samoa., 10: e0145099.
De’Ath, G., Fabricius, K. E., Sweatman, H., and Puotinen, M., 2012. The 27-year decline of coral cover on the Great Barrier Reef and its causes., 109: 17995-17999.
Eren, A. M., Morrison, H. G., Lescault, P. J., Reveillaud, J., Vineis, J. H., and Sogin, M. L., 2015. Minimum entropy decomposition: Unsupervised oligotyping for sensitive partitioning of high-throughput marker gene sequences., 9: 968.
Fautin, D. G., and Buddemeier, R. W., 2004. Adaptive bleaching: A general phenomenon., 530: 459-467.
Franklin, E. C., Stat, M., Pochon, X., Putnam, H. M., and Gates, R., 2012. GeoSymbio: A hybrid, cloud-based web application of global geospatial bioinformatics and ecoinformatics for–Host symbioses.,12: 369-373.
Freudenthal, H. D., 1962.gen. nov. andspp. nov., a Zooxanthella: Taxonomy, life cycle, and morphology., 9: 45-52.
Green, E. A., Davies, S. W., Matz, M. V., and Medina, M., 2014. Quantifying crypticdiversity withinandat the Flower Garden Banks, Gulf of Mexico., 2: e386.
Hirose, M., Reimer, J. D., Hidaka, M., and Suda, S., 2008. Phylogenetic analyses of potentially free-livingspp. isolated from coral reef sand in Okinawa, Japan., 155: 105-112.
Hughes, T. P., Kerry, J. T., Lvarez-Noriega, M., Lvarez-Romero, J. G., and Wilson, S. K., 2017. Global warming and recurrent mass bleaching of corals., 543: 373-377.
Johnson, M. D., 2011. The acquisition of phototrophy: Adaptive strategies of hosting endosymbionts and organelles., 107: 117-132.
Jones, R. J., 2008. Coral bleaching, bleaching-induced mortality, and the adaptive significance of the bleaching response., 154: 65-80.
Keshavmurthy, S., Meng, P. J., Wang, J. T., Kuo, C. Y., Yang, S. Y., and Hsu, C. M., 2014. Can resistant coral–associations enable coral communities to survive climate change? A study of a site exposed to long-term hot water input., 2: e327.
Kinzie, R. A., Takayama, M., Santos, S. R., and Coffroth, M. A., 2001. The adaptive bleaching hypothesis: Experimental tests of critical assumptions., 200: 51-58.
Lajeunesse, T. C., 2002. Diversity and community structure of symbiotic dinoflagellates from Caribbean coral reefs., 141: 387-400.
Lajeunesse, T. C., 2005. ‘Species’ radiations of symbiotic dino- flagellates in the Atlantic and Indo-Pacific since the Miocene–Pliocene transition., 22: 570- 581.
Lajeunesse, T. C., and Thornhill, D. J., 2011. Improved resolution of reef-coral endosymbiont () species diversity, ecology, and evolution through psbA non-coding region genotyping., 6: e29013.
Lajeunesse, T. C., Loh, W., van Woesik, R., Hoegh-Guldberg, O., Schmidt, G. W., and Fitt, W. K., 2003. Low symbiont diversity in southern Great Barrier Reef corals, relative to those of the Caribbean., 48: 2046- 2054.
Lajeunesse, T. C., Parkinson, J. E., Gabrielson, P. W., Jeong, H. J., Reimer, J. D., Voolstra, C. R., and Santos, S. R., 2018. Sys- tematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts., 28: 1- 11.
Manning, M. M., and Gates, R. D., 2008. Diversity in populations of free-livingfrom a Caribbean and Pacific reef., 53: 1853-1861.
Pochon, X., Putnam, H. M., and Gates, R. D., 2014. Multi-gene analysis ofdinoflagellates: A perspective on rarity, symbiosis, and evolution., 2: e394.
Pochon, X., Putnam, H. M., Burki, F., and Gates, R. D., 2012. Identifying and characterizing alternative molecular markers for the symbiotic and free-living dinoflagellate genus., 7: e29816.
Prada, C., McIlroy, S. E., Beltrán, D. M., Valint, D. J., Ford, S. A., Hellberg, M. E., and Coffroth, M. A., 2014. Cryptic diversity hides host and habitat specialization in a gorgonian-algal symbiosis., 23: 3330-3340.
Putnam, H. M., Stat, M., Pochon, X., and Gates, R. D., 2012. Endosymbiotic flexibility associates with environmental sensitivity in scleractinian corals.–, 279: 4352-4361.
Quigley, K. M., Davies, S. W., Kenkel, C. D., Willis, B. L., Matz, M. V., and Bay, L. K., 2014. Deep-sequencing method for quantifying background abundances oftypes: Exploring the rarebiosphere in reef-building corals., 9: e94297.
Reimer, J. D., Herrera, M., Gatins, R., Roberts, M. B., Parkinson, J. E., and Berumen, M. L., 2017. Latitudinal variation in the symbiotic dinoflagellateof the common reef zoantharianon the Saudi Arabian coast of the Red Sea., 44: 661-673.
Santos, S. R., and Coffroth, M. A., 2003. Molecular genetic evidence that dinoflagellates belonging to the genusare haploid., 204: 10-20.
Sawall, Y., Al-Sofyani, A., Banguera-Hinestroza, E., and Voolstra, C. R., 2014. Spatio-temporal analyses ofphysiology of the coralalong large- scale nutrient and temperature gradients in the Red Sea., 9: e103179.
Smith, E. G., Ketchum, R. N., and Burt, J. A., 2017. Host specificity ofvariants revealed by an ITS2 metahaplotype approach., 11: 1500-1503.
Stat, M., Loh, W. K. W., Hoegh-Guldberg, O., and Carter, D. A., 2008. Symbiont acquisition strategy drives host-symbiont associations in the southern Great Barrier Reef., 27: 763-772.
Stat, M., Pochon, X., Franklin, E. C., Bruno, J. F., Casey, K. S., and Selig, E. R., 2013. The distribution of the thermally tolerant symbiont lineage (clade D) in corals from Hawaii: Correlations with host and the history of ocean thermal stress., 3: 1317-1329.
Takabayashi, M., Adams, L. M., Pochon, X., and Gates, R. D., 2012. Genetic diversity of free-livingin surface water and sediment of Hawai’i and Florida., 31: 157-167.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S., 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0., 30: 2725-2729.
Ter Braak, C. J. F., and Smilauer, P., 2002. CANOCO reference manual and CanoDraw for Windows user’s guide: Software for canonical community ordination (version 4.5). http://www. canoco.com.
Thomas, L., Kendrick, G. A., Kennington, W. J., Richards, Z. T., and Stat, M., 2014. Exploringdiversity and host specificity incorals from geographical extremes of western Australia with 454 amplicon pyrosequencing., 23: 3113-3126.
Tonk, L., Sampayo, E. M., Weeks, S., Magno-Canto, M., and Hoegh-Guldberg, O., 2013. Host-specific interactions with en- vironmental factors shape the distribution ofacross the Great Barrier Reef., 8: e68533
van Oppen, M. J. H., Baker, A. C., Coffroth, M. A., and Willis, B. L., 2009. Bleaching resistance and the role of algal endosymbionts. In:. van Oppen, M. J. H., and Lough, J. M., eds., Springer, Berlin, Heidelberg, 83-102.
Yamashita, H., and Koike, K., 2013. Genetic identity of free- livingobtained over a broad latitudinal range in the Japanese coast., 61: 68-80.
Ziegler, M., Arif, C., Burt, J. A., Dobretsov, S., Roder, C., and Lajeunesse, T. C., 2017. Biogeography and molecular diversity of coral symbionts in the genusaround the Arabian Peninsula., 44: 674-686.
October 22, 2019;
July 6, 2020;
September 8, 2020
© Ocean University of China, Science Press and Springer-Verlag GmbH Germany 2021
. Tel: 0086-21-34204036
E-mail: zyli@sjtu.edu.cn
(Edited by Chen Wenwen)
杂志排行

Journal of Ocean University of China的其它文章
- Elevated Ducts and Low Clouds over the Central Western Pacific Ocean in Winter Based on GPS Soundings and Satellite Observation
- Control System Design and Implementation at Flexible, Distributed Offshore Sensor Test Sites in the Yangtze Estuary Area
- A Preliminary Investigation of Arctic Sea Ice Negative Freeboard from in-situ Observations and Radar Altimetry
- Comparison of the Pore Structure of Ultralow-Permeability Reservoirs Between the East and West Subsags of the Lishui Sag Using Constant-Rate Mercury Injection
- The Stable Carbon Isotopic Compositions of n-Alkanes in Sediments of the Bohai and North Yellow Seas: Implications for Sources of Sedimentary Organic Matter
- Composition and Distributions of Nitrogen and Phosphorus and Assessment of Eutrophication Status in the Maowei Sea