石河子引种文冠果不同株系抗寒性的鉴定及分析*
2021-02-24连琼宇李照然兖攀姜继元叶春秀
连琼宇,李照然,兖攀,姜继元,叶春秀
(1.新疆农业大学 林学与园艺学院,新疆 乌鲁木齐 830000;2.新疆农垦科学院林园所,新疆 石河子 832000)
文冠果(XanthocerassorbifoliaBunge)属无患子科(Sapindus Linn.)文冠果属(XanthocerasBunge)的落叶灌木或小乔木,素有“千花一果”称号。种子黑褐色,种仁含油率高[1]。文冠果木材纹理致密,是制作家具、农具和手工艺品的好材料。文冠果树形优美,是园林绿化及蜜源植物,其具有很高的观赏价值和生态效益[2]。主要自然分布区为暖温带大陆性气候区,以内蒙古、河北、河南、山西、陕西、甘肃6省(区)的资源较多,其根系发达、保水力强,耐干旱、严寒和贫瘠,是山区绿化、退耕还林和防风固沙的首选生态树种,具有巨大的开发和利用潜能[3]。近年来其引种范围越来越大,在生态利用与保护方面的价值也发挥得越来越充分[4],在引种研究过程中,相关研究[5]表明低温胁迫是文冠果引种过程中需要考虑的重要生态因子,低温不仅导致生长受到严重损害,甚至导致植株死亡。植物在低温胁迫下,膜脂相变及膜脂过氧化产物丙二醛含量升高[6],细胞膜透性增大[7],渗透调节物质(可溶性糖、可溶性蛋白和游离脯氨酸)积累保护酶活性升高[8]。目前对于植物抗寒性研究主要集中在杨树(PopulusL.)、杏(ArmeniacavulgarisLam.)、李(PrunussalicinaLindl.)等树种,对于文冠果抗寒性的研究较少[9-11]。曾祥谓对文冠果连续3 a进行了抗寒性测定,发现其叶片相对含水率82%,抗寒性能力中等[12];曹振岭等[13]证实文冠果能经受-35.3 ℃的特大冻害;马新等[5]发现低温伤害使文冠果细胞膜的透性增大,细胞内形成大量可溶性物质增加渗透势,参与植物的抗寒过程。曹振岭等[14]提出引种文冠果时要注意纬度不跨越过大,距离越远,引种成功的机率越低。本研究在前期形态学等指标筛选的基础上,筛选出6个性状表现优异的株系进一步分析其抗寒性,以期为文冠果是否能在石河子地区进行种植和推广作出科学评价提供参考。
1 材料与方法
1.1 试验区概况
试验设在新疆农垦科学院林园所试验地,位于新疆石河子市(44°17′N,85°51′E),属典型的温带大陆性气候,日照时间长,冬季长而严寒,夏季短而炎热,地处天山北麓中段,准噶尔盆地南部,平均海拔高度450.8 m,年平均气温7.5~8.2 ℃,年日照时数2 721~2 818 h,空气干燥。年降水量180~270 mm,年蒸发量1 000~1 500 mm,无霜期168~171 d。试验地土壤为灌耕灰漠土,耕层为0~20 cm,有机质16.7 g/kg,全氮1.13 g/kg,速效磷23.87 mg/kg,速效钾246.8 mg/kg。
1.2 材料采集及处理
在前期形态学指标的分析及主成分综合评价筛选出‘G 27’‘BD特异’‘G 13’‘株系2’‘建2’‘BD 12’这6个文冠果的株系作为抗寒性试验材料。于2019年11月下旬采集1 a枝条,每个株系采集30个枝条,将采集的枝条用自来水冲洗其表面尘埃,然后用蒸馏水冲洗3遍,用吸水纸吸干水分后剪成长度10 cm的小段,贴上标签放入密封袋保存备用。以4 ℃为对照,设置-10、-20、-30 ℃低温梯度在人工气候箱中进行低温胁迫处理,每个温度胁迫12 h,然后置于温度为4 ℃的冰箱中保存解冻,进行生理生化指标的测定。
1.3 生理指标测定及方法
相对电导率测定参照蔡永萍[15]、高风俊[16]的方法,处理后的枝条去除分节部位,剪去节间部位,将样品剪成0.1 cm的小段并混合,取0.1 g放入锥形瓶,加入蒸馏水50 mL,室温浸泡12 h;用DDS-12A型电导仪测定蒸馏水电导率(SCK),测定各样品浸提液的电导率(S1);随后所有样品置于沸水浴处理30 min,恢复到室温;测定煮沸后的电导率(S2),以(1)的公式计算其相对电导率(REC)。各材料每处理各设3次重复,用相对电导率结合Logistic方程计算每份材料的半致死温度(LT50)。
采用MDA、TP、PRO、POD、SOD活性测定试剂盒进行测定,参照试剂盒说明书进行提取和测定操作,根据测定的吸光值计算不同指标的含量,各指标设定3次重复。
1.4 数据处理与分析
相对电导率(REC)按以下公式进行计算,然后拟合Logistic方程,计算各材料的半致死温度(LT50)[23];
相对电导率(%)=S1/S2×100%
①
②
式②中:y为电导率;K为曲线渐近线,即为最大电解质渗出率;α、b均为方程参数;求曲线的拐点(lna/b),即为低温半致死温度(LT50)。利用Excel 2007对其它数据进行处理和方差分析。
利用隶属函数值对实验材料抗寒性进行综合评价。其抗寒性综合评价隶属函数值的计算公式如下,与抗寒性呈正相关的指标的隶属函数值用公式③计算,与抗寒性呈负相关的指标的隶属函数值用公式④计算,具体如下:
各指标与抗性成正相关,则
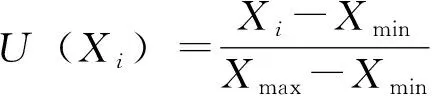
各指标与抗性成负相关,则
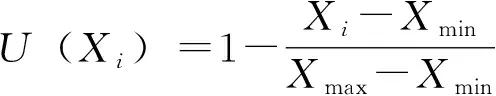
式中,U(Xi)为隶属函数的大小,Xi为各指标的当测值;Xmax为指标最大值;Xmin为指标最小值。
2 结果与分析
2.1 低温胁迫对不同株系电导率的影响
在不同低温的胁迫下,6个文冠果株系的电导率呈现不同的变化(图1),‘株系2’‘G 13’‘BD特异’‘G 27’‘建2’这5个株系枝条相对电导率值随着温度的降低呈现先升高后降低的趋势,株系‘BD 12’随着温度的升高呈现先降低后升高的趋势,在-20 ℃达到最大,为36.24%,温度从4 ℃降到-10 ℃时相对电导率值增加速度较慢;‘株系2’在-10 ℃相对电导率达到最大,为60.92%;温度从-10 ℃降到-20 ℃时‘G 13’‘BD 12’‘G 27’‘建2’这4个株系相对电导率大幅增加,均在-20 ℃达到最大值,分别为59.69%、36.24%、32.70%、31.58%;温度从-20 ℃降到-30 ℃时6个株系组织细胞全部死亡,组织液完全渗出,相对电导率值均呈现下降的趋势,最高电导率与4 ℃差值的大小从大到小依次为‘G 13’>‘G 27’>‘BD 12’>‘BD特异’>‘建2’>‘株系2’。
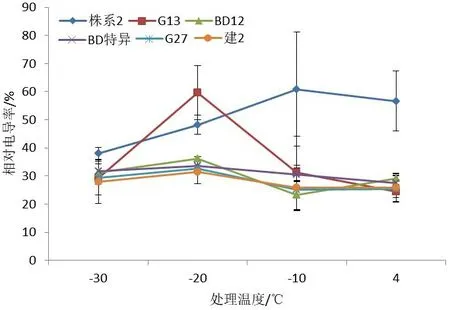
图1 不同株系低温处理下相对电导率变化Fig.1 Changes of relative conductivities of branches from different strains under low temperature stress
2.2 各株系半致死温度(LT50)
在抗寒性的研究过程中,半致死温度可以在一定程度上衡量植物的抗寒性[17-18]。6个文冠果株系的相对电导率用Logistic方程拟合后,所得的半致死温度见表1。

表1 不同株系文冠果低温处理下的半致死温度(LT50)Tab.1 LT50 of branches from different material under low temperature treatments
所有材料的Logistic方程拟合度均达到了0.5以上,可以显示6个株系的半致死温度,其从高到低依次为‘株系2’>‘G 13’>‘建2’>‘BD 12’>‘G 27’>‘BD特异’,其中‘株系2’‘G13’‘建2’的半致死温度在-10 ℃以上,其余3个株系的半致死温度在-10 ℃以下,‘株系2’的半致死温度最高为-2.64 ℃,‘BD特异’的半致死温度最低为-13.27 ℃。结合相对电导率初步分析,‘BD特异’的抗寒性在6个株系中最强。
2.3 低温胁迫对不同株系丙二醛(MDA)的影响
在植物受到低温胁迫时细胞内的代谢平衡会遭到破坏,导致对细胞膜结构和膜上相关大分子有害的活性氧和自由基大量积累,丙二醛是膜脂过氧化作用的最终产物,大量丙二醛与蛋白质结合将会引起膜蛋白变性,最终导致生物膜严重损伤[19],组织内丙二醛的含量越高,组织抗寒性越差,所以丙二醛的含量是抗寒性评价的重要指标[17,20]。
如图2所示,6个文冠果株系在不同梯度低温的胁迫下,枝条内的丙二醛含量均随着温度的下降逐渐上升。在达到半致死温度之前,枝条内丙二醛的含量上升较缓慢,当低于半致死温度之后,枝条丙二醛的含量随着温度的下降而大幅增加,‘BD特异’和‘建2’低温达到-10 ℃至-15 ℃之间,枝条的丙二醛合量几乎不再变化;‘BD特异’的枝条丙二醛含量在-10 ℃达到最大,为1.847 μmol/g;‘G 13’‘BD 12’‘G 27’‘建2’‘株系2’的枝条丙二醛含量在-20℃达到最大,分别为1.300、1.217、1.473、1.308、1.543 μmol/g。根据丙二醛最大含量与4 ℃差值排序得到从大到小依次为‘G 13’>‘株系2’>‘G27’>‘BD12’>‘建2’>‘BD特异’。‘BD特异’各温度下丙二醛含量最高且与其他株系差异显著,判断‘BD特异’‘建2’这2个株系的抗寒性相较其他株系较强。
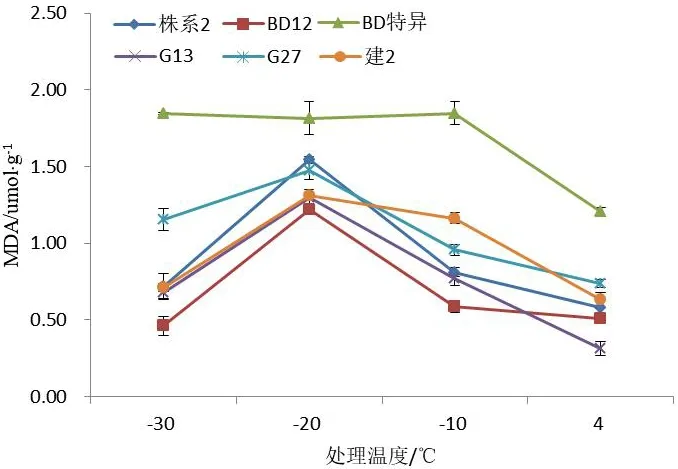
图2 不同株系文冠果低温处理下的丙二醛含量变化Fig.2 Changes of alondialdehyde content of branches from different strains under low temperature stress
2.4 低温胁迫对不同株系可溶性蛋白(TP)的影响
可溶性蛋白是亲水胶体,可在低温胁迫下降低原生质因结冰而导致的对细胞的伤害,保持细胞内水分。因此,低温胁迫下植物体内可溶性蛋白质含量也是衡量植物抗寒性的重要指标[17,21]。如图3所示,在不同低温的胁迫下,6个文冠果株系的可溶性蛋白呈现不同的变化趋势,‘BD特异’‘株系2’‘G 27’‘BD 12’这4个株系枝条可溶性蛋白值随着温度的降低呈现先升高后降低的趋势,其中‘株系2’‘BD特异’这两个株系枝条在-10 ℃达到最大分别为3.920、4.137 g/L;‘G27’‘BD 12’这2个株系枝条在-30 ℃达到最大,分别为3.386、3.410 g/L;‘建2’‘G 13’这2个株系可溶性蛋白值随着温度的降低呈现先降低后升高的趋势,在-30 ℃达到最大,分别为3.873 g/L。6个株系可溶性蛋白最大含量与4 ℃差值的大小从大到小依次为:‘BD特异’>‘G 13’>‘BD 12’>‘BD 12’>‘G 27’>‘株系2’>‘建2’,可初步推断‘BD特异’受到低温胁迫时可溶性蛋白含量上升最快,抗寒性最强。
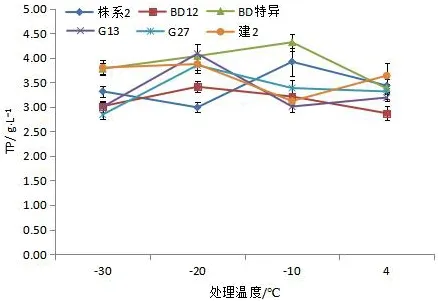
图3 不同株系文冠果的低温处理下可溶性蛋白变化Fig.3 Changes of soluble protein content of branches from different strains under low temperature stress
2.5 低温胁迫对不同株系脯氨酸(PRO)含量的影响
脯氨酸在植物的抗逆生理代谢过程中,是提供代谢原料和能量的一种重要内含物质,可以抵抗渗透胁迫,脯氮酸在枝条中含量的高低及变化趋势是枝条抗寒性的一种主要体现[20]。如图4所示,在不同低温的胁迫下,6个文冠果株系的脯氨酸含量呈现不同的变化,‘BD特异’‘株系2’‘建2’这3个株系枝条脯氨酸含量值随着温度的降低呈现先降低后升高的趋势,‘BD 12’‘G 27’‘G 13’这3个株系枝条脯氨酸含量随着温度的降低呈现先升高后降低的趋势。‘BD特异’‘株系2’‘G 13’‘BD 12’‘G 27’这5个株系枝条中的脯氮酸中含量在-30 ℃达到最大,分别为1.847、1.687、1.857、1.437、1.993 ug/mg,‘建2’枝条中的脯氮酸中含量在-10 ℃达到最大为1.483 ug/mg。
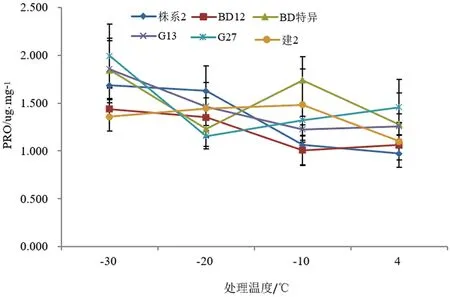
图4 不同株系文冠果的低温处理下脯氨酸含量变化Fig.4 Changes of proline content of branches from different strains under low temperature stress
2.6 低温胁迫下不同株系超氧化物歧化酶(SOD)活性的变化
植物在受到低温胁迫时超氧化物歧化酶能够调节细胞膜的透性,有效地清除活性超氧自由基,保护细胞不被伤害并维持正常的代谢,保护细胞膜的结构和功能稳定[21-22]。在不同低温胁迫下,6个不同株系的超氧化物歧化酶活性呈现不同的变化趋势(图5),其中‘BD特异’‘株系2’‘G 13’‘G 27’的超氧化物歧化酶活性值随着温度的降低呈现先升高后降低的趋势,‘株系2’‘BD 12’枝条的超氧化物歧化酶活性随着温度的降低呈现先升高后降低的趋势。‘BD特异’‘G 13’‘G 27’‘建2’的超氧化物歧化酶在-10 ℃达到最大值,分别为25.777、17.013、19.507、16.330 U/mg,‘BD 12’枝条在-30 ℃达到最大为12.513 U/mg,‘株系2’枝条在-20 ℃达到最大为21.873 U/mg。根据最高SOD含量与4 ℃差值的大小从大到小依次为‘株系2’‘BD特异’>‘G 27’>‘G 13’>‘建2’>‘BD 12’,初步推断‘BD特异’‘G 27’‘株系2’的文冠果枝条抗寒性较强。
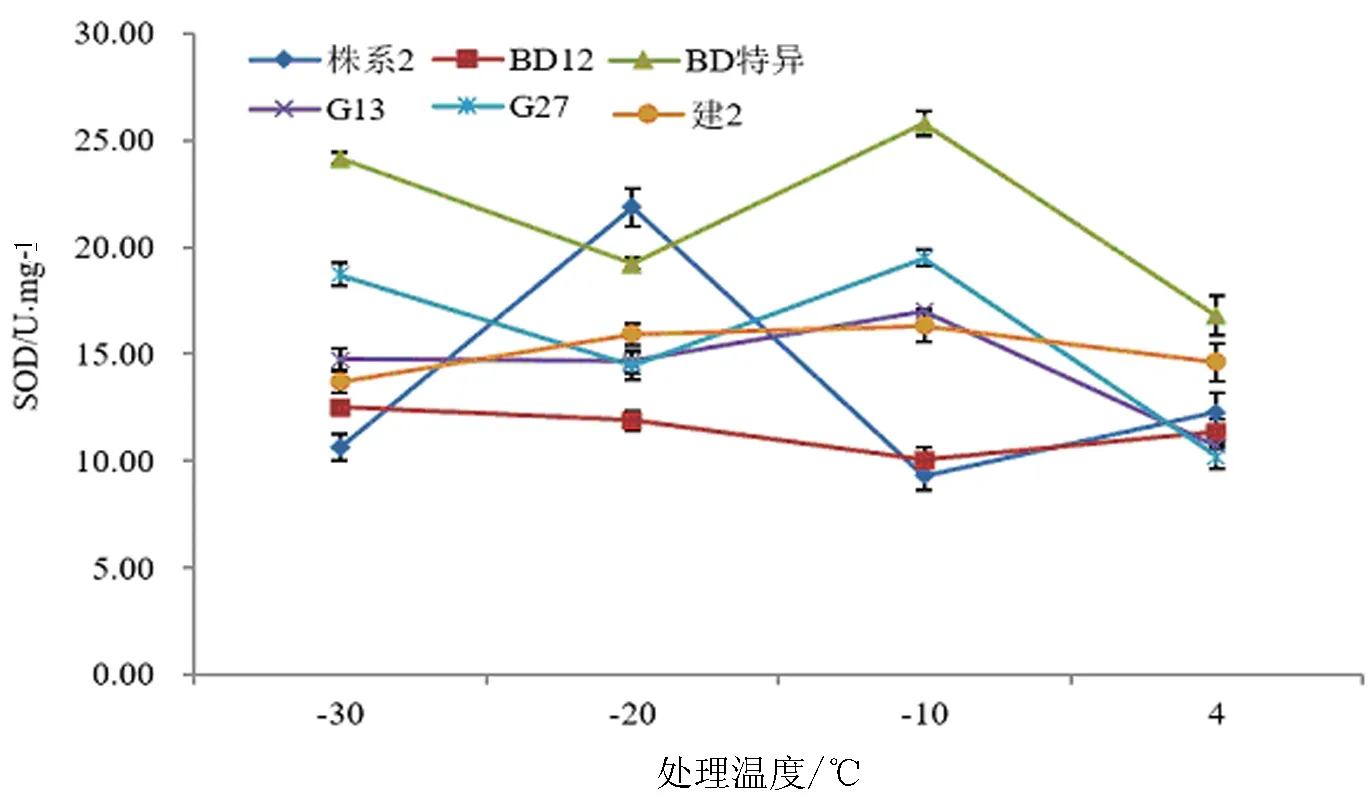
图5 不同株系文冠果低温处理下的超氧化物歧化酶活性变化Fig.5 Changes of content of superoxide dismutase of branches from different strains under low temperature stress
2.7 低温胁迫下不同株系过氧化物酶(POD)活性变化趋势
过氧化物酶清除细胞内氧化还原产生的H2O2等过氧化物,在抗氧化防御系统中有非常重要的位置[21-22]。
如图6所示,在不同低温的胁迫下,过氧化物酶活性呈现不同的变化,6个株系枝条的过氧化物酶活性随着温度的降低呈现先降低后升高的趋势。‘株系2’‘BD 12’‘G 13’‘建2’在受到低温胁迫过程中整体活力与4 ℃相比呈现下降趋势,都受到了低温胁迫的影响。‘BD特异’‘G 27’枝条在受到低温胁迫时,过氧化物酶活性最大值相对4 ℃分别增加了0.48、1.45,抗寒性较强。6个株系枝条过氧化物酶活性的最高值与4 ℃的差值从大到小依次是‘G 27’>‘BD特异’>‘BD 12’>‘株系2’>‘建2’>‘G 13’,初步推断‘BD特异’‘G 27’抗寒性较强。
2.8 不同材料各单项抗寒指标的隶属度值与综合抗寒性的关系
植物的抗寒能力是一个综合的表现,不是靠单一的指标就能得出来的结果。因此采用模糊数学中隶属函数法来对6个文冠果株系的抗寒性的强弱进行综合评价,综合评价排序越靠前,抗寒性越强。评价指标中电导率、可溶性蛋白含量、脯氨酸含量、超氧化物歧化酶活性、过氧化物酶与抗寒性成正相关,丙二醛与抗寒性成负相关[20-25]。从表2可以看出,6个文冠果株系相对隶属函数值的大小顺序为:‘BD特异’>‘株系2’>‘G 27’>‘建2’>‘G 13’>‘BD 12’,结合半致死温度及各项生理指标、形态指标分析结果,筛选出‘BD特异’这个株系在物候期、形态学指标、抗寒性各个方面均表现很好,综合评价生长适应性最好,可以作为优良株系进行繁殖推广。
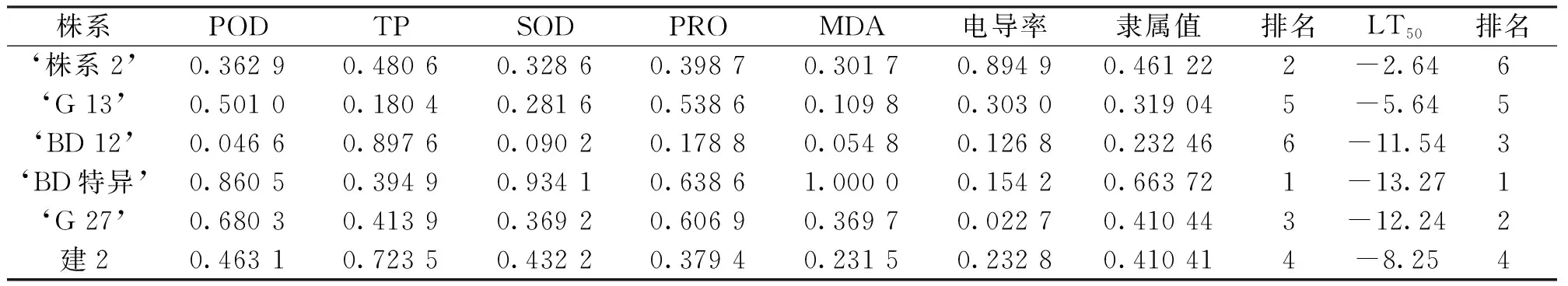
表2 不同株系文冠果的抗寒性指标隶属值Tab.2 Subordination values of cold resistance indexes of different materials
3 讨论与结论
3.1 讨论
在鉴定植物抗寒性时,相对电导率是一种较为直观的表示植物在低温胁迫下细胞质膜透性变化的方法,细胞膜系统的稳定性与植物抗寒性存在密切关系,其直接反映植物受害程度[26-28],电解质外渗率结合Logistic曲线方程推导半致死低温能准确地反映植物耐受的低温极限[23-24]。多数研究认为电导率与植物抗寒性呈现正相关关系[20-21]。通过测定分析发现:在不同低温的胁迫下,6个文冠果株系的电导率呈现不同的变化,6个株系的半致死温度从高到低依次为‘株系2’>‘G 13’>‘建2’>‘BD 12’>‘G 27’>‘BD特异’,其中‘株系2’的半致死温度最高为-2.64 ℃,‘BD特异’的半致死温度最低为-13.27 ℃,可初步推断‘BD特异’的抗寒性最强。
活性氧超过正常水平时,可导致膜脂过氧化、蛋白质变性等一系列不良反应。丙二醛是膜脂过氧化作用的产物,常作为反映生物膜受伤害程度的指标[25-29]。6个文冠果株系在不同梯度低温的胁迫下,枝条内的丙二醛含量均随着温度的下降逐渐上升。在达到半致死温度之前,枝条内丙二醛的含量上升较缓慢,当低于半致死温度之后,枝条丙二醛的含量随着温度的下降而大幅增加,这与马新等人对文冠果抗寒性的研究结果一致[5,13]。
SOD和POD等保护酶系统在低温条件下积极发挥作用,减轻由膜脂过氧化所引起的膜伤害,SOD能清除超氧阴离子自由基,而POD能清除产生的过氧化氢[22-23]。可溶性蛋白可以调节细胞的渗透势,同时也使细胞的水合度增大,保水能力增强,降低细胞结冰的温度[20-31]。多数研究认为低温胁迫下可溶性蛋白、超氧化物歧化酶、过氧化物酶与植物抗寒性正相关[32-34],‘BD特异’各温度下丙二醛、可溶性蛋白、超氧化物歧化酶、过氧化氢酶含量均随着温度的降低出现先升高后降低的趋势,这与马新等[5]对文冠果抗寒性研究结果一致。
隶属函数法作为能够综合所有指标对植物抗寒性进行综合评价的一种方法,已经广泛应用于长柄扁桃(AmygdaluspedunculataL.)、木瓜(ChaenomelessinensisKoehne)等植物的抗寒性评价[34-36]。文冠果株系相对隶属函数值的大小顺序为:‘BD特异’>‘株系2’>‘G 27’>‘建2’>‘G 13’>‘BD 12’,综合抗寒性生理生化指标、物候期适应性和形态学指标筛选,最终筛选出‘BD特异’这个株系,其在开花结实能力,抗寒性各个方面均表现很好,综合评价生长适应性最好,可以作为优良株系进行繁殖推广。
3.2 结论
通过前期的形态学指标分析结果,结合抗寒性研究结果得出:(1)6个文冠果株系的电导率呈现不同的变化,半致死温度从高到低依次为‘株系2’>‘G 13’>‘建2’>‘BD 12’>‘G 27’>‘BD特异’,其中‘株系2’的半致死温度最高,为-2.64 ℃,‘BD特异’的半致死温度最低,为-13.27℃,可初步推断‘BD特异’的抗寒性最强。(2)6个文冠果株系枝条内的丙二醛含量均随着温度的下降逐渐上升,在达到半致死温度之前,枝条内丙二醛的含量上升较缓慢;当低于半致死温度之后,枝条中丙二醛的含量随着温度的下降而大幅增加。(3)‘BD特异’在各温度下其丙二醛、可溶性蛋白、超氧化物歧化酶、过氧化氢酶含量均随着温度的降低呈现先升高后降低的趋势。(4)文冠果6个株系相对隶属函数值的大小顺序为:‘BD特异’>‘株系2’>‘G 27’>‘建2’>‘G 13’>‘BD 12’。最终筛选出‘BD特异’这个株系在各个方面均表现很好,综合评价生长适应性最好,既可作为油料树种备选株系,又可作为生态树种备选株系,在新疆石河子地区进行进一步推广种植。
