雅安周公山不同发育阶段峨眉含笑的枝叶性状
2021-03-04赵安玖陈智超吕佳瑶李曜汐
彭 希,赵安玖,陈智超,吕佳瑶,李曜汐
(四川农业大学 林学院,四川 成都 611130)
植物功能性状是指对植物个体生存与发展有着一定影响的植物特征,与植物自身的生存策略密切相关[1−2]。植物的生物学特性间接反映植物各功能性状间的权衡方式,影响着植物在群落中的生存与发展。在植物的各种器官间,小枝和叶片是植物器官分支系统中最敏感的部分,在植物的不同发育阶段,小枝和叶性状是植物与环境相互作用的结果[3]。植物如何通过调整小枝内各构件之间的生长关系,来适应不断变化的环境,是研究植物生态策略的重要内容[4]。作为植物光合作用重要的器官,叶片能够将光能转化为自身发育所需要的物质,其大小直接影响着植物个体的发育模式以及对光能的捕获和转化能力[5]。小枝是植物叶片直接着生的器官,它能够传输茎运输过来的养分及叶光合作用产生的同化物质,与植物在空间上的开展情况密切相关,并决定着叶片的投资方式[6]。小枝与叶片之间的关系是植物个体在不同发育阶段的生态策略体现[7]。不同物种对环境的适应情况不同,因此生存策略方式也不同,即快速高效策略和慢速高质量策略[8−10]。物种内存在不同发育阶段的植物个体,它们在环境中获得的资源情况不同。为了提高对环境的适应性,植物枝叶可能会在不同发育阶段有着不同的投资方式。植物小枝与叶片对环境变化的敏感性强,植物个体在不同发育阶段的生态策略容易在其性状上表现出来[10]。种群空间格局指种群个体在空间上的分布情况,是与外界环境相互作用的结果,在一定程度上反映种群的生态策略方式[11]。目前,关于种群空间格局的研究主要集中在不同物种间、物种内不同发育阶段等方面[11−12],在植物性状间空间格局的分析较少。地统计学作为研究植物空间格局的重要方法,可以真实反映植物的空间变化[13]。SIEFERT[14]采用半方差变异函数对农田植物叶功能性状的空间格局进行了研究,发现叶功能性状具有中等空间自相关性,植物功能性状在空间上是连续变化的,进而能够预测植物性状的空间变化情况。探究植物枝叶性状的空间变化有助于理解植物对环境的适应策略。峨眉含笑Michelia wilsonii是多年生常绿乔木,主要生长于气候湿润,海拔600~2 000 m的常绿阔叶林中。虽然峨眉含笑的结实量较多,但其自然更新困难,分布区域较小,现已濒临灭绝,为国家Ⅱ级保护树种。目前,对于峨眉含笑的研究主要集中在群落结构、凋落物特征等方面[15−16],对其生态策略方式还不清楚。因此,本研究以四川省雅安周公山峨眉含笑优势阔叶林为基础,选取种群内不同发育阶段个体为研究对象,分析枝叶性状在空间上的自相关性,以期为了解峨眉含笑枝叶性状之间的资源权衡方式和生态策略方式提供依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于四川省雅安周公山国家森林公园内 (29°58′09″N,103°02′58″E)。该区属亚热带季风性湿润气候,降水量大,年均降水量为1 774.3 mm;湿度大,年平均相对湿度79%;气温较低,年平均气温14.0 ℃,海拔900~1 180 m;土壤类型以山地黄壤为主。研究区内植被丰富,以常绿阔叶林为主,除峨眉含笑优势树种外,伴生有杉木Cunninghamia lanceolata、华中樱桃Cerasus conradinae、栗Castanea mollissima、日本杜英Elaeocarpus japonicus、刺楸Kalopanax septemlobus、灯台Bothrocaryum controversum、南酸枣Choerospondias axillaris等乔木树种,林分郁闭度为0.8~0.9;林下主要有水竹Phyllostachys heteroclada、姬蕨Hypolepis punctata及一些禾草等植物,盖度达80%以上[15]。
1.2 样地设置
在峨眉含笑种群分布较多的地区,建立1块200 m×200 m的固定样地,并将样地划分为100个20 m×20 m的样方。逐一对样方内所有胸径≥5 cm的峨眉含笑个体进行挂牌登记,并记录胸径、树高等信息,以便进行后期采样工作。
1.3 枝叶采集
于2018年7月采集样地内峨眉含笑个体的叶片和枝条。根据研究区域峨眉含笑种群的年龄结构及数量动态特征[16],将其划分为小树(胸径5~15 cm)、中树(胸径15~25 cm)和大树(胸径>25 cm) 3个阶段。定义当年生小枝为1年生小枝,沿着1年生小枝依次向内为2年生、3年生小枝。为了保证每株树采集的1、2年生枝条和叶来自同一个树枝,本研究在树冠南部枝条分叉处直径约1 cm粗的地方剪取1个树枝,从树枝上采集6个能明显区分1、2年生的小枝。采集小枝上成熟的当年生叶片15片,并将1、2年生小枝中均匀生长的部分截断,将其保存在自封袋中,做好标记带回实验室进行后续指标测定。共采集20株小树、59株中树、34株大树的枝叶。
1.4 指标测定
叶片鲜质量使用电子天平(精度0.000 1 g)称量;将称量过后的叶片放入烘箱中,在105 ℃下烘20 min进行杀青,然后在80 ℃下烘48 h,用电子天平称其干质量(精度0.000 1 g)。用叶面积(LI-3100C,LI COR,美国)扫描仪测定叶片后,在PS6中计算叶片面积;用游标卡尺测量叶片厚度(精度0.01 mm)。叶干鲜比=叶片干质量(g)/叶片鲜质量(g);比叶面积=叶片面积(cm2)/叶片干质量(g)。将所有小枝浸入去离子水中8~12 h,待小枝达到饱和状态后将其取出,用排水法测量小枝体积。将测量完体积的小枝,用吸水纸吸去表面水分后用电子天平称量(精度0.000 1 g),获得小枝鲜质量,然后将其放入烘箱中80 ℃烘48 h,用电子天平称量(精度0.000 1 g),获得小枝干质量。小枝干鲜比=小枝干质量(g)/小枝鲜质量(g);小枝密度=小枝干质量(g)/小枝体积(cm3)。
1.5 数据处理
采用单因素方差分析对峨眉含笑种群不同发育阶段枝叶性状进行差异分析;采用标准化主轴分析计算枝叶性状间的相关性。为了使枝叶性状呈正态分布,对其进行了对数转换(以10为底数)。数据分析在R语言的smatR包中进行。
通过主成分分析获得每株树木第1轴枝叶性状的得分值,然后采用半方差函数法对峨眉含笑种群不同发育阶段枝叶性状主成分得分进行空间格局分析[17−18]。其计算公式为:
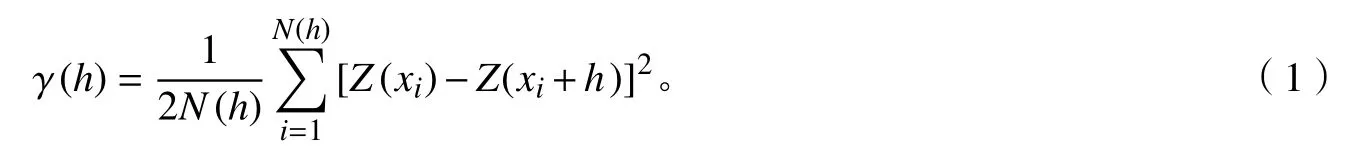
式(1)中:γ(h)为林木个体间距为h时的半方差函数值;N(h)为林木个体间距为h时的样本对数;Z(xi)为林木个体i在位置点xi处的实测值;Z(xi+h)为林木个体在位置点xi+h处的实测值。通过半方差值进行模型拟合,绘制半方差函数图。
本研究选取4种模型进行拟合(球状模型、指数模型、高斯模型和线性模型),选取模型决定系数(R2)最大和残差最小的模型。通过最优模型获得基台值(C0+C)、偏基台值(C)、块金值(C0)和变程(A),求得结构方差比[C/(C0+C)]来表示研究区变量的最大变异程度。一般认为结构方差>75%时,空间自相关性强;结构方差位于25%~75%时,空间自相关性中等,结构方差<25%时,空间自相关弱,此时不宜采用克里格(Kringing)插值进行变量预测。
2 结果与分析
2.1 峨眉含笑不同发育阶段枝叶性状特征
不同发育阶段叶鲜质量、叶干质量和2年生小枝密度差异显著(P<0.05),大树与中树、小树之间叶面积、比叶面积、1年生小枝密度差异显著(P<0.05),中树和小树之间叶干鲜比和2年生小枝干鲜比差异显著(P<0.05),其余性状差异不显著(P>0.05)(表1)。叶鲜质量、叶干质量、叶面积、叶厚、1年生小枝干鲜比、1年生小枝密度随着林木胸径的增加呈显著增加(P<0.05),比叶面积随着林木胸径的增加显著降低(P<0.05)(表2)。主成分分析前2轴的解释总方差为56.99%,其中第1轴的解释方差为39.95%,第2轴的解释方差为17.04%(图1)。对枝叶性状进行排序,叶性状与第1排序轴的相关性较高,枝性状与第2排序轴相关性较好。在第1排序轴中,除比叶面积外,从左到右,枝叶性状增加,幼树有着较高的比叶面积值,而大树除比叶面积外,其他枝叶性状较高;在第2排序轴中,从下到上,枝性状逐渐增加,叶性状逐渐减小,幼树有着较低的枝叶性状。这意味着更高发育阶段的个体有着更高的枝叶性状。
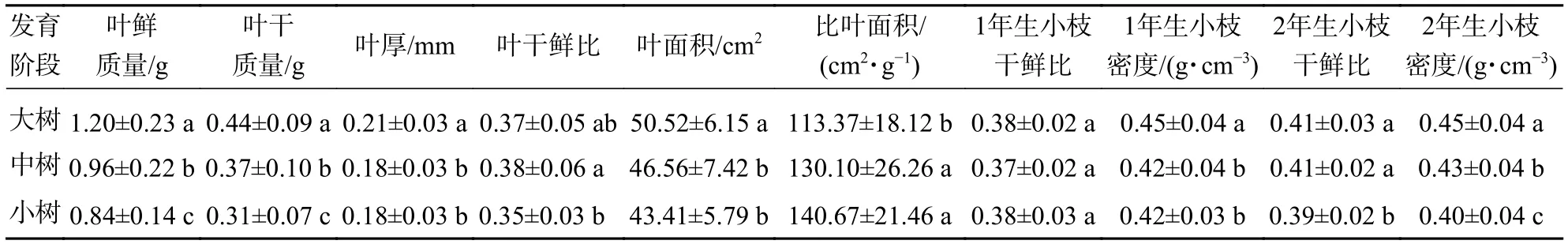
表1 不同发育阶段枝叶性状特征Table 1 Traits of twig and leaf in different development stages

表2 枝叶性状与胸径之间的相关系数Table 2 Correlation coefficient between twig, leaf traits and DBH
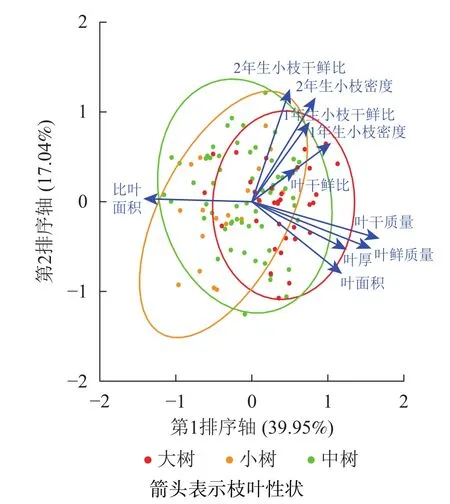
图1 不同发育阶段枝叶性状主成分排序图Figure 1 PCA ordination of twig and leaf traits at different developmental stages
2.2 不同发育阶段枝叶性状相互关系
标准化主轴分析发现(表3):叶性状之间及枝性状之间有着较强的相关关系,叶性状与茎性状之间相关性较弱,仅叶鲜质量和叶干质量与小枝性状具有相关性。主成分分析(PCA)也发现 (图1):叶性状与小枝性状存在正交关系。从表4~6可以看出:不同发育阶段叶性状间相关显著(P<0.05),其中,大树和中树阶段叶性状间相关性均比小树阶的段相关性强;不同发育阶段枝性状之间相关性从大到小依次为小树、中树、大树。枝叶性状之间相关性,除了小树阶段叶干质量和1年生小枝密度,大树阶段叶鲜质量和1年生小枝干鲜比、1年生小枝密度,中、小树阶段比叶面积和1年生小枝密度,中树阶段叶厚和1年生小枝密度显著相关外(P<0.05),其余均不显著(P>0.05)。
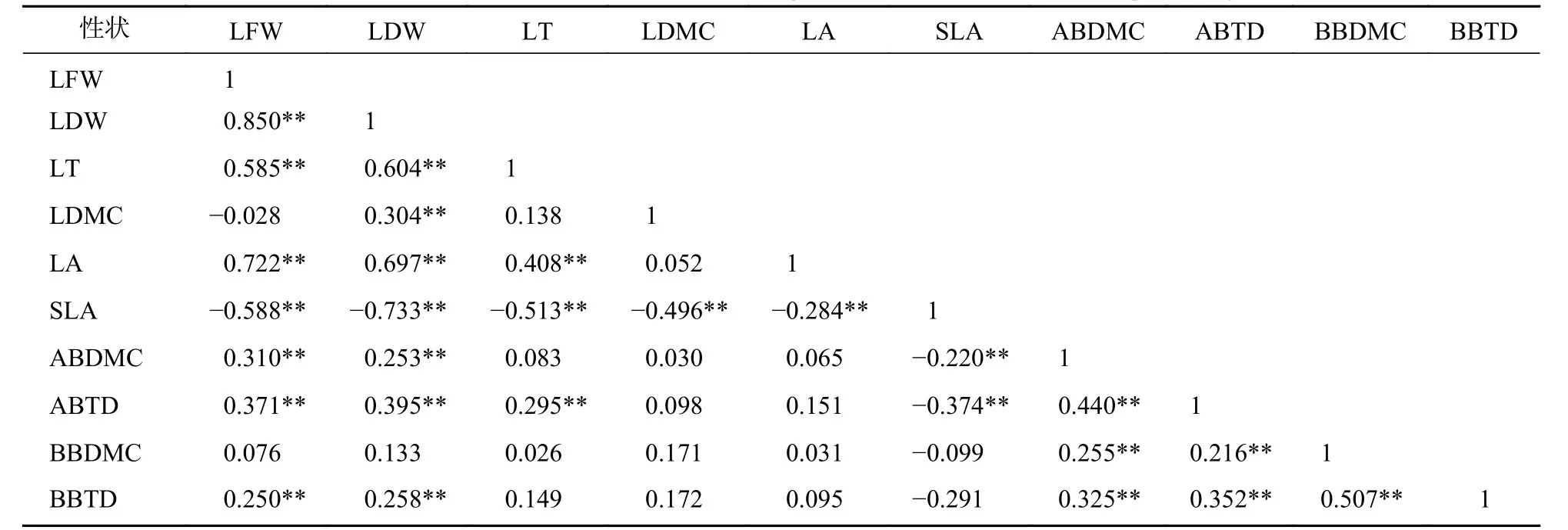
表3 整个发育阶段枝叶性状之间的相关系数Table 3 Correlation coefficient between the twig and leaf traits in the whole development stage
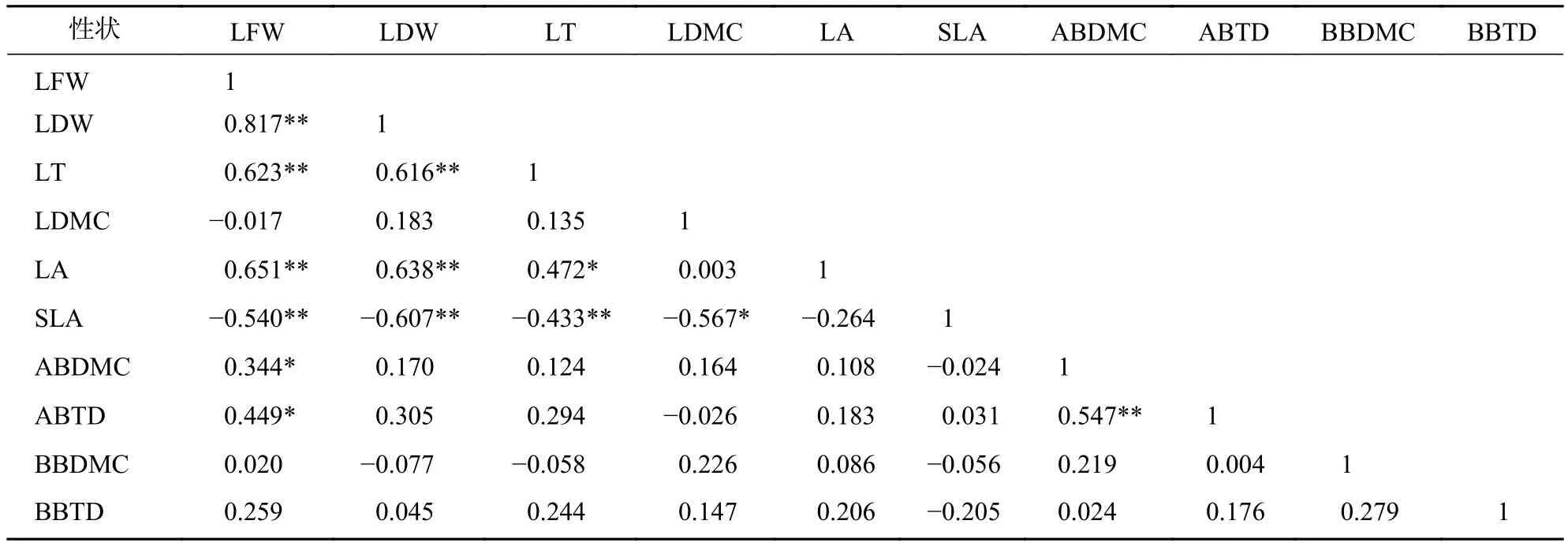
表4 大树阶段枝叶性状之间的相关系数Table 4 Correlation coefficient between the twig and leaf traits in big tree stage
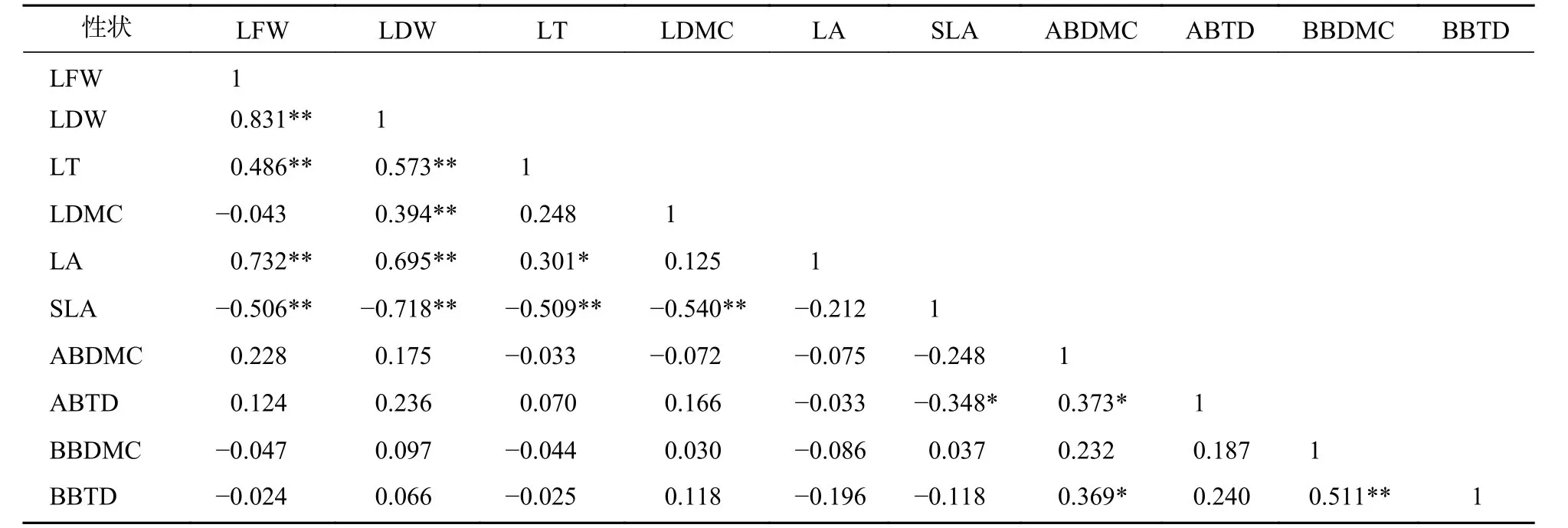
表5 中树阶段枝叶性状之间的相关系数Table 5 Correlation coefficient between the twig and leaf traits in middle tree stage
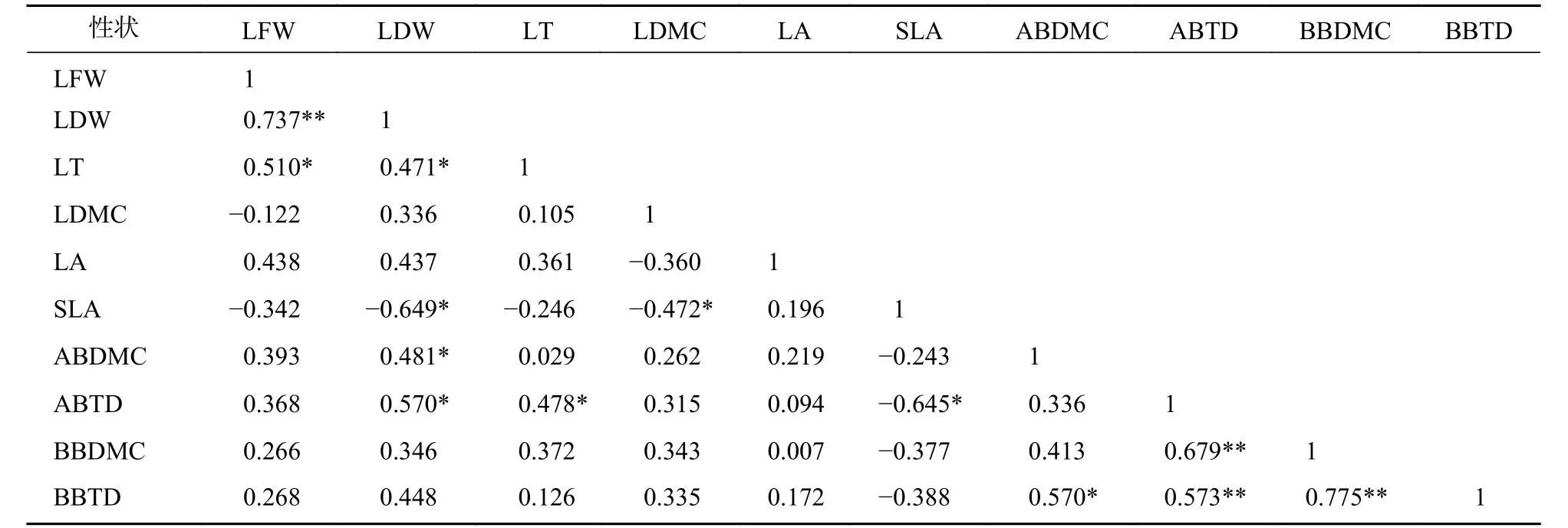
表6 小树阶段枝叶性状之间的相关系数Table 6 Correlation coefficient between the twig and leaf traits in small tree stage
2.3 不同发育阶段枝叶性状半方差分析
在不同发育阶段枝叶性状中,选择第1轴的林木个体枝叶性状主成分得分进行半变异函数方差分析(表7和图2)。对整体林分而言,单木整体性状、叶性状和枝性状的结构方差比分别为25.2%、31.8%、0,单木整体性状和叶性状具有中等程度的空间自相关,枝性状空间自相关性弱,拟合最优模型为线性,说明其性状表现呈随机分布。在大树阶段,单木整体、叶性状和枝性状的结构方差比分别为50.5%、50.4%和50.5%,具有中等的空间自相关程度,有效变程分别为75.60、73.70和212.70 m,拟合最优模型分别为球状模型、球状模型和指数模型,呈聚集分布,枝性状的有效变程最大,说明枝性状的空间连续性更大。在中树和小树阶段,模型拟合效果差,且结构方差低,说明小树阶段枝叶性状空间自相关性弱,不宜采用克里格插值(Kringing)空间预测。

表7 枝叶性状半方差模型及参数Table 7 Isotropic semi-variogram model and parameters of twig and leaf traits
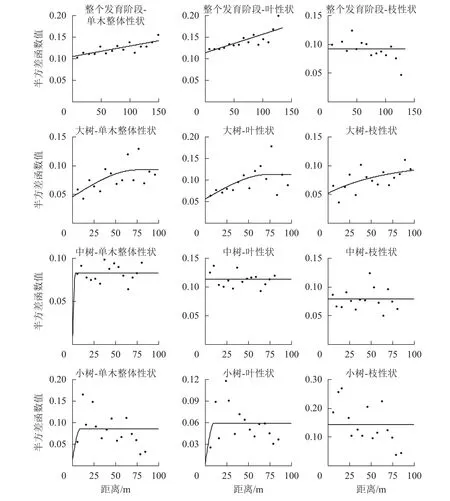
图2 枝叶性状半方差函数图Figure 2 Isotropic semi-variogram of twig and leaf functional traits
3 讨论
3.1 不同发育阶段枝叶性状的差异
植物之间的生存策略方式会在植物的性状上表现出来。比叶面积与植物的光合利用效率有关[19],本研究中比叶面积随着林木大小增加显著降低,这与其他不同物种及生境下得到的研究结果一致[20]。耿梦娅等[21]通过对不同发育阶段叶性状的研究发现:发育后期的植物个体有着更大更厚的叶片,且不同的发育阶段叶片性状差异显著,与本研究结果相符。较高的林木在进行长距离水分运输时,必须克服阻力问题,而较高的茎干鲜比将有利于水分的运输[22]。本研究发现:枝干鲜比与林木大小的关系不显著,可能是研究区内湿度高,水资源没有成为限制植物生长的环境因子,这与HE等[23]的研究结果一致。研究还发现:随着林木的发育,小枝密度逐渐增大,高密度的小枝除了能够加强对外界环境的抵抗外,还能够承载更大更多的叶片;较低的小枝密度意味着有更低构造细胞的成本,有利于小树阶段枝条的快速生长[24]。这反映了种群的生态策略方式从小树到大树阶段由快收益向慢收益转变,这是物种的生存策略所决定的。自然界中,无论从单个物种的发育阶段还是不同生活型的物种来看,大部分矮小的林木有着小的叶片,高大的林木个体叶片和枝密度更高[25],与本研究结果相符。
3.2 枝叶性状间的相关性
本研究发现:在小枝与叶片的10个性状中,叶性状间的相关性强,与FORTUNEL等[26]的研究结果一致。部分性状间在小树阶段相关性较低或不显著,可能是在激烈的竞争环境下,由于生存策略的优化方式,不同器官间的权衡方式在各性状间独立运行的原因[27]。小枝与叶功能性状相关性较弱,仅小枝密度与叶鲜质量和干质量呈正相关,然而MÉNDEZ-ALONZO等[28]研究发现:茎密度与水分传导速率呈显著负相关,本研究结果与其相反,原因是茎的密度远大于小枝密度,足以支持林木生长的需要,因此更应该考虑长距离运输过程中高密度组织细胞带来的水分运输问题,这是茎与小枝不同的功能作用造成的。对茎叶经济谱的研究发现[9,29]:茎经济谱与叶经济谱呈正交状态,本研究中小枝与叶性状在排序轴呈正交状态,这是造成枝叶性状相关性弱的重要原因。叶片质量与枝密度呈显著正相关,较大的枝密度意味着小枝能够承受更多的质量负荷。对不同发育阶段叶片质量与枝密度之间的相关性分析发现:这种现象在大树阶段表现得更加显著,其原因是小枝密度与叶生物量密切相关[6],本研究也发现大树有着更大更厚的叶。通过研究4个枝性状之间的关系,两两之间呈正相关,较高的小枝密度需要更多的结构组织,这种现象在不同的发育阶段都有着不同程度的表现,小树阶段小枝性状间相关性更大,这主要是因为小树阶段的生物学特征决定的。关于茎经济谱[9]的研究表明:茎密度与茎干鲜比密切相关,本研究中小枝性状间也出现这种相关性。对于茎经济谱理论,同样也能适用于小枝部分性状的研究。
3.3 不同发育阶段枝叶性状空间特征
在地统计学分析中,整体林分的枝叶性状分布模式以随机分布为主,其空间自相关性为中等,其原因是天然林中阔叶树种的分布方式常以随机分布为主[29]。大树阶段,枝叶性状以聚集分布为主,刘妍妍[30]通过对阔叶林的空间格局分析发现:具有较大叶片的个体通常以聚集形式分布,与本研究相符。叶片和小枝性状是影响植物冠幅大小的重要因素,能够影响成年植物冠层的结构形式及发展状态,进而影响植物对资源的获取能力。大树阶段枝性状主成分得分的有效影响变程均大于整体和叶性状,说明枝条的开展情况比叶的大小更能加剧大树个体之间的影响。中、小树阶段枝叶性状的拟合效果和结构方差比较小,不成连续变化状态,中、小树阶段林木枝叶性状的预测不宜采用克里格(Kringing)插值估算,与SIEFERT[14]的研究不一致,主要原因可能是中、小树阶段的空间格局分布方式为随机分布,林木间的距离较远,空间自相关性弱。若要预测样地整体功能性状的表现,需要测量更多林木个体的功能性状值。
综上所述,不同发育阶段枝叶性状差异显著,大树倾向于高质量的投资策略,小树的投资策略为快速投资;不同发育阶段叶性状间和小枝性状间相关性显著;峨眉含笑种群大树阶段枝叶性状具有空间自相关性。
