广双瘤蚜茧蜂对棉蚜脂质合成通路的影响
2021-02-23高雪珂常国凤崔金杰陈利珍
薛 汇,高雪珂,常国凤,崔金杰*,陈利珍
(1. 华中农业大学植物科学技术学院,武汉 430070;2. 中国农业科学院棉花研究所/棉花生物学国家重点实验室,安阳 455000)
棉蚜Aphis gossypiiGlover是一种广泛分布于世界各国的常见害虫[1]。棉蚜是刺吸式口器,大量棉蚜若虫和成虫集中在棉叶背面和幼茎上吸食寄主植物汁液,造成叶片卷缩畸形,植株矮小,甚至幼苗枯死[2]。此外,棉蚜分泌的蜜露会造成棉花叶片的煤污病,影响光合作用,棉蚜还能传播多种植物病毒,降低作物的产量和品质[3],严重危害棉花的生产[4]。蚜茧蜂是蚜虫的寄生性天敌昆虫的主要类群,可有效控制棉蚜的发生[5],其中,广双瘤蚜茧蜂是棉花蚜茧蜂的优势种群[6]。
寄生蜂和寄主之间有着复杂的制约关系,寄生蜂能够通过化学物质或者气味准确定位寄主位置[7]。寄生形式分为内寄生和外寄生两种,寄生蜂幼虫取食寄主体表或取食寄主体内组织,通常情况下,寄生蜂幼虫前期主要取食寄主血淋巴,后期直接取食寄主脂肪体[8,9]。幼虫随着寄主的生长发育,并最终导致寄主死亡[10]。动物以脂肪的形式储存能量的能力是生存和繁殖所必需的[11-13],绝大多数的寄生蜂缺乏脂质合成的能力[14]。
寄生蜂为满足自身对脂质的需求,对寄主的生理生化进行调节,如生殖、代谢、生物合成等,会造成寄主体内脂质大量积累,包括全身脂质含量的增加[15],脂肪体三酰基甘油代谢的增强[8,16]以及血淋巴中游离脂肪酸水平的升高等[8,9]。1974年发现寄生蜂调节寄主脂肪酸组成[17],2004年报道寄生蜂体内的脂肪酸构成高度相似于蚜虫体内的脂肪酸[18],Visser和 Ellers[19]推断,寄生蜂缺乏与脂肪酸合成相关的酶系,通过调节寄主脂肪酸合成来完成自身的生物合成代谢。寄生蜂还可以通过调节寄主体内激素滴度来调节寄主的摄食习惯和生长[7,20],目前已发现寄生物能够引起宿主内、外寄生蜂体内氨基酸、蛋白质、丙酮酸和碳水化合物数量的变化[21-25]。同时通过破坏蚜虫的免疫系统和改变蚜虫的营养流向来达到改造的目的,使幼虫更好地发育[7,26]。
目前广双瘤蚜茧蜂对棉蚜脂质合成调控的报道较少,本试验采用荧光定量PCR技术,检测了被寄生3 d棉蚜和未被寄生棉蚜脂肪酸合成通路、甘油酯合成通路、糖酵解以及氨基酸代谢通路中的关键基因,明确被寄生棉蚜脂质合成相关基因的表达变化,对更好地了解广双瘤蚜茧蜂影响棉蚜脂质合成机制及棉蚜的生物防控具有重要意义。
1 材料与方法
1.1 供试植株
试验所用的棉花品种为中棉所49,由中国农业科学院棉花研究所棉花种质资源中期库提供,棉花种植于人工气候室(26±1)℃,光周期14L:10D,RH(75±5)%。
1.2 供试昆虫
所用棉蚜为实验室继代饲养的孤雌繁殖的单个种群棉蚜,饲养条件为(26±1)℃,RH(65±5)%,光周期14L:10D。广双瘤蚜茧蜂采自中国农业科学院棉花研究所试验农场(安阳县),并于(26±1)℃,RH(65±5)%,光周期14L:10D人工智能光照培养箱内饲养。
1.3 寄生棉蚜的获取
将2龄若蚜接入含有棉花叶片的培养皿中,然后接入寄生蜂,观察寄生蜂的寄生行为,寄生蜂寄生棉蚜后立即将被寄生棉蚜转入新鲜棉花叶片上饲养3 d,光照培养箱饲养条件为 (26±1)℃,RH(65±5)%,光周期14L:10D。被寄生3 d的棉蚜进行解剖,取出寄生蜂幼虫,收集被寄生的棉蚜样品,同时未被寄生的棉蚜也进行相同的解剖操作。试验组和对照组各设置3个生物学重复,每个生物学重复20头棉蚜。
1.4 RNA的提取及cDNA的合成
使用 TRIZOL试剂(Invitrogen,Carlsbad,CA,USA)从被寄生的棉蚜和未被寄生的蚜虫中提取总RNA,操作方法参照厂商说明。使用1.5%琼脂糖凝胶电泳检测RNA的完整性,并使用Nano-Drop 2000(Thermo Scientific,Wilmington,DE,USA)检测RNA的浓度及纯度。cDNA的合成利用500 ng上述RNA通过PrimeScriptfi RT Reagent Kit(Takara,Dalian,China)参照试剂盒的步骤进行合成。
1.5 实时荧光定量技术的基因表达量验证
实时荧光定量检测使用Mastercycler ep realplex(Eppendorf,Hamburg,Germany)进行,基因特异性引物利用BEACON DESIGNER 7.6(PREMIER Biosoft International,CA,USA)设计并通过上海生工生物工程公司合成,引物序列见表1,内参基因选用二甲基腺苷转移酶dimethyladenosine transferase(GenBank:KM507111)和肽酰-脯氨酰-顺反式异构酶peptidyl-prolyl cis-trans isomerase(GenBank:KF018924)[27],取两种内参基因表达量的平均值进行数据分析。检测基因表达量的反应程序为:95 ℃,3 min;95 ℃,5 s和60 ℃,30 s,共40个循环。

表1 棉蚜脂质合成相关基因及引物序列Table 1 Gene and primer sequence related to lipid synthesis of Aphis gossypii
1.6 数据统计与分析
对实时荧光定量PCR数据采用2-ΔΔCt方法[28]进行分析,利用GraphPad Prism 7制作图表,并利用IBM SPSS Statistics 20采用独立样本t检验进行差异显著性分析,P<0.05被认为是差异显著的。“*”:P<0.05;“**”:P<0.01;n.s.:无显著性差异。
2 结果与分析
2.1 被寄生和未被寄生棉蚜糖酵解基因转录水平差异比较
在糖酵解途径中,被寄生棉蚜的1,6-二磷酸果糖醛缩酶基因表达量上调了1.7倍,磷酸甘油酸激酶基因表达量上调了1.6倍。但蔗糖转化为丙酮酸的关键限速酶丙酮酸激酶表达量降低了0.3倍(图1)。

图1 棉蚜被广双瘤蚜茧蜂寄生后糖酵解通路基因的表达变化Fig. 1 Changes of glycolysis pathway gene expression of cotton aphid after parasitized by Binodoxys communis
2.2 被寄生和未被寄生棉蚜脂肪酸合成基因转录水平差异比较
图2揭示了被寄生棉蚜和未被寄生棉蚜脂肪酸合成相关基因的表达情况。棉蚜被广双瘤蚜茧蜂寄生3 d后,除了脂肪酸合成酶1和硬脂酰辅酶A去饱和酶1表达量没有显著差异外,其他基因都显著上调。其中乙酰辅酶A羧化酶基因表达量上调了6.5倍,三磷酸腺苷(ATP) 柠檬酸裂解酶基因表达量上调了2.3倍,脂肪酸合成基因2、3表达量分别上调了6.1和3.3倍,硬脂酰辅酶A去饱和酶2表达量上调了7.8倍。
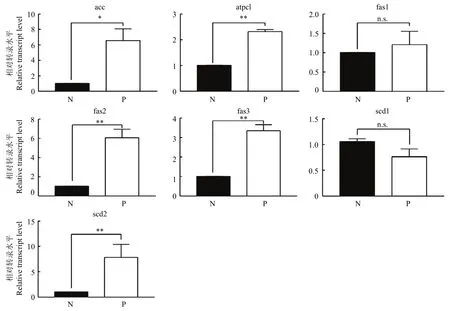
图2 棉蚜被广双瘤蚜茧蜂寄生后脂肪酸合成通路基因的表达变化Fig. 2 Changes of fatty acid synthesis pathway gene expression of cotton aphid after parasitized by Binodoxys communis
2.3 被寄生和未被寄生棉蚜甘油酯合成基因转录水平差异比较
在三酰甘油合成途径中,棉蚜被寄生3 d后,被寄生棉蚜体内1-酰基-sn-甘油-3-磷酸酰基转移酶基因表达量上调2.8倍、二酰甘油O-酰基转移酶基因表达量上调3.4倍、甘油三磷酸酰基转移酶基因表达量上调1.7倍、脂滴表面蛋白基因表达量上调2.6倍、三酰基甘油脂酶1和2基因表达量分别上调1.6和2.5倍(图3)。
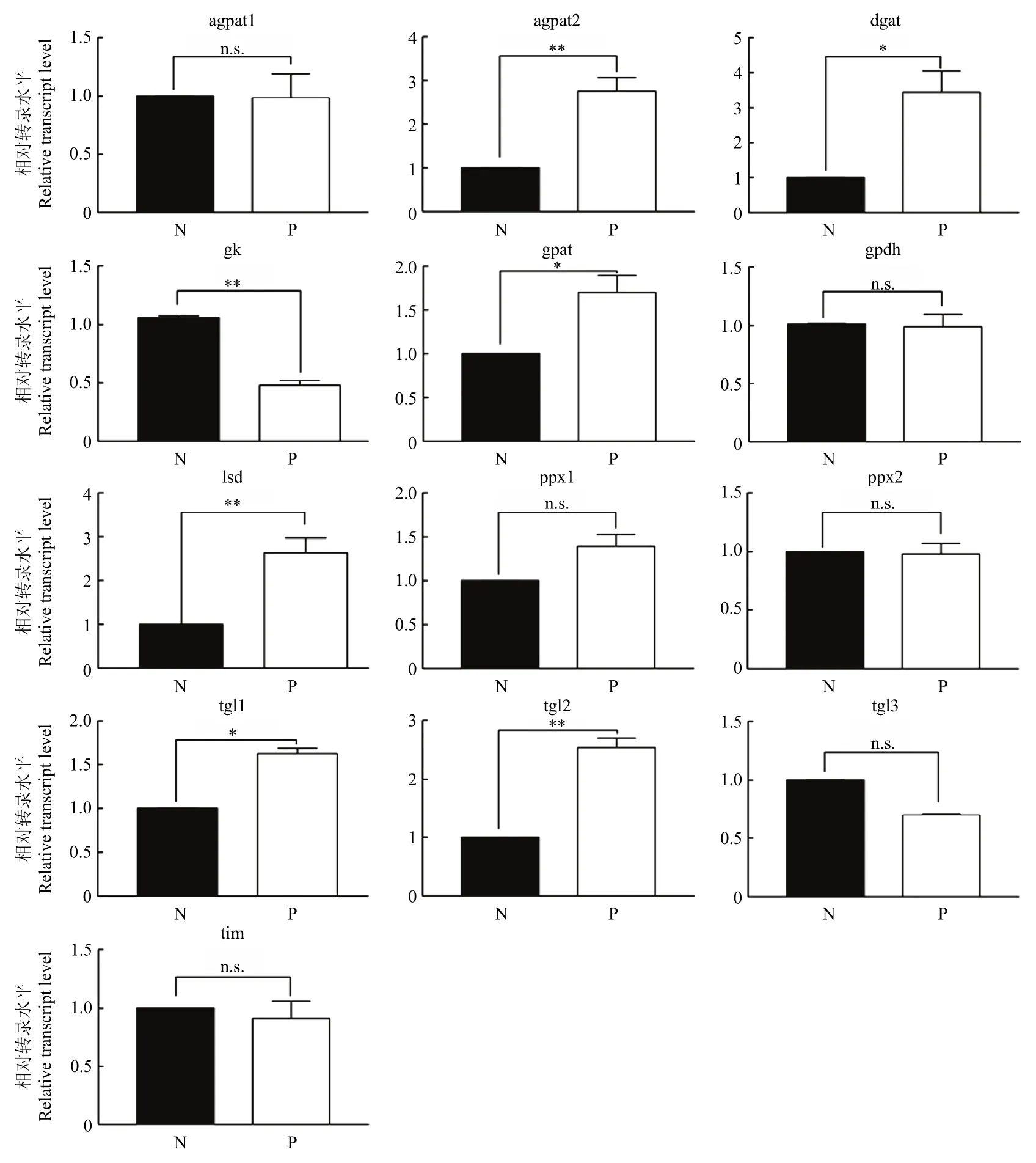
图3 棉蚜被广双瘤蚜茧蜂寄生后甘油酯合成通路基因的表达变化Fig. 3 Changes of glyceride synthesis pathway gene expression of cotton aphid after parasitized by Binodoxys communis
2.4 被寄生和未被寄生棉蚜氨基酸代谢基因转录水平差异比较
氨基酸是棉蚜生长发育所需的氮源[29],我们对氨基酸转化为柠檬酸,然后到乙酰辅酶A通路中的关键基因的表达量进行检测。结果显示被寄生棉蚜中氨基酸转化为乙酰辅酶A通路中柠檬酸合酶基因表达量上调7.1倍,异柠檬酸脱氢酶基因表达量上调9.5倍,线粒体异柠檬酸脱氢酶β亚基基因表达量上调5.7倍,线粒体异柠檬酸脱氢酶γ亚基基因表达量上调4.1倍,线粒体苹果酸脱氢酶基因表达量上调2.8倍。虽然胞质异柠檬酸脱氢酶基因、异柠檬酸脱氢酶α亚基基因、细胞质苹果酸脱氢酶基因表达量在统计意义上变化不具显著性,但其表达量分别上调了5.5倍、1.6倍和2.2倍(图4)。
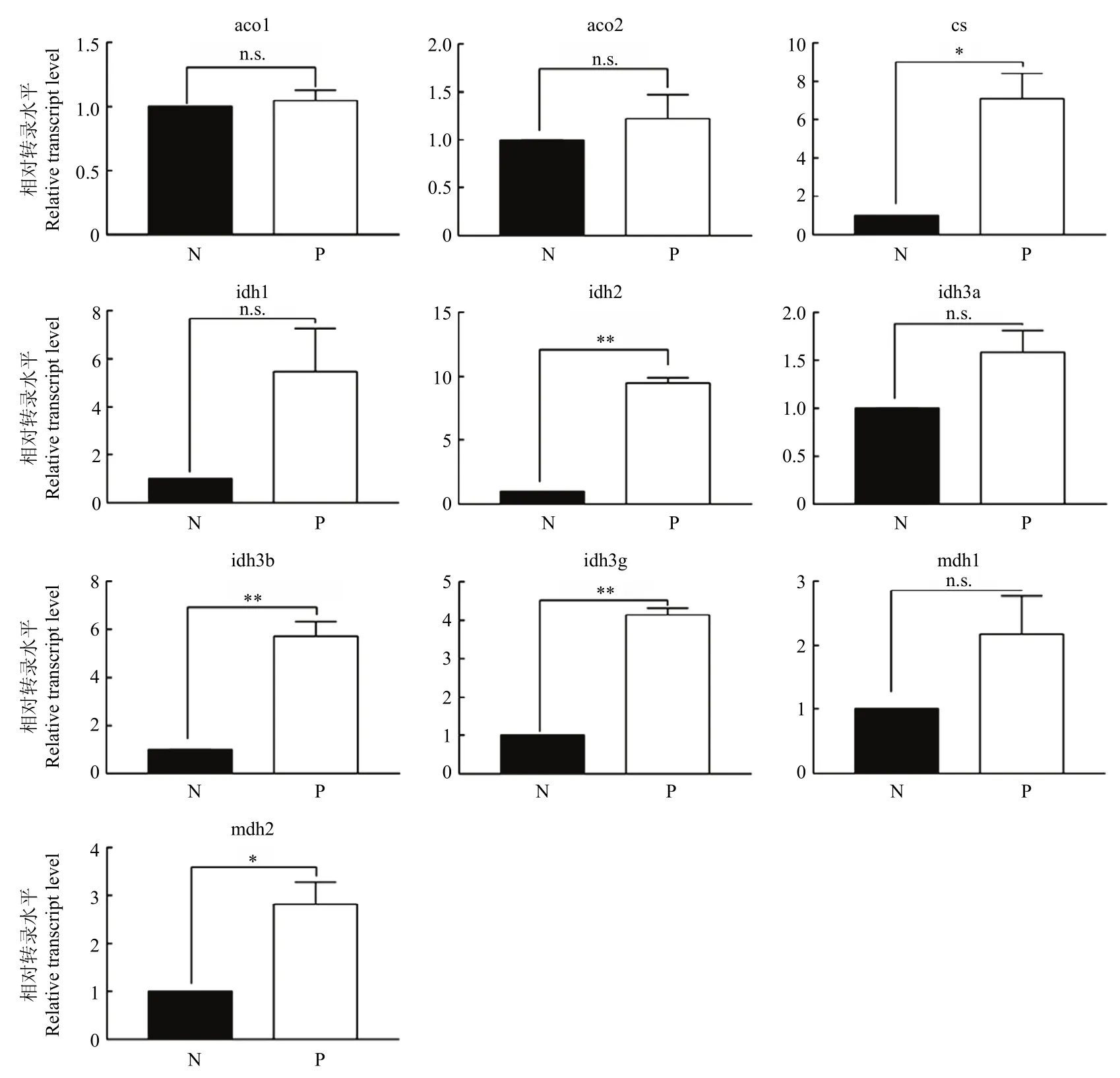
图4 棉蚜被广双瘤蚜茧蜂寄生后氨基酸代谢通路基因的表达变化Fig. 4 Changes of amino acid metabolism pathway gene expression of cotton aphid after parasitized by Binodoxys communis
3 讨论
寄生蜂通常能够引起宿主体内脂质含量的变化[16,30],增加血淋巴中游离的脂肪酸含量,增强脂肪体中三酰甘油的代谢能力[16]。如黑头柔茧蜂通过对宿主脂肪体内的营养物质精细调控来满足自身的营养适应性[31],棉蚜被棉蚜茧蜂寄生后,棉蚜体内游离的脂肪酸显著性增加[32],粘虫被粘虫姬小蜂寄生后,体内脂肪含量明显高于未被寄生的粘虫[16]。寄生蜂引起寄主体内脂肪代谢通路的变化多有报道,但在广双瘤蚜茧蜂对棉蚜脂质通路上的调控作用研究较少,本试验采用荧光定量PCR技术,明确了被广双瘤蚜茧蜂寄生3 d的棉蚜脂质合成通路相关基因的表达变化,揭示了广双瘤蚜茧蜂对棉蚜脂质合成通路的调控。
研究表明,棉蚜茧蜂寄生棉蚜后,被寄生的棉蚜几乎所有的与甘油酯合成相关基因的表达量都明显升高,其中限速酶二脂酰甘油酰基转移酶基因表达量上调了3.2倍,脂滴表面结合蛋白基因表达量上调了5.0倍[27],同时被棉蚜茧蜂寄生的棉蚜体内的甘油酯和甘油磷酯的含量也会增加[33]。与此研究结果一致,本研究中,棉蚜被广双瘤蚜茧蜂寄生3 d后,与甘油三酸酯合成相关的关键基因的表达量多数上调,限速酶二脂酰甘油酰基转移酶上调了3.4倍,脂滴表面结合蛋白基因表达量上调了2.6倍。其中,脂滴能够储存胆固醇和脂酰甘油,是膜形成和维持的重要物质,而且能主动参与脂肪和能量代谢[7]。脂滴表面蛋白基因可以调节脂解,促进或者抑制脂解[34]。结果表明广双瘤蚜茧蜂能够激活棉蚜甘油酯合成通路,促进被寄生棉蚜中甘油酯的合成。
蔗糖是棉蚜生长发育所需的碳源,蔗糖在昆虫体内的代谢途径已比较明确,主要是通过糖酵解过程将蔗糖分解成葡萄糖和果糖参与到脂质合成中来。蔗糖被分解成葡萄糖和果糖,然后转换成果糖-1,6-二磷酸,果糖-1,6-二磷酸被分解成磷酸二羟丙酮(DHAP)和甘油醛-3-磷酸(GAP),DHAP和GAP有两种代谢途径,一种途径中DHAP和GAP被转化成三酰甘油,另一种途径就是DHAP和GAP被代谢为丙酮酸,丙酮酸能够转化成乙酰辅酶A,乙酰辅酶A是合成脂肪酸的前提[35]。Gao等[36]在2020年的研究中指出,被棉蚜茧蜂寄生后的棉蚜糖酵解通路中关键基因甘油醛-3-磷酸脱氢酶、葡萄糖-6-磷酸异构酶表达量显著上调。本研究中,被广双瘤蚜茧蜂寄生3 d后的棉蚜体内1,6-二磷酸果糖醛缩酶基因表达量上调了1.7倍,磷酸甘油酸激酶基因表达量上调了1.6倍。但丙酮酸激酶表达量降低了,虽然没达到显著水平,丙酮酸激酶是蔗糖转化为丙酮酸的关键限速酶,丙酮酸能够转化为合成脂肪酸的原料——乙酰辅酶A。我们推测虽然糖酵解途径产生丙酮酸的能力减弱了,合成脂肪酸的C骨架可能来自游离的氨基酸。同时我们对氨基酸转化为柠檬酸然后到乙酰辅酶A途径中的相关基因的表达量进行了检测,结果显示被寄生棉蚜中氨基酸转化为乙酰辅酶A通路中的基因表达量显著上调,也暗示在被寄生棉蚜中氨基酸可能被用于作为合成脂肪酸的原料。
脂肪酸是能量的主要来源,也是细胞膜的重要组成部分[37]。Gao等[32]通过检测被棉蚜茧蜂寄生后棉蚜的脂肪酸合成通路基因表达情况,发现乙酰辅酶A羧化酶基因表达量上调了5.0倍,脂肪酸合成基因1和3分别上调6.9倍和4.1倍。本研究中,棉蚜被广双瘤蚜茧蜂寄生3 d后,几乎所有与脂肪酸合成相关基因的表达量都显著上调。寄生蜂由于缺乏与脂肪酸合成相关的酶系统,调控寄主脂肪酸合成通路来完成自身生长发育的需要[38]。
长期以来,寄生蜂和寄主的互作关系一直是研究的热点。棉蚜是重要的农业害虫,不仅为害棉花,对一些园艺作物的为害也日益加重,而蚜茧蜂作为棉蚜主要寄生蜂种类,具有较好的生防价值。本研究就广双瘤蚜茧蜂寄生棉蚜后,对棉蚜脂质合成通路中的脂肪酸合成途径、氨基酸代谢途径、甘油酸三酯合成途径以及糖酵解途径的影响做了初步研究,明确了被寄生棉蚜脂质合成通路相关基因的表达变化,对于揭示广双瘤蚜茧蜂与棉蚜之间的互作以及寄生蜂与寄主昆虫的协同进化有重要意义,同时对开发新型棉蚜防控技术具有重要的理论实践价值。
