北京东灵山辽东栎林树木生长对气候要素的响应特征
2021-02-25李宗善陈维梁韦景树买尔当克依木张育新王晓春
李宗善,陈维梁,韦景树,买尔当·克依木,张育新,*,张 霜,王晓春
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085
2 中国科学院大学,北京 100049
3 东北林业大学生态研究中心,哈尔滨 150040
森林是陆地生态系统重要类型,森林生物量和生产力分别占陆地生态系统的45%和50%,森林生态系统可以固定人类排放到大气的二氧化碳总量的25%,是地球上重要碳汇[1-2]。森林生长对极端干旱气候响应也是非常敏感的,2000—2009年是器测观测以来最热的十年,期间发生的区域干旱事件导致全球森林净初级生产力(NPP)出现明显下降,下降速率达每年 5.5亿吨碳,且生产力下降最为明显的区域出现在热带亚马逊、印度尼西亚和非洲地区[3];2003年 欧洲热浪导致区域植被NPP下降30%,相当于植被减少5亿吨碳固定量,使欧洲地区从碳汇变成碳源[4]。已有大量研究表明,气候变暖引发的区域尺度干旱胁迫,已使得世界许多地区出现了森林生长衰退和树木死亡现象,严重削弱了森林生态系统固碳能力[5-6]。
华北地区属于暖温带半干旱半湿润季风气候区,降雨时空分布不均匀,年际间变化波动大,极端干旱年份发生频率也较高;20世纪80年代至今,华北地区降雨呈不断减少趋势,气候整体上呈现暖干化的趋势[7]。从器测1951—2013年时段,华北地区年总降雨量呈波动下降趋势,时段内线性减少趋势约为10%[7];从年轮和石笋重建的降雨波动历史来看,华北地区从20世纪中期开始的干旱时期在过去数百年尺度上也是较为明显的[8-9]。在2001—2014年时段,华北地区大部分区域植被覆盖度指数NDVI和植被NPP与干旱指数呈显著正相关关系,且以生长季夏季的相关程度最高,可见干旱气候胁迫已经成为华北地区植被生长最为重要的限制性因素[10-11]。
辽东栎是壳斗科栎属的落叶树种,是我国暖温带阔叶落叶林的主要优势种之一,在北京东灵山地区辽东栎林是分布最广的林型,是该地区的地带性植被,对暖温带落叶阔叶林的外貌、结构、动态及物种组成都有重要作用。目前对东灵山辽东栎群落区系组成[12]、植物多样性特征[13]、群落结构[14]、空间分布、植物功能性状[15]、群落动态和功能[16]都已经有了大量报道,然而关于辽东栎林的树木年轮学研究在东灵山地区还未引起人们足够的重视。本文基于北京东灵山辽东栎林的树木年轮宽度材料,分析了辽东栎树木生长对干旱胁迫气候的响应特征,为在全球变暖背景下该地区暖温带落叶落叶林的可持续保护提供科学依据。
1 材料与方法
1.1 研究区概况
北京东灵山位于北京市西郊的门头沟区(115°26′E—115°30′E, 40°00′N—40°03′N),与河北省毗邻,距市区约100 km,东灵山为小五台山余脉,属太行山系(图1)。海拔高度多在1000 m以上,最高峰为2303 m[13]。地质构造上位于华北陆台中部的燕山沉降带,地貌以山地侵蚀结构类型为主,地势陡峭,河流下切严重。地带性土壤类型为肥沃褐色土和棕色森林土[14]。该地区属于暖温带半湿润大陆季风气候,寒冷期长,积温低,冻土期长,生长季短,年均为5—10℃,无霜期约195 d,年降雨量为500—650 mm,多集中于夏季,春旱严重。植被为暖温带落叶落叶阔叶林,辽东栎为该区的地带性植被,主要分布在海拔800—1700 m范围,但由于人类活动的影响,辽东栎在海拔1000 m以下已不多见。本区植物区系丰富,共计127科、476属、997种,其重要来源是第三纪植物区系后裔,温带成为最为突出[12]。
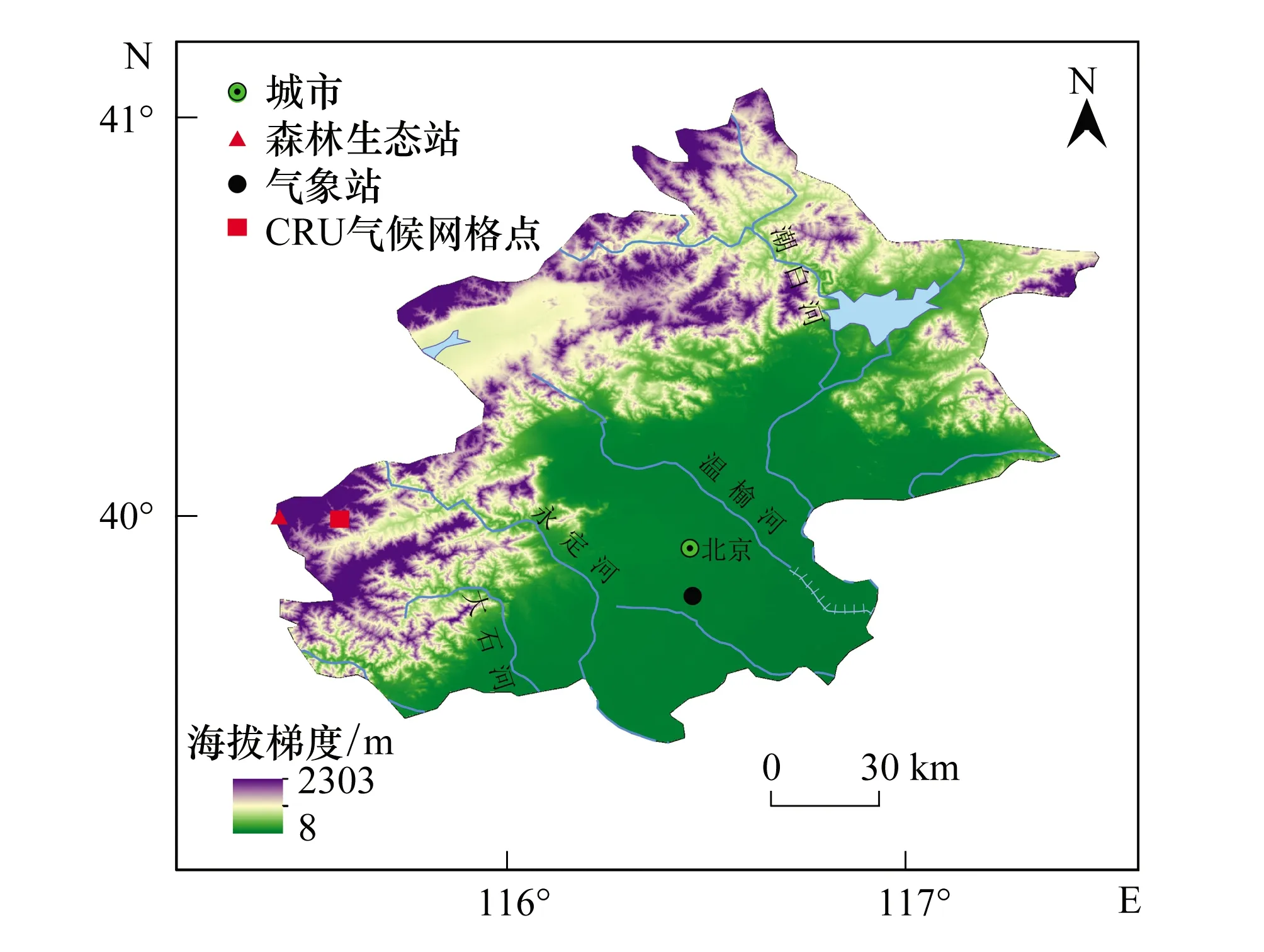
图1 北京东灵山树木年轮采样点和气象站分布图
1.2 年轮样品采集及处理
本文所用辽东栎树轮资料是2011年9月在北京东灵山东南坡采集的,采样地点(115.43°E, 40°N)在北京森林生态定位生态站东南方向约3 km,采样点海拔高度为1400 m。选择辽东栎林分布典型、受人为活动干扰较少的地段采集年轮样品;为获得较长的树木年轮样本,在树轮样品采集点选择年龄较老的树木个体获取样本。样芯采集通常在胸高部位,在垂直于山坡方向上用生长锥钻取树轮样本。取到的样芯放置在塑料样管内,并在样管上用油性笔标注代码。本研究每棵树采集一根树轮样芯,最终获得78根年轮样芯。
样品预处理基本按照Stokes和Smiley[17]的方法进行。首先将样品放置于平坦处晾干,然后将其粘在特制的木槽中;将粘牢的样芯依次用颗粒由粗到细(280—800目)的砂纸进行打磨。
1.3 树轮年表构建
样本经预处理后, 用骨架示意图进行交叉定年[18],然后使用LINTAB5.0年轮分析仪对树木年轮宽度逐年进行测量,该测量系统精度可达0.01 mm。利用COFECHA程序[19]对定年和测量结果进行了检验,保证定年与测量的准确性。对于那些效果较差、与主序列之间的相关系数达不到99%置信区的样芯予以剔除,最终66根辽东栎树芯用于年表的构建。
经过交叉定年的年轮序列,用ARSTAN程序[20]进行去趋势和标准化,这一过程消除了树木生长中与年龄增长相关联的生长趋势及部分树木之间的非一致性扰动,排除了其中的非气候信号。首先对每个年轮宽度序列进行负指数曲线拟合,以去除与树木年龄有关的生长趋势;如果不成功,则用任意斜率的线性回归进行直线拟合,并以取树轮宽序列和与其对应的生长趋势拟合曲线的比值,对所得到的去趋势年轮序列进行双权韧性平均处理,合成了平均值为l、无量纲的树轮宽度标准年表。
1.4 气象资料
北京东灵山的中国科学院北京森林生态系统定位研究站(Beijing forest ecosystem research station, BFERS)距离采样点最近(约5 km),但是该生态站气象数据时间较短且不连续,因而无法与年轮数据进行相关分析。鉴于此,本研究的气象数据选取离采样地点最近的北京气象站(国家标准气象台站,海拔313 m, 115.47°E, 39.8°N),其位于采样点东部约 80 km。气 候 资 料 气 象 数 据 由 国 家 气 象 局(http://cdc.cma.gov.cn/)提供,区间为1951—2010年,气候要素包括月平均气温(TMP)、月平均最高气温(TMX)、月平均最低气温(TMN)、月降雨量(PRE)。考虑到地面气象站点与采样地点的海拔一定差异,且具有一定距离,为了能够较好反映出研究地点气候特征,本研究选取了与采样点临近(约10 km)的CRU网格化气候格点数据,以期与地面气象站数据进行比对。CRU气候格点数据从CRU TS v.3.22 全球气候数据库(http://www.cru.uea.ac.uk/cru/data/hrg/)[21]下载,时间跨度为1951—2010年,精度为0.5 度(约为25 km),共下载4个气候指标,平均温度、最高温度和最低温度,降雨量和PDSI干旱指数。
1.5 数据分析
采用相关分析统计树轮生长对于气候要素的响应,考虑到前一年气候状况对当年树木生长的影响[18],所用气候资料的时间跨度为前一年7月到当年10月,气候要素包括月平均气温、月平均最高气温、月平均最低气温、月降雨量、PDSI干旱指数。为了能够显示树木生长与气候关系特征在时间轴上变化特征,又进一步统计了年表与气候数据的滑动相关分析,滑动窗口设定为30 a。以上固定时间段的相关分析及滑动相关分析均用DENDROCLIM2002 程序实现[22],相关统计显著性水平为P<0.05。
2 研究结果
2.1 研究区气候变化特征
因地面气象站距离采样点较远,海拔也有一定差异,为验证气象站数据的可靠性,对气象站数据与CRU网格点数据进行了校对分析(图2),结果表明气象站年际温度和年总降雨量与对应的CRU温度(R2=0.94,P<0.01)和降雨(R2=0.61,P<0.01)数据均具有显著相关关系。利用两者之间的回归关系。利用气象站与CRU数据间的回归关系,建立预测CRU年际温度(Y=0.79X-2.14)和年总降雨量(Y=0.47X+205.15)的线性模型,发现CRU数据与预测序列具有较好的吻合度,这说明气象站数据具有较好的空间代表性,可以反映研究区的气候变化特征。
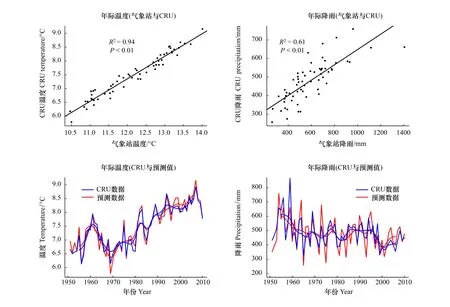
图2 北京气象站和英国气候研究所(CRU)网格点气候数据校对(1951—2010)
从气象站过去60 a(1951—2010)气候数据来看(图3),研究区气候总体上向暖干化趋势发展, 平均温度(TMP)呈显著上升趋势,每10年上升0.41℃,其中最低温度(TMN,每10年上升0.52℃)上升速率明显高于最高温度(TMX,每10年上升0.2℃),而降雨量(PRE)和相对湿度(RH)则呈不断下降趋势,每10年分别下降44.32 mm和14.8%。研究区附近的CRU网格点的气候数据变化趋势与气象站点数据具有一致规律,不同温度指标均有线性上升的趋势,而降雨量和PDSI干旱指数则有不断下降的趋势,气候暖干化特征非常明显。

图3 北京东灵山气象站及CRU格点气候数据波动趋势
2.2 树轮年表统计特征
本研究构建的辽东栎树轮宽度标准年表序列(简称年表)时间长度为85 a(1926—2010年)(图4),年表的起始年份(1926年)的最低复本量为5根样芯,构成该年表66根样芯的平均长度为67 a,其中,超过60 a的年轮序列有44条,超过80 a的年轮序列有10条,所以该年表可较可靠地记录东灵山地区过去数十年的树木年际径向生长的波动特征。
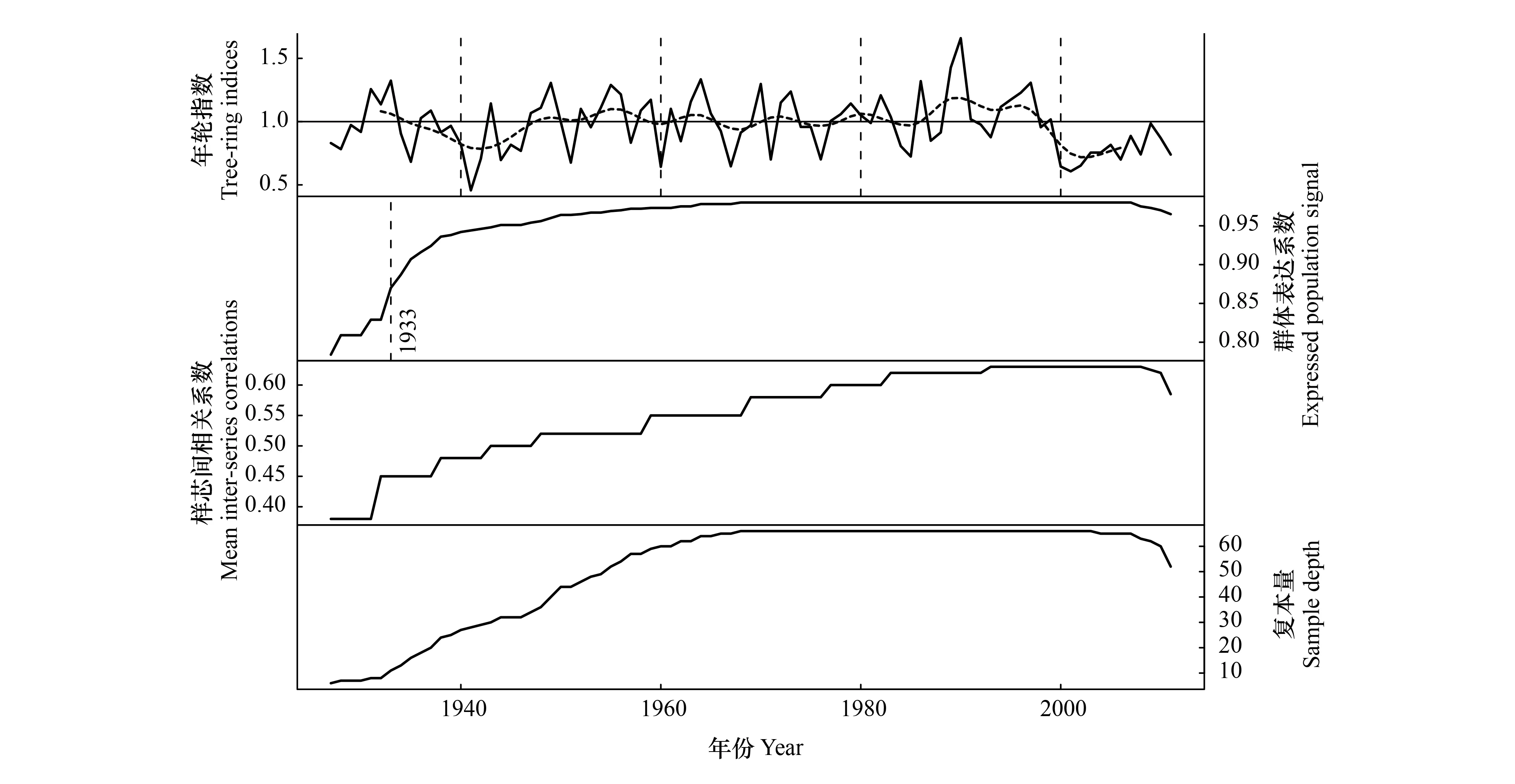
图4 北京东灵山辽东栎树轮标准年表
年表的统计量特征可以反映树木生长的一些基本特征以及树轮年表所含不同频率信息量的多少(表1)。结果表明,不同树木年轮样本间具有较好的相关性(样芯间相关系数Rbar=0.623,P<0.01),表明不同样芯之间的轮宽变化有很好的一致性。各年表的一阶自相关系数为0.236,说明树木前一年的生长状况对当年生长的影响较小[18]。年表的平均敏感度(MS)数值为0.322,这说明了树轮资料具有较强的高频波动信号。年表的信噪比(SNR)和第一主成分解释方差量(VFE)数值分别为30.919和42.4%,均达到了较高的水平;年表具有较高的样本群体表达信号(EPS),数值为0.969,这说明了本调查采集样本量所含的信号是能代表总体特征的[23]。以上年表各种统计特征表明了样本之间的树轮宽度变化有很好的一致性,记录了可靠区域气候信息,所以本次调查所用的树轮资料适于进行年轮气候学分析。

表1 北京东灵山树轮采样点概况、标准年表主要特征参数及共同区间分析结果
2.3 树木生长对气候的响应特征
与气象数据分析结果表明(图5),年表主要与当年夏季温度(7—9月)温度保持负相关关系,其中以平均温度(R=-0.271 —-0.341,P<0.05)和最高温度(R=-0.263 —-0.33,P<0.05)相关性最为明显,与最低温度的相关性较弱,仅与夏季7月份(R=-0.312,P<0.05)有显著负相关关系;年表与去年夏季温度也有较为明显的负相关关系,只是相关程度与当年夏季温度相比略低。另外,当年春季5月份温度对树木生长也有一定影响作用,主要体现在与平均和最高温度的负相关关系上(R=-0.24 —-0.259,P<0.05)。年表与当年夏季7月份降雨量(R= 0.311,P<0.05)和相对湿度(R= 0.307,P<0.05)则有较为明显的正相关关系,另外年表与去年夏季的降雨和相对湿度也有较强的正相关关系。

图5 北京东灵山辽东栎树轮年表与器测和CRU格点气候要素相关分析
年表与CRU网格点气候数据的相关性结果与器测数据具有较好的一致性(图5),与夏季(7—9月)温度主要体现出负相关关系,与夏季(7月)降雨主要体现出正相关关系。另外,年表与PDSI干旱指数则普遍保持正相关关系,其中年表与夏季(7—9月)月份PDSI的相关性(R= 0.302—0.315,P<0.05)最为显著。
2.4 树木生长对气候响应的滑动相关分析
从年表与气候要素的滑动相关分析结果来看(图6),年表与夏季(7—9月)温度在时间轴上一致维持较为稳定的负相关关系,且在最近负相关性有逐渐加强的趋势;年表与夏季6月份温度的相关性在2000年前后,由正相关关系转变为负相关关系。年表与夏季8月和秋季9月降雨在时间轴上基本维持正相关关系,且这种正相关关系在最近趋于加强;年表与相对湿度和PDSI干旱指数的滑动相关分析结果较为相似,均由2000年之前的负相关关系,转为2000年之后正相关关系。

图6 北京东灵山辽东栎树轮年表与器测和CRU格点气候要素滑动相关分析
3 讨论
树轮年表与气象站和CRU网格点气候数据相关分析均表明(图5),树木径向生长与夏季(7—9月)温度指标基本上呈负相关关系,而与夏季(7月)降雨和夏季(8—9月)PDSI干旱指数则基本上呈正相关关系,这表明夏季水分胁迫条件是限制北京东灵山地区辽东栎树木生长的主要限制性因素。水分条件对树木生长的限制作用在我国西北地区均可得到验证,如在山东蒙山地区黑松(Pinusthunbergii)径向生长变化发现与当年4—9月降水量呈显著正相关,当年5—7月高温对黑松径向生长变化产生显著抑制[24];陕西华山地区华山松(Pinusarmandii)对初夏4—9月的水分条件响应最为敏感[25],而春季至初夏的干旱条件则是内蒙古中部的油松(Pinustabulaeformis)径向生长主要限制性气候因子[26]。正因为我国西北地区树木生长受水分条件限制性较为突出,该地区树木年轮材料也常用来重建该地区过去数百年来季节或年际尺度降雨和干旱的历史波动特征[27-30]。基于2001—2014年区间植被遥感数据,华北地区的植被指数NDVI和植被生产力NPP在大部分地区与干旱指数呈正相关,且在夏季干旱指数相关程度最高,表明夏季干旱是影响植被生长状况的重要影响因素[11],这与本文的研究结果相吻合。
夏季处于树木生长最为旺盛的时段,夏季水热条件一般对树木生长起着重要的作用,夏季温度过高会造成树木蒸腾作用加强,并加快土壤蒸发失水量,土壤水分有效性显著下降,直接导致树木光合作用能力下降,导致树木生长减速[31-32];另外,夏季夜间较高温度条件还会使得树木的呼吸作用加强,会大量消耗树木白天合成的营养物种,从而对树木生长起到抑制性作用[33]。夏季降雨偏少则直接降低土壤湿度,夏季高温和低降雨量引发的干旱胁迫会显著降低树木的光合作用能力,从而导致窄轮出现[34]。树木生长还与春季5月份温度呈一定负相关关系,春季5月份温度已经达到较高水平,但是同期的降雨量往往是偏少的,高温增强土壤蒸发和植被蒸腾失水能力,土壤湿度下降,树木根系得不到充足的水分而不利于树木生长[35-36]。
树轮年表与气候要素相关分析结果还表明(图5),树轮年表还与去年夏季的气候条件具有一定相关特征,体现为与去年夏季温度(8—9月)的负相关关系,以及与去年夏季降雨(8月)和相对湿度(8月)的正相关关系,这说明夏季气候条件对树木生长影响具有一定的时滞效应[32]。夏季高温少雨的气候条件会显著增加树木的蒸腾和呼吸作用,导致土壤中水分和养分含量下降,势必大量消耗树干储存水分及营养物质来满足树木生理代谢需要,导致冬季生长休眠时期储存的水分及营养物质下降,直接影响树木第二年春季的正常生长[37-38]。
气象站数据表明,北京地区在过去60年间(1951—2010年)年平均气温由10.8℃上升至13.2℃,每10年上升0.41℃,是世界气温平均上升速率(每10年上升约0.2℃)的2倍,也要明显高于中国地区气温上升速率(每10年上升约0.3℃)[39-40]。北京地区则年降雨量则由780 mm 下降至530 mm,降雨量每十年下降幅度为5.64%,有研究表明华北地区年总降雨量在1951—2013年时段呈波动下降趋势,降雨量每10年下降幅度约为1.97%[7],可见北京地区降雨量下降幅度要明显大于华北地区平均水平。北京地区气候有明显暖干化趋势,水分匮乏导致的干旱胁迫条件将成为限制该区域植被生长的重要环境因子[7, 9, 25]。滑动相关分析表明,年表与夏季温度的负相关性和夏季降雨及PDSI干旱指数的正相关性在2010年以后明显加强,这主要是因为研究区干旱化程度在2010年以后明显加剧的结果(图6)。从气象站数据来看,研究区年降雨量从1951—1999年时段的628 mm下降到2000—2010年时段的426 mm,下降幅度达32.3%;干旱指数PDSI则从1951—1999年时段的0.298下降到2000—2010年时段的-2.2,气候向干旱化发展趋势明显[25]。干旱化气候条件已导致树木生长在2000年以来出现明显抑制,树轮指数从1951—1999年时段的1.046下降到2000—2010年时段的0.762,下降幅度达23.7%,表明北京东灵山辽东栎径向生长受干旱胁迫影响在2000年后有强化趋势。
辽东栎木材解剖结构属于环孔材,早材导管较大,导管直径最大在200—300 μm左右,是普通散孔材树种导管直径(50—60 μm)的4—5倍,具有较高的导水效率和蒸腾能力[41],辽东栎林在生长季的林分总蒸腾耗水量可到200 mm左右,是人工山杨林和刺槐林林分蒸腾量的3.5倍和1.5倍左右[42-43]。然而,辽东栎较大的导管直径,使其在负水势条件下导管内腔容易产生塌陷,引起木质部空穴化和栓塞,从而产生水分传导障碍(Hydraulic failure),导致树木生长出现抑制[44];与散孔材树种相比,辽东栎木质部导管密度较小、导管连接度低,在低水势条件下形成栓塞的导管,水分很难通过相邻导管或相邻导管壁纹孔进行水分运输,从而加重了栓塞对树木生长的影响作用[45]。另外,辽东栎树木木质部栓塞脆弱曲线呈“r”形,意味着当水势降低到某一阈值时,其木质部的导水能力急剧下降,木质部导管很快发生栓塞,木质部50%导水率损失的压力值(P50)仅仅约为0.5 MPa[46-47]。由此可知,辽东栎对干旱胁迫承受能力较弱,在暖干化气候不断发展条件下,木质部栓塞发生可能性大幅上升,从而导致水力传导障碍,限制树木生长和发育,这可能是树木生长与夏季干旱胁迫关系随时间逐渐趋于加强的重要原因。
4 结论
全球气候变暖导致的干旱胁迫已对区域尺度森林生长产生了深远影响,森林生长对干旱胁迫的敏感性程度明显上升,而对温度响应敏感性出现下降趋势,导致受世界范围受水分条件影响和控制的森林面积明显扩大[47-48],干旱胁迫已导致很多区域出现森林生长衰退和树木死亡问题,干旱胁迫对森林生长的负面响应在很大程度上抵消了气候变暖和二氧化碳浓度上升对森林生长的正面效应[49]。基于树木年轮宽度资料,本研究发现北京东灵山辽东栎森林生长主要受干旱胁迫条件影响,而且干旱胁迫对树木生长的控制作用随时间有逐渐加强的趋势。已有研究表明东灵山辽东栎种群密度在1991—2002年时段出现明显下降趋势[16],本研究显示辽东栎树轮年表在1990—2010年期间处于明显生长下降时期;若未来气候继续向暖干化方向发展,北京东灵山地区森林生长受干旱胁迫影响将进一步加剧,从而对该地区森林植被生产力和固碳潜力产生明显负面影响。
