Flight activity promotes reproductive processes in the fall armyworm,Spodoptera frugiperda
2021-02-25GEShishuaiHEWeiHELimeiYANRanZHANGHaowenWUKongming
GE Shi-shuai ,HE Wei ,HE Li-mei ,YAN Ran ,ZHANG Hao-wen ,WU Kong-ming
1 State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops,Institute of Applied Ecology,Fujian Agriculture and Forestry University,Fuzhou 350002,P.R.China
2 State Key Laboratory for Biology of Plant Diseases and Insect Pests,Institute of Plant Protection,Chinese Academy of Agricultural Sciences,Beijing 100193,P.R.China
3 College of Plant Protection,Henan Agricultural University,Zhengzhou 450002,P.R.China
Abstract The fall armyworm (FAW),Spodoptera frugiperda (Lepidoptera:Noctuidae),has invaded many countries in Africa and Asia since 2016,posing a major threat to world food security.Long-distance migration and strong reproductive ability form the biological basis of its rapid population expansion,but the relationship between the flight and reproduction of FAW has not been studied in depth.Here,an empirical assessment of this relationship in an invasive FAW population in China found that 1–3-day-old adults which had undergone 10-h tethered flights had a significantly shorter pre-oviposition period and greater oviposition synchronization,but did not show any differences in fecundity,oviposition period,mating percentage or other reproductive variables.Further studies on moths after 1.25–15-h tethered flights indicated that the reproductive process of adults could be fully triggered by flight activity longer than 2.5 h.Dissection of the reproductive organs also showed that tethered flight promoted ovarian and testicular development of FAW.These results show that appropriate moth flight activity significantly speeds up the reproductive process of FAW,which increases our knowledge on its migratory biology in relation to regional outbreaks.
Keywords:flight,reproduction,migration,invasion biology,movement ecology
1.Introduction
The fall armyworm (FAW;SpodopterafrugiperdaSmith)originated from the tropical and subtropical regions of America (Sparks 1979).In just the past few years,it has invaded many countries and regions in Africa,South Asia,Southeast Asia and East Asia (Goergenet al.2016;Sharanabasappaet al.2018; IPPC 2019; Wuet al.2019b).FAW larvae can damage many species of plants (Montezanoet al.2018),and the average fecundity of each female can reach up to 1 000 eggs (Muruaet al.2006).Currently,FAW has become a major agricultural pest in both the New and Old Worlds,threatening corn and other crops,and causing an estimated average yield loss of 12% (Baudronet al.2019).In December 2018,FAW made its first arrival in China(Sunet al.2021),and it quickly invaded most parts of the country,affecting 1.07 million hectares of China’s corn crop in 2019 (Jianget al.2019).Although FAW cannot overwinter through diapause in northern China,it reproduces normally in the winter season of southern China (i.e.,Yunnan,Hainan,Guangdong,Guangxi),where it forms a stable source population for northward migration in the spring season of the following year.
FAW is a typical migratory pest (Sparkset al.1986; Wolfet al.1995; Nagoshiet al.2008; Westbrooket al.2016),and its migratory behavior is an adaptive strategy for finding more favorable habitats (Dingle and Drake 2007).Migration enables the rapid spread of FAW,and its strong reproductive capacity guarantees a large population; but the migration is a high-energy process (Velde and Dyck 2013),which may only occur at the expense of some reproductive processes,such as reduced fecundity,extended pre-oviposition period and reduced longevity (Rankin and Burchsted 1992; Karlsson and Johansson 2008; Gibbs and Dyck 2010).Thus,the relationship between migration and the reproduction of migratory insects has been a focus of migration research in general,and results on the effects of migration on reproduction have been contradictory.According to the classic theory on the oogenesis-flight syndrome,there is a “switch” between migration and reproduction; the development of the ovarian system stops temporarily before migration,so that energy resources can be used to develop or maintain the physical flight system.After migration,the flight system degenerates and the energy substances can be used for the reproductive system (Johnson 1969; Liet al.2001; Polilov 2011).Whether the migration occurs before or during reproduction,the effect of migration on reproductive behavior is apparent.
Migration,as an integral part of the insect life cycle,plays an important role in population maintenance and evolution(Dingle 1996).Insects complete migration under suitable physiological and environmental conditions,and reduce the reproductive costs as much as possible.The reproductive cost often varies with insect species,and depends on factors such as migratory age and flight duration.For example,inGryllustextensis,Melitaeacinxiaand other insects,reproduction can be promoted after migration (Guerra and Pallack 2007,2009).Jianget al.(2010) found thatSpodopteraexiguaadults showed reduced mating and fecundity after a flight of 1-day-old individuals,but there was no effect on reproduction after flight in individuals of other ages.After a 6-or 12-h tethered-flight test,Cnaphalocrocis medinalis(Guenée) had a shorter pre-oviposition period and greater oviposition synchronization; however,tetheredflights over 18 h had no such effects (Zhanget al.2015).
At present,the reproductive cost of migration to insects is unclear,especially since the impact of migration on individual insects in experimental systems may differ from that on an entire insect population.For example,the longevity of insects may be reduced after migration,as a negative impact on the individual level,but at the population level it could increase the amount of energy available for reproduction.
Since the recent outbreaks of FAW occurred,this moth has become a hot research topic,especially regarding its control methods and insecticide resistance mechanisms(Kumelaet al.2018; Ramalhoet al.2018; Rivero-Borjaet al.2018; Cuiet al.2019; Tamboet al.2020).However,the flight biology of FAW has received less attention,as have the ecological significance of migration and its contribution to devastating epidemics of this pest.To clarify whether flight inhibits or promotes reproduction in FAW,we subjected adults to tethered-flight tests to determine the influence of simulated migration on reproductive processes,with the goal of better understanding FAW population dynamics during epidemics.
2.Materials and methods
2.1.Study insects
FAW larvae were collected in corn fields in Dehong Autonomous Prefecture (Yunnan,China) in January 2019,and reared in the laboratory on corn leaves until pupation.The offspring were maintained on an artificial diet of soybean flour and wheat bran for cotton bollworm(Helicoverpaarmigera) described by Lianget al.(1999)at (25±1)°C,(70±10)% relative humidity (RH) and 16 h light/8 h dark.The experimental population was drawn from different generations of this laboratory population.Male and female pupae were separated and placed in Petri dishes with moist cotton.After pupal emergence,male and female moths were kept separate to prevent mating,maintained in 30 cm×30 cm×30 cm cages and fed on a daily basis with 10% honey solution.
2.2.Experimental assays
Flight mill and tethered flightFAW adults were individually attached to a flight mill (FXMD-24-USB flight mill; Jiaduo Science,Industry and Trade Co.,Ltd.,Henan,China)in an artificial climate chamber (70 cm×50 cm×114 cm;MGC-450HP; Yiheng Technology Instrument Co.,Ltd.,Shanghai,China) using the method of Fuet al.(2017).Active,undamaged adults were gently collected from the rearing cage and lightly anesthetized with ether.Their wings were expanded,scales were brushed from the junction of the abdomen and thorax,and tethering was done using a small droplet of cyanoacrylate glue (Deli Group Co.,Ltd.,Zhejiang,China).After the moths recovered,only undamaged and active individuals were attached to the flight mill,and maintained at 25°C and 75% RH.The chamber was completely darkened and moths were allowed to engage freely in tethered flight.
Experimental designThe 1–3-day-old male and female moths were tested in a 10-h tethered flight,then removed from the apparatus and placed in pairs into a disposable plastic cup (500 mL) for mating (1 pair/cup).The total sample sizes of paired moths at the three ages were 45,52 and 54,respectively.The 1-day-old adults that had not flown were paired as the controls (1 pair/cup),and this sample size was 47.The top of the plastic cup was covered with sterile gauze to collect eggs,and a cotton ball dipped in a 10% (v/v)honey solution was placed in the cup for nutrition and water.The gauze and cotton were replaced daily.The length of pre-oviposition period (POP; interval from emergence to the first oviposition),daily fecundity,oviposition period,egg hatchability and longevity of the moths were determined.Mating status (mated/unmated) and mating frequency were determined by the presence and number of spermatophores in the female after death.
Because the flying ability of most 1-day-old moths is too weak to complete a 15-h flight,2-day-old male and female moths were selected for tethered flights of 1.25-,2.5-,5-,10-and 15-h durations,and total sample sizes of the five durations were 57,58,66,53 and 60,respectively.The reproductive variables described above were again determined for these moths.
We also carried out similar experiments to observe the anatomy of the reproductive structures of 1-day-old male and female moths after a 5-h tethered flight in comparison to 1-day-old control males and females that had not flown.The ovaries and testes of 3-,5-,7-and 9-day-old male and female moths were excised using a stereoscopic dissecting mirror(TS-75X; Shanghai Shangguang New Optical Technology Co.,Ltd.,Shanghai,China),ovarian development levels and female mating times were determined,and excised testes were photographed with a microscope (TS-63X;Shanghai Shangguang New Optical Technology Co.,Ltd.).The Old-SG Software (Shanghai Shangguang New Optical Technology Co.,Ltd.) was used to measure the testis major axis length.Ovarian development was assessed using the method of Zhaoet al.(2019),and testes were excised as described by Heet al.(2019).
2.3.Data analysis
Data were log-transformed (egg hatchability data were arcsine-transformed) before analyses to meet assumptions of normality and homogeneity for parametric analysis.Oneway analysis of variance (ANOVA) was used to test for differences in mean values of the reproductive variables and longevity of both sexes,followed by a post-hoc Tukey’s HSD multiple comparison test among the different groups.The Mann-WhitneyU-test was used to test differences in ovarian development levels between the control group and the flight group,and the Student’st-test was used to test differences in testis major axis length.The Chi-squared test was used to test for differences in mean mating percentage.The Levene test was used to test the homogeneity of variance of daily average fecundity between pairs of different groups.The significant difference level wasP<0.05.IBM SPSS Statistics for Windows,version 20.0 (IBM Corp,Armonk,NY,USA)was used for the analyses.
3.Results
3.1.Effect of young moth flight on reproductive performance
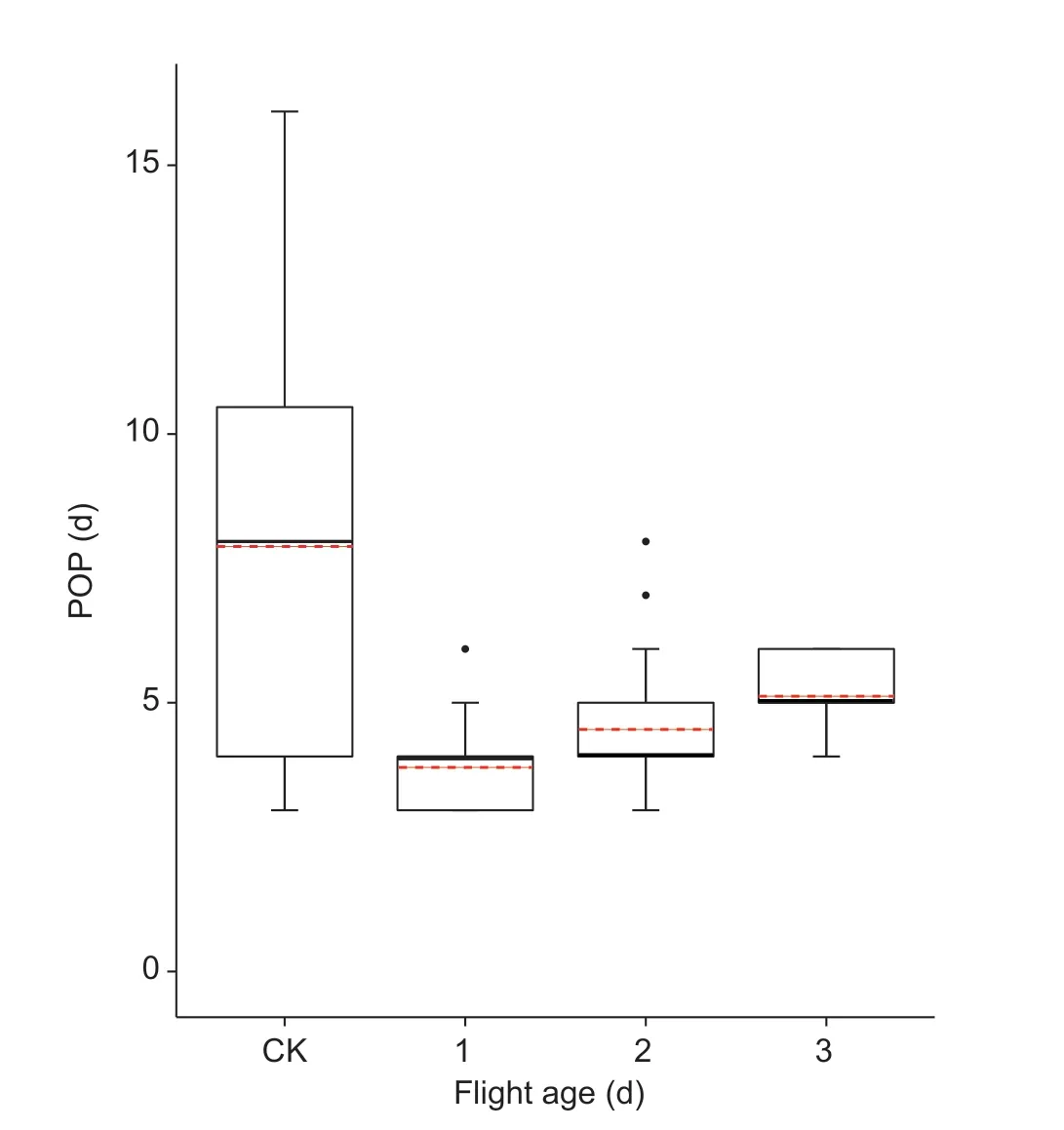
Fig.1 Pre-oviposition period (POP) of 1–3-day-old fall armyworm adults after a 10-h tethered-flight test.CK,no flight at 1-day-old.The bottom and top of the black box indicate the lower and upper quartile values,respectively; the horizontal solid black line shows the median for each category,while the red dashed line represents the mean.Whiskers indicate the maximum and the minimum; the black circles represent the outliers for the data set.Sample sizes of four groups are 23,25,28,and 26 from left to right.
The 1–3-day-old moths subjected to the 10-h tethered flight had significantly shorter POP compared with the control group(F3,98=17.06,P<0.001),but did not differ significantly from each other (Fig.1).The control group had the longest POP(mean±SE:7.91±0.84 days),and the 1-day-old moths that had flown had the shortest POP (mean±SE:3.79±0.16 days).In the control group,the range between the maximum and minimum POP values was 13 days,the difference between the upper quartile and the lower quartile was 7 days; while the differences between the upper quartile and the lower quartile of the three age groups were each 1 day.
Oviposition period,fecundity,hatchability,mating percentage and mating frequency did not differ significantly among the four groups (F3,98=0.42,P=0.74;F3,98=1.10,P=0.35;F3,98=1.04,P=0.38;χ2=0.28,df=3,P=0.96;F3,98=0.89,P=0.45,respectively) (Table 1).Whether females or males,the longevity differed significantly among the four groups (F3,194=7.66,P<0.001;F3,194=6.02,P<0.001).Female longevity of each of the three age groups after flying was significantly shorter than that of the control group,and the male longevity of the 1-day-old and 2-day-old groups was significantly shorter than that of the control group.There were no significant differences in both female longevity and male longevity among the three age groups.
Daily average fecundity of the three age groups had greater variations than the controls (Levene test:1-day-oldvs.CK,P<0.001; 2-day-oldvs.CK,P=0.001; 3-day-oldvs.CK,P=0.026),while comparisons between pairs of the three age groups showed no variations (1-day-oldvs.2-day-old,P=0.649; 1-day-oldvs.3-day-old,P=0.542;2-day-oldvs.3-day-old,P=0.805).In Fig.2,there were obvious oviposition peaks for the three age groups,but not for the control group.
3.2.Effect of moth flight duration on reproductive performance
Among 2-day-old FAW females,POP differed significantly after the five durations of tethered flights (F5,188=12.30,P<0.001) (Fig.3).POP did not differ significantly between the control group and the 1.25-h flight group,but was significantly longer for the controls than for the 2.5-,5-,10-and 15-h flight groups.The differences between the upper and lower quartiles for all flight groups were significantly shorter than that for the control group.
Mean oviposition period,fecundity,hatchability,mating percentage and mating frequency did not differ significantly among the flight durations for 2-day-old females (F5,188=0.34,P=0.89;F5,188=1.61,P=0.16;F5,188=1.70,P=0.14;χ2=2.17,df=5,P=0.83;F5,188=1.53,P=0.18,respectively) (Table 2).Longevity of females did not differ among flight durations,but they were significantly shorter than that of the control group (F5,335=3.83,P=0.002).Longevity of males after 10-h flights was significantly shorter than that of the control group (F5,335=6.86,P<0.001),while male longevity did not differ among the control group and the other flight groups.
All flight groups showed greater variation in daily average fecundity than the control group (Levene test:1.25 hvs.CK,P=0.021; 2.5 hvs.CK,P=0.012; 5 hvs.CK,P=0.043;10 hvs.CK,P=0.001; 15 hvs.CK,P=0.001).There were no significant variations in daily average fecundity between any of the flight durations (1.25 hvs.2.5 h,P=0.427; 1.25 hvs.5 h,P=0.948; 1.25 hvs.10 h,P=0.295; 1.25 hvs.15 h,P=0.270; 2.5 hvs.5 h,P=0.511; 2.5 hvs.10 h,P=0.904;2.5 hvs.15 h,P=0.889; 5 hvs.10 h,P=0.392; 5 hvs.15 h:P=0.368; 10 hvs.15 h,P=0.985).There were obvious oviposition peaks between all treatment groups,but not for the control group (Fig.4).
3.3.Effect of moth flight on development of reproductive organs
For 1-day-old male and female moths after a 5-h tethered flight,the development process of the female reproduction system over time was significantly accelerated compared with the control group (when 3-days-old:z=–5.661,P<0.001;5-days-old:z=–3.411,P=0.001; 7-days-old:z=–1.793,P=0.073; 9-days-old:z=–2.665,P=0.008),and the testis major axis length of 9-day-old male moths was significantly shorter than that of the control group (3-days-old:t=1.20,P=0.236; 5-days-old:t=0.46,P=0.645; 7-days-old:t=0.72,P=0.474; 9-days-old:t=4.21,P<0.001) (Table 3; Fig.5).In addition,based on the presence of spermatophores in the female moth,mating percentage for the control group was0% at 3-days-old and 18.75% at 5-days-old,compared with 33.33 and 58.06%,respectively,for the flight group.
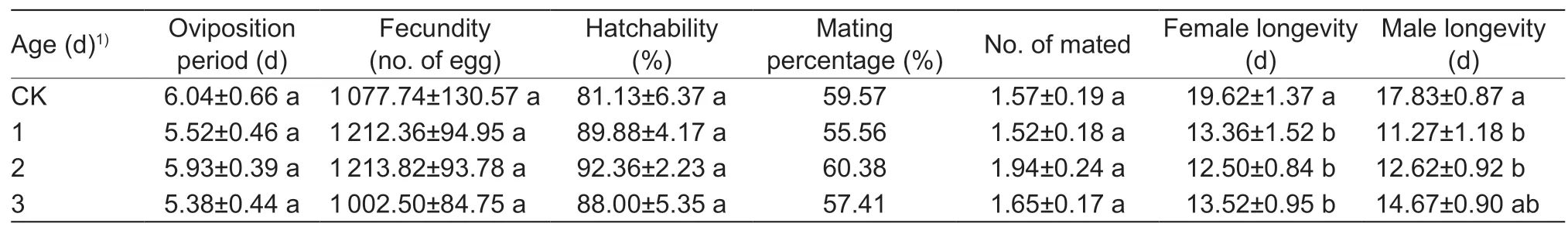
Table 1 Reproductive performance of 1–3-day-old fall armyworm adults after a 10-h tethered flight
4.Discussion
After a 10-h flight at the ages of 1 to 3 days,FAW females had a significantly shorter POP than those in the control group,and the oviposition synchronization was significantly enhanced,while the other reproductive characteristics did not change.Thus,flight activities by 1–3-day-old females promote reproductive activities,which is a significant advantage for migratory females.In a previous study,the flight capacity of 1-day-old FAW adults was shown to be weak (Geet al.2019),and the flight muscles may not be fully developed enough for long-distance migration.Therefore,2–3-days-old may be the most suited for migration.For 2-day-old females after varying durations of tethered flight,except for the 1.25-h flight,POP was significantly shorter than that for the control group and the oviposition synchronization was significantly enhanced,while the other reproductive variables did not change.All flight durations significantly reduced female longevity compared to the controls,such was similar to the migratory pestsCnaphalocrocismedinalisandMythimnaseparata(Lv 2014; Zhanget al.2015),but different fromS.exiguaandLoxostegesticticalis(Jianget al.2010; Chenget al.2012).
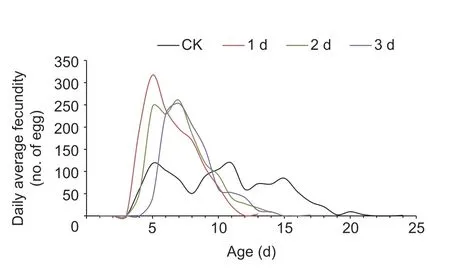
Fig.2 Daily average fecundity curves of female moths after a 10-h tethered flight.CK,no flight at 1-day-old; 1 d,10-h tethered flight at 1-day-old; 2 d,10-h tethered flight at 2-days-old; 3 d,10-h tethered flight at 3-days-old.
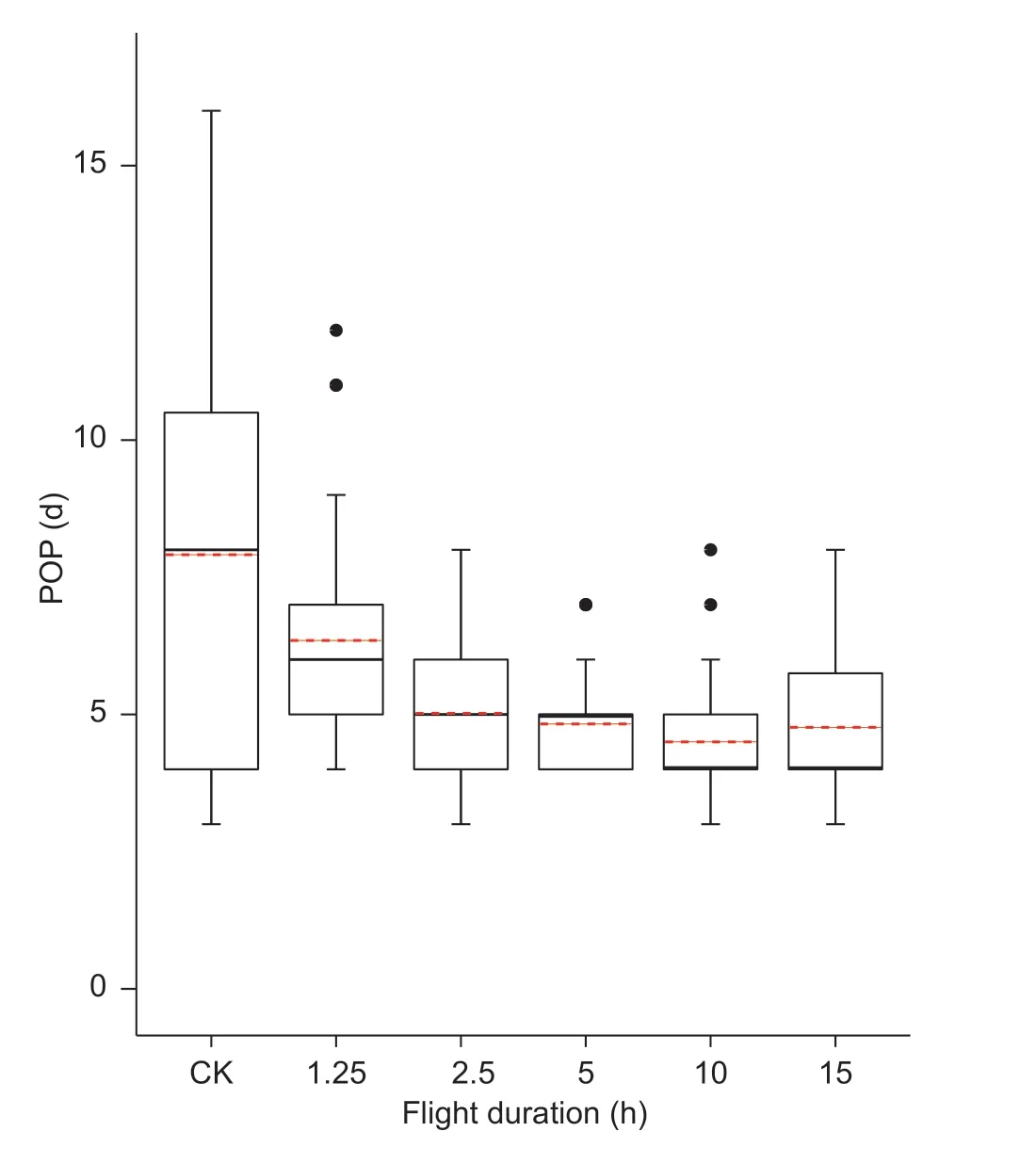
Fig.3 Pre-oviposition period (POP) for 2-day-old female adults after different durations of tethered flight.CK,no flight at 1-dayold.The bottom and top of the black box indicate the lower and upper quartile values,respectively; the horizontal solid black line shows the median for each category,while the red dashed line represents the mean.Whiskers indicate the maximum and the minimum; the black circles represent the outliers for the data set.Sample sizes of the six groups are 23,34,40,39,28 and 30 from left to right.
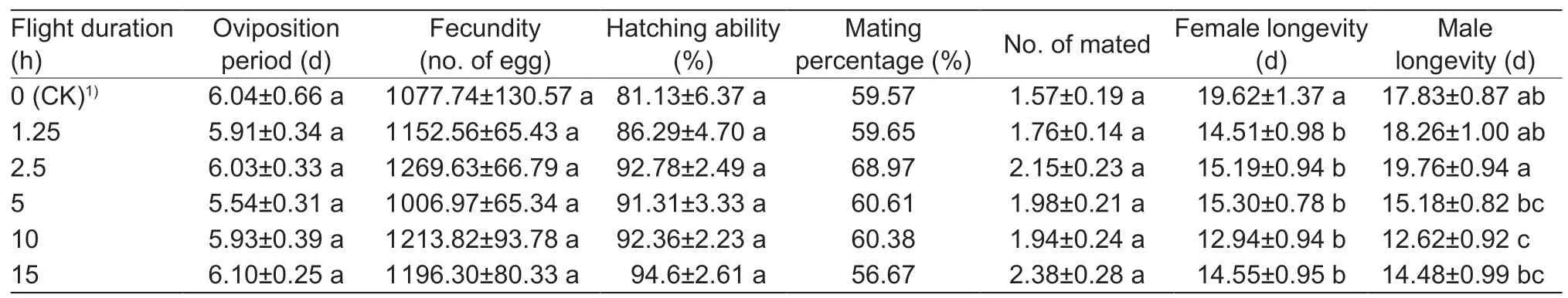
Table 2 Reproductive performances of 2-day-old fall armyworm moths after different durations of tethered flight
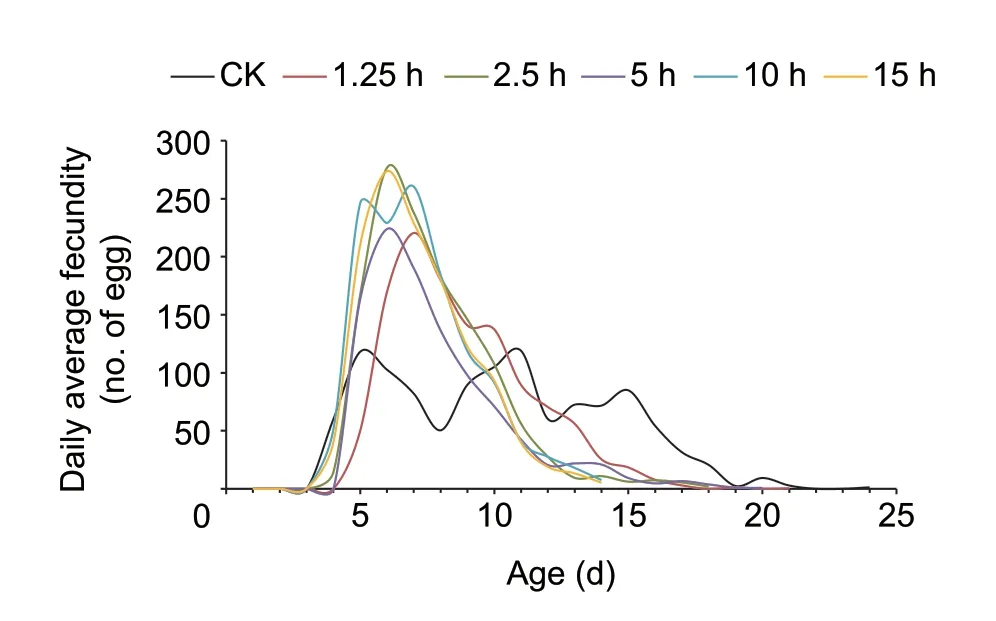
Fig.4 Daily average fecundity curves of 2-day-old female moths after different durations of tethered flight.CK,no flight at 1-days-old; 1.25 h,2.5 h,5 h,10 h and 15 h indicate 1.25-,2.5-,5-,10-and 15-h tethered flight at 2-days-old,respectively.
In our experiments,the fecundity of FAW did not change after the flights,but female longevity significantly shortened.We speculate that the unchanged fecundity is associated with the decrease of female longevity.The decrease of female longevity and POP will increase the proportion of energy that can support reproductive activities,maximizing the limited resources for reproduction,and ensuring the continuity of the population.This allocation of resources may thus be a long-term survival strategy for FAW.The fecundity is also related to supplementary nutrition of adults (Morales-Ramos 1996; Boggs 2010; Fanget al.2019).When the flights had been completed,we quickly replenished the food source,which would have made up for the energy consumption of flight to some extent and ensured that energy was supplied for subsequent reproduction.Although there was no significant effect of flight on the total fecundity after flight,the daily average fecundity was affected.After flight,there was an obvious oviposition peak,and more females were ovipositing at approximately the same time,which was related to the increase of oviposition synchronization.
The shortened POP was related to the acceleration of ovarian development after flight,as confirmed by observations of the ovaries.Flight accelerated ovarian development compared with the controls,especially in the early days after flight.In the flight groups,the ovarian development of 3-day-old individuals was mostly grade 2.0–3.0 (Table 3),and some had mated.In the flightless controls,ovarian development of 3-day-old individuals was mostly grade 1.0–2.0 (Table 3),and none had mated.Ovarian development in 5-day-old females was mostly grade 3.0–4.0 (Table 3),which indicated that they had entered the stage of oviposition; while ovarian development of the control group was mostly grade 2.0–3.0 (Table 3),and only 18% of them had mated.Liet al.(2005) found that flight improves the activity of the corpus allatum (CA) inMythimna separataand promotes the secretion of juvenile hormone(JH).Higher concentrations of JH are generally thought to promote ovarian development and inhibit the accumulation of triglycerides,which are used as an energy source for flight(Zera 2007; Jindraet al.2013; Liuet al.2016).Therefore,we speculate that FAW is similar toM.separata,and has a mechanism for flight that promotes ovarian development,thus speeding up ovarian maturation,mating and oviposition activities.Although the mating period occurred sooner in FAW after flight than in those that had not flown,flight did not significantly increase mating percentage and mating frequency,unlike the case forGryllustexensisandG.bimaculatus(Bertram 2007; Dyakonova and Krushinsky 2008).
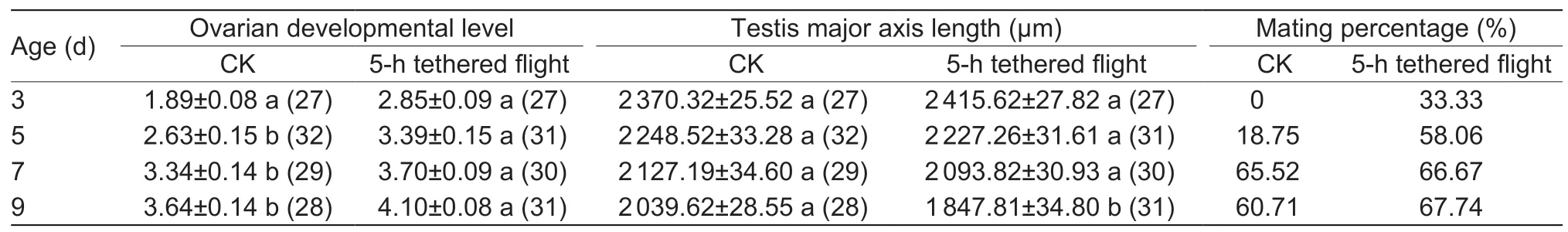
Table 3 Reproductive development and mating status over time of 1-day-old adults of control (CK,no flight) and after 5-h tethered flight
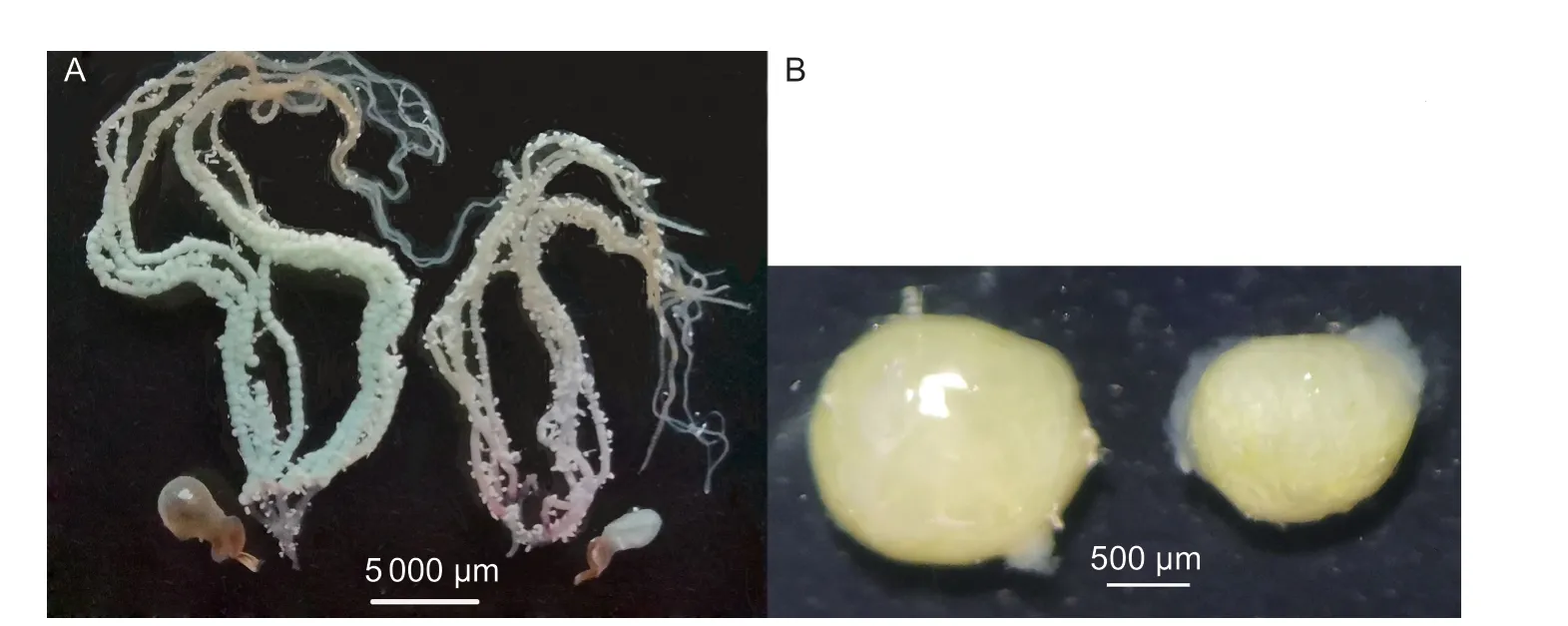
Fig.5 Ovarian and testis development of fall armyworm between control and flight groups.A,ovarian development of 3-day-old female moths for the flight group (left) and the control group (right).B,testis development of 9-day-old male moths for the control group (left) and flight group (right).
The reproductive system of FAW moths gradually matured with age,and appropriate flight activities accelerated this process in our study.Long-distance migration activities of some migratory insects promoted the ovarian development of female moths,such asScotogrammatrifolii,Agrotis segetum,MamestrabrassicaeandHeliothisviriplaca,which had migrated across the Bohai Sea and were captured by searchlights on a small island (Guoet al.2015; Wuet al.2015; Zhaoet al.2016; Heet al.2017).The ovarian development levels of the captured moths were high in May and June,which was due to the fact that the moths coming from the far south had a long flight distance and duration.In contrast,the ovarian development levels of the moths mostly were low in August and September,which was due to the fact that the moths coming from the near northeast had a shorter flight distance and duration.We speculate that these differences of the moths’ ovarian development levels between the two periods are mainly caused by both age and flight,which will be consistent with our research results.
After the different durations of tethered flight,oviposition synchronization of FAW was enhanced.We can speculate that both a short flight and a long-distance migration will enhance oviposition synchronization of FAW.Enhancement of oviposition synchronization will also promote synchronicity of larval herbivory during an outbreak of FAW.
The abundant food resources and monsoon climate in China provide good natural conditions for the cross-regional migration of FAW populations (Wuet al.2019a).Migration is generally a collective behavior,and migrating insects generally land together in a suitable environment; thus,flight enhancement of oviposition synchronization greatly increases the possibility of a rapid outbreak and extensive damage.Monitoring,an early warning system for potential outbreaks and control measures to avoid large-scale outbreaks of FAW are needed in China.
5.Conclusion
FAW has become an important global pest for corn,wheat and other crops,in large part due to its ability to migrate long distances and reproduce rapidly,which have allowed it to greatly expand its population across Africa and Asia within just a few years.Our empirical assessment of the relationships between flight and reproduction of FAW indicates that appropriate adult flights can promote the development of reproductive organs,shorten the POP and enhance the synchronization of oviposition.This basic foundation should help elucidate the factors that contribute to regional outbreaks caused by the migration of FAW.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31727901 and 31621064).
Declaration of competing interest
The authors declare that they have no conflict of interest.
杂志排行

Journal of Integrative Agriculture的其它文章
- Spread of invasive migratory pest Spodoptera frugiperda and management practices throughout China
- Biology,invasion and management of the agricultural invader:Fall armyworm,Spodoptera frugiperda (Lepidoptera:Noctuidae)
- Case study on the first immigration of fall armyworm,Spodoptera frugiperda invading into China
- Windborne migration routes of newly-emerged fall armyworm from Qinling Mountains–Huaihe River region,China
- Laboratory-based flight performance of the fall armyworm,Spodoptera frugiperda
- Adult nutrition affects reproduction and flight performance of the invasive fall armyworm,Spodoptera frugiperda in China