云南派姬小蜂对其寄主及寄主植物相关化学线索的行为反应
2021-02-23程曼曼马梦然王树香崔建州李继泉张国伟
程曼曼,王 淼,马梦然,王树香,崔建州,3,李继泉,3**,张国伟
(1. 河北农业大学林学院,保定 071000;2. 河北农业大学生命学院,保定 071000;3. 河北省林木种质资源与森林保护重点实验室,保定 071000;4. 河北省洪崖山国有林场管理局七里亭示范场,易县 074200)
寄生蜂的寄生效率在一定程度上取决于其自身的寄主栖境定位和寄主定位能力。雌蜂要成功完成寄生须经历5个阶段:寄主栖境定位、寄主发现、寄主接受、寄主适应和寄主调节。在每个阶段,寄生蜂都要对来自与寄主植物和寄主相关的化学线索,做出正确的反应[1,2]。证据表明,寄生蜂首先利用化学线索找到寄主栖境,然后再搜寻寄主[2]。寄主植物挥发物常作为互益素,用于寄主栖境定位(长距离搜索)[3-5];来自寄主昆虫及其相关物质(虫粪、老熟幼虫、茧和蛹体表角质层)的化学线索作为利它素,用于寄主定位(短距离搜索)[3,6-10]。寄主植物挥发物虽然易检测,但据此判断寄主是否存在并不可靠。与寄主昆虫相关的化学刺激非常可靠,但通常在远距离难以检测到。因此,虫害诱导植物产生的挥发物将这种气味的可检测性和可靠性结合起来,对寄主栖境远程定位起着关键作用[11-13]。
微红梢斑螟Dioryctria rubellaHampson是我国重要的钻蛀性害虫,主要以幼虫蛀食多种松科植物的枝梢和球果[14]。越冬代幼虫,在枝条内发育到3龄幼虫时,转入当年生枝梢或球果中继续为害[15]。为害初期,幼虫虫粪排到松枝和球果外,后期则留在蛀道内。老熟幼虫在化蛹前先咬出羽化孔,然后回到蛹室内结茧,2~3 d后化蛹。此时,枝梢枯萎,球果局部坏死,严重影响木材的利用价值及种子产量[14,16]。因其幼虫生活隐蔽,化学防治效果不理想[14],利用天敌昆虫对钻蛀类害虫进行生物防治,日益受到重视[17]。云南派姬小蜂Pediobius yunnanensisLiao是微红梢斑螟的蛹寄生蜂,寄生率较高[18]。马梦然等[18-20]对其寄生生物学特性、发育起始温度和有效积温以及环境条件对其生长发育的影响等方面进行了系统研究,但该寄生蜂对寄主植物和寄主相关化学线索的行为反应还不清楚。因此,本文对此方面的内容进行研究,为揭示该寄生蜂寄主搜寻行为的化学调控机制以及微红梢斑螟的生物防治提供理论依据。
1 材料与方法
1.1 供试植物
在河北农业大学标本园内,选取8~12年生的油松为试验材料,该标本园的油松生长健壮且未受到微红梢斑螟为害。
1.2 供试昆虫
4月末,在秦皇岛北戴河区道路两侧的被害油松上,采集带有云南派姬小蜂幼虫的松枝和球果,带回实验室后,将寄生蜂的幼虫置于试管(2 cm×15 cm)中,在25 ℃、RH 70%和光周期14L:10D的培养箱内培养,羽化后的寄生蜂作为种蜂,转入另一试管,待交配后,供给微红梢斑螟蛹,在上述相同的条件下培养,以建立室内饲养种群。待成蜂羽化后,饲喂15%的蜂蜜水,1~3日龄的雌蜂用于生测试验。5月初—8月底,于河北省易县七里亭示范场的油松林中,采集带有微红梢斑螟幼虫和蛹的球果和松梢,带回实验室后剥出蛹和幼虫,低日龄的蛹(淡黄色或淡红色)放入试管中,7 ℃下贮存备用,幼虫放入养虫笼内(40 cm×40 cm×40 cm),用新鲜的油松嫩枝和球果饲养。其中,3龄幼虫用于油松处理,其他幼虫培养至化蛹后,低温贮存备用。
1.3 寄生蜂对寄主植物和幼虫虫粪挥发物的趋性行为
利用“Y”型嗅觉仪,测定了寄生蜂对不同处理球果和松枝以及幼虫虫粪挥发物的趋性行为。
1.3.1 寄主植物气味源 含蛹球果:在每株健康的油松上,选取8个带1年生球果的枝条,用尼龙网将微红梢斑螟的3龄幼虫和球果罩住,球果被害10 d后去除尼龙网,每天定期检查球果,出现羽化孔后,每天从羽化孔观察老熟幼虫的化蛹情况,当虫体完全被茧包裹时,表明已化蛹,以此时间点作为化蛹起始时间。当蛹为 2~5日龄时,剪下含蛹球果并作为气味源。含空蛹壳的球果:处理方法同含蛹球果,以微红梢斑螟成虫羽化3 d后的球果作为气味源。幼虫为害的球果:处理方法同含蛹球果,以3龄幼虫为害3 d后的球果(除去球果外虫粪)作为气味源。健康球果:以健康植株上的球果作为气味源。含蛹松枝:在每株油松上,选取8个粗壮的1年生健康枝条,用尼龙网将枝条罩住(不包含球果),然后放入1头3龄幼虫,让其取食为害。因1个松枝不能满足3龄幼虫发育至蛹对食物的需求,当幼虫将松枝木质部吃空后,将其转移到相邻的1年生松枝上,松枝上出现羽化孔后,每天观察化蛹情况,当蛹为2~5日龄时,剪下含蛹松枝并作为气味源,切口包以湿润脱脂棉后,外层再用铝箔包裹,以防止松枝气味从切口处释放。幼虫为害的松枝:方法同含蛹松枝,以3龄幼虫为害3 d后的松枝(除去松枝外虫粪)作为气味源。含蛹球果的系统枝:方法同含蛹球果,以去除含蛹球果后的1年生松枝为气味源。含蛹松枝的系统枝:方法同含蛹松枝,以含蛹松枝同节上的1年生枝条为气味源。健康松枝:在健康植株上,剪取1年生松枝作为气味源。
1.3.2 幼虫虫粪气味源 3龄幼虫虫粪:3龄幼虫取食球果3 d后,收集球果外的虫粪,以10 g虫粪作为气味源。含蛹球果内的虫粪:剥开含蛹(2~5日龄)球果,收集蛀道内的虫粪(10 g)作为气味源。
1.3.3 生测方法 参照王薇等[21]的方法进行测定。Y形嗅觉仪主臂长8 cm,两个测试臂(外径2.5 cm)长10 cm、夹角80°。两个测试臂分别与装有待测气味源和空气(对照)的广口瓶(5 L,用于松枝测定;250 mL,用于球果测定)或层析柱(长25 cm,内径1 cm,用于虫粪测定)相连,以气泵为动力将空气依次通过活性炭和蒸馏水,过滤、加湿后的洁净空气再经三通分为两股流速相同的气流(100 mL/min),分别流入装有待测气味源和空气的广口瓶或层析柱后进入2个测试臂,最后经主臂进入大气中。嗅觉仪放入用金属框架和白布制成的围帐中,在其正上方放置一个20 W的荧光灯管,并使两测试臂的光强保持一致,环境温度(26±1)℃。用硅胶管将气味源和嗅觉仪连接好后开启气泵,将 1只雌蜂放入主臂后,观察并记录其选择情况。在5 min 内未作出选择的雌蜂,不作为试验数据进行分析。每个处理重复8~10次后,更换新鲜气味源。每头雌蜂只使用1次。
1.4 寄生蜂对老熟幼虫爬行痕迹的行为反应
由于老熟幼虫在化蛹前会从球果或枝条内部向外部取食,直至在球果或枝条表面咬出羽化孔,然后回到蛹室内准备化蛹。老熟幼虫在羽化孔和蛹室间爬行时留下的痕迹,是否作为该寄生蜂寄主定位的化学线索,还不清楚。故参照Peri等[22]的方法,测定了雌蜂对老熟幼虫爬行痕迹的行为反应。将1个金属环(内径6.0 cm)放在一方形滤纸(20 cm×20 cm)的中部,环内放入1头老熟幼虫,在棉签刺激下使其爬行30 min 后取出。这样,该区域内就留有老熟幼虫的爬行痕迹,以此作为处理区,其他区域为未处理区。然后将滤纸放入帷帐中,其上方放置一个荧光灯管(20 W)。将1头雌蜂放入处理区后开始试验,当雌蜂飞走或爬出滤纸时结束,统计雌蜂在滤纸上的停留时间(在处理区和未处理区内的停留时间之和)。用未经老熟幼虫爬行过的滤纸(20 cm×20 cm)作为空白对照,并用同样的方法测定雌蜂在滤纸上的停留时间。重复测试5次后更换滤纸,每头寄生蜂只使用1次,处理和对照试验各重复25次,共50头雌蜂。
1.5 寄生蜂对茧或蛹浸提液的行为反应
为了检测微红梢斑螟茧中或蛹的体表是否存在寄生蜂用于寄主定位的化学线索,利用双选择试验测定了寄生蜂对茧或蛹体表浸提液的行为反应。
1.5.1 茧中或蛹体表信息物质的提取 将0.3 g茧(约4个茧)或10个蛹(2~5日龄),放入装有3 mL溶剂(甲醇、二氯甲烷或正己烷)的指形管中,4 ℃条件下浸提24 h,浸提液经微孔滤膜(0.45 μm)过滤后,用N2浓缩至200 μL备用。
1.5.2 生测方法 量取10 μL待测液及等体积的相应溶剂,分别施加于两个圆形滤纸(φ=1.0 cm)上,自然干燥5 min后,放入底部铺有滤纸的培养皿(φ=6.0 cm)中,两者对称放置于距培养皿中心1 cm的位置。为了避免雌蜂在刚放入培养皿时,因不适应环境而产生非正常的行为,在培养皿的中心放置一个顶部有孔(φ=0.1 cm)的塑料盖(φ=0.5 cm,H=0.4 cm),试验前用毛笔将寄生蜂放入塑料盖中,当寄生蜂从其顶部的孔中出来后开始计时,10 min后结束。将培养皿放入一个正上方有荧光灯管的围帐中,并使培养皿内的光强均匀一致,在培养皿的正上方放置一个摄像机记录雌蜂行为。用视频跟踪系统软件(Ethovison TX,Noldus),解析雌蜂首次接触到待测滤纸和对照的时间及在每个滤纸上的停留时间。
1.6 数据统计与分析
采用χ2检验,分析雌蜂对寄主植物和虫粪挥发物的趋性行为;雌蜂对老熟幼虫爬行痕迹的行为反应,通过Mann-Whitney U test分析;对茧或蛹浸提液的行为反应,则用Wilcoxon signed-rank test处理。
2 结果与分析
2.1 寄生蜂对不同处理球果挥发物的趋性行为
含蛹球果或含空蛹壳的球果挥发物对雌蜂均具有极显著的引诱作用(含蛹球果:χ2=7.50,P<0.01;含空蛹壳的球果:χ2=8.28,P<0.01),而幼虫为害的球果和健康球果无引诱活性(幼虫为害的球果:χ2=0.30,P>0.05;健康球果:χ2=0.833,P>0.05)(图1)。
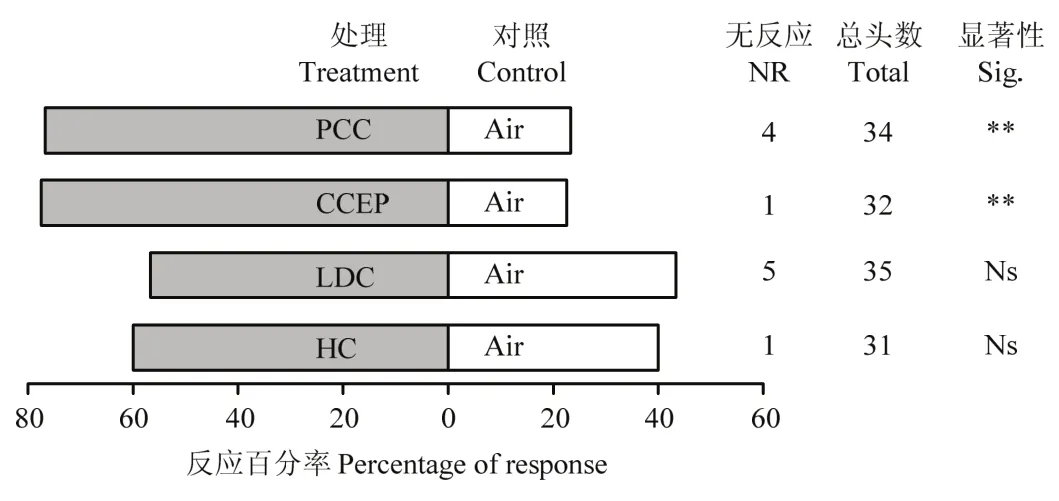
图1 寄生蜂对不同处理球果挥发物的趋性反应Fig. 1 The taxis responses of P. yunnanensis to the volatiles from the differently treated cones
2.2 寄生蜂对不同处理松枝挥发物的趋性行为
雌蜂对含蛹松枝挥发物表现为明显的正趋性行为(χ2>4.97,P<0.05),而对幼虫为害的松枝、含蛹球果的系统枝、含蛹松枝的系统枝和健康松枝无明显的趋性反应(幼虫为害松枝:χ2=0.78,P>0.05;含蛹球果的系统枝:χ2=0.89,P>0.05;含蛹松枝的系统枝:χ2=2.20,P>0.05;健康松枝:χ2=1.94,P>0.05)(图2)。

图2 寄生蜂对不同处理松枝挥发物的趋性反应Fig. 2 The taxis response of P. yunnanensis to the volatiles from different treatment of branches
2.3 寄生蜂对幼虫虫粪挥发物的趋性行为
含蛹球果内的虫粪挥发物对寄生蜂具有明显的引诱作用(χ2=4.32,P<0.05),而3龄幼虫虫粪无引诱活性(χ2=0.03,P>0.05)(图3)。
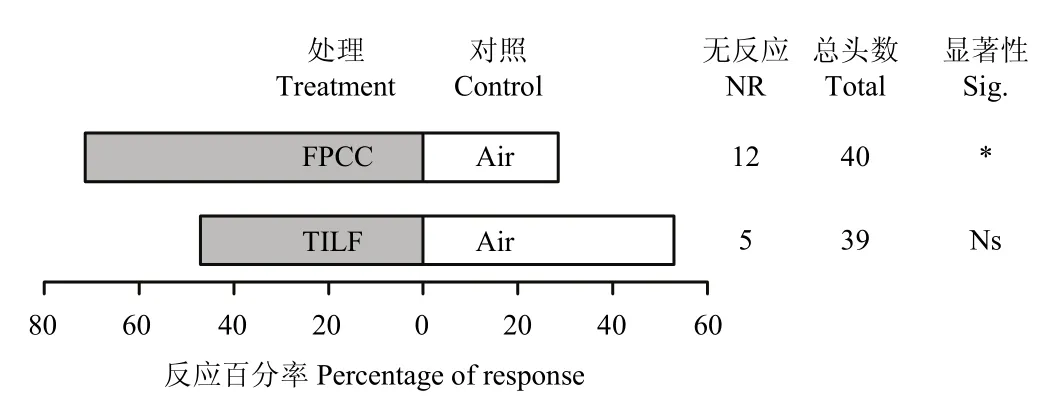
图3 寄生蜂对幼虫虫粪挥发物的趋性反应Fig. 3 The taxis response of P. yunnanensis to the volatiles of larva frass
2.4 寄生蜂对老熟幼虫爬行痕迹的行为反应
寄生蜂在处理滤纸和空白对照滤纸上的停留时间分别为(60.84±57.66)s和(52.20±44.06)s(均值±标准差),且两者间差异不显著(P=0.677),表明老熟幼虫的爬行痕迹对寄生蜂的行为无明显影响。
2.5 寄生蜂对茧浸提液的行为反应
无论溶剂是甲醇、二氯甲烷还是正己烷,寄生蜂首次接触到茧浸提液的时间均明显短于对照(甲醇:P=0.005;二氯甲烷:P=0.002;正己烷:P=0.004),并且在浸提液处理滤纸上的停留时间都显著长于对照(甲醇:P=0.004;二氯甲烷:P=0.001;正己烷:P=0.006)(表1)。
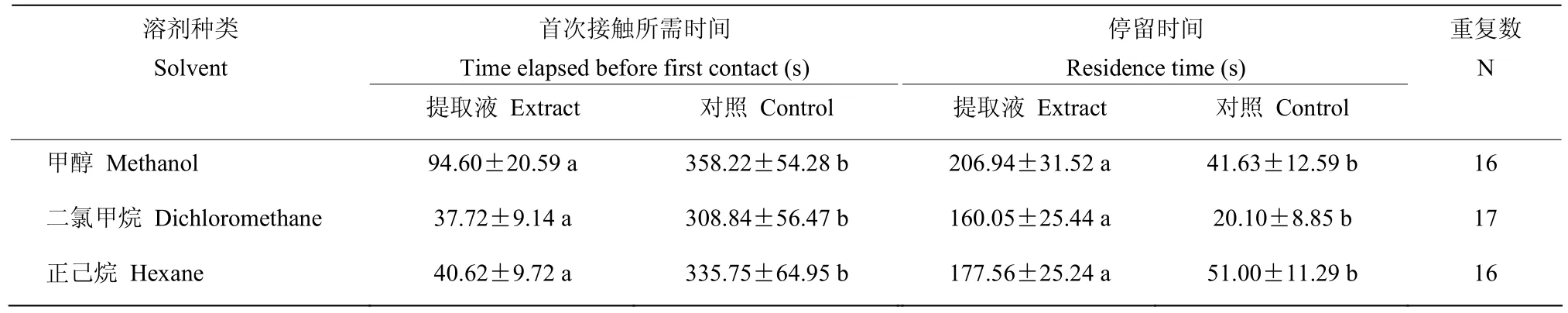
表1 寄生蜂对微红梢斑螟茧浸提液的行为反应Table 1 Behavioral responses of P. yunnanensis to the different extracts of D. rubella cocoon
2.6 蛹体表浸提液对寄生蜂行为的影响
寄生蜂首次接触到甲醇浸提液处理滤纸的时间,以及在滤纸上的停留时间与对照间均无显著差异(首次接触所需时间:P=0.255;停留时间:P=0.379),而首次接触到二氯甲烷或正己烷浸提液处理滤纸所需要的时间均明显短于对照(二氯甲烷:P=0.022;正己烷:P=0.041),并且在这两种处理滤纸上的停留时间均明显长于各自的对照(二氯甲烷:P=0.013;正己烷:P=0.021)(表2)。
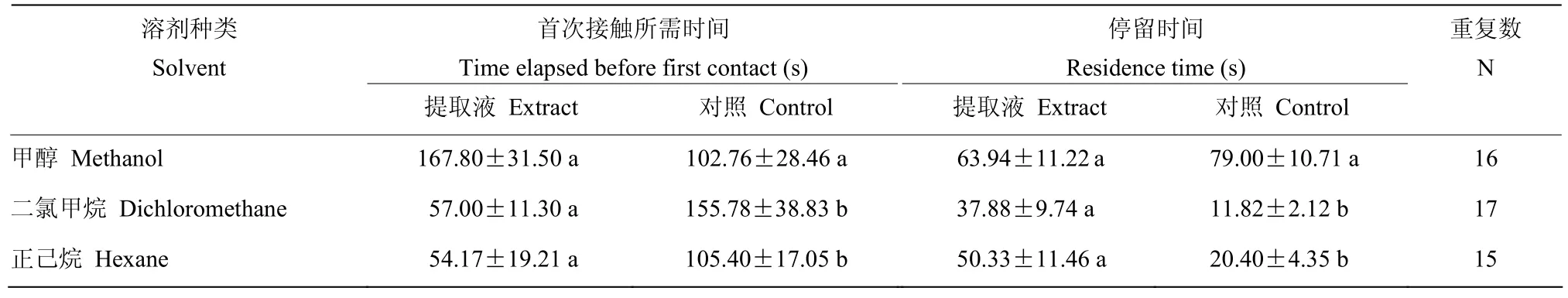
表2 寄生蜂对微红梢斑螟蛹提取液的行为反应Table 2 Behavioral responses of P. yunnanensis to the different extracts of D. rubella pupa
3 讨论
被害植物挥发物的组分及含量,会随着害虫虫态的不同而发生改变,进而影响寄生蜂的趋性行为[23,24]。欧洲粉蝶Pieris brassicae成虫产卵为害,能增强黑芥Brassica nigra对粉蝶盘绒茧蜂Cotesia glomerata的招引作用,但当卵发育成幼虫后,植株则表现出趋避作用[25]。与之类似,本研究中微红梢斑螟幼虫为害的松枝和球果对云南派姬小蜂的趋性行为均无显著影响,而含有微红梢斑螟蛹的球果和松枝则表现出明显的引诱作用(图1,图2),说明微红梢斑螟在从3龄幼虫发育为蛹的过程中,球果或松枝的挥发性组分发生了明显变化,在蛹期时释放出了引诱组分,为该寄生蜂的寄主栖境远程定位提供化学线索。
本研究还发现,含有空蛹壳的球果对寄生蜂有强烈的引诱作用(图1),说明成虫羽化后的球果依然持续释放引诱组分,目前,还未见类似现象的相关报道。其可能的生态学意义在于:微红梢斑螟在松林中为聚集分布,且世代重叠现象严重[14],成虫羽化后的球果中虽然已不存在寄主蛹,但其周围空间内仍可能大量存在其适宜寄生的寄主蛹(在含蛹球果内)。因此,成虫羽化后的球果持续释放引诱组分,对于寄生蜂的寄主栖境定位同样具有重要作用。
研究表明,虫害植株的受害部位和未受害部位,均能释放招引天敌昆虫的挥发物[26]。但本研究中,只有含蛹球果和含蛹松枝对该寄生蜂有引诱作用,而系统枝并无引诱活性(图 1,图 2),说明引诱活性物质仅来自被害部位。也正是因为如此,当云南派姬小蜂找到被害油松后,可能会更容易区分含蛹球果(或含蛹松枝)与其周围的系统枝,从而快速找到寄主的微栖境(含蛹球果或含蛹松枝)。
在进入寄主栖境后,与寄主昆虫相关的物质(如粪便、寄主分泌物、卵、茧和蛹)会为寄生蜂的寄主定位提供化学线索[3,4,6,9]。嗅觉测定结果显示,含蛹球果内的幼虫虫粪对云南派姬小蜂有明显的引诱作用(图3),表明其挥发物中有引诱组分,因其有较强的挥发性,它可能为已找到含蛹球果的寄生蜂,在搜寻并引导其进入羽化孔的过程中提供化学线索。与之类似,谷蠹Rhyzopertha dominica幼虫虫粪气味对雅脊金小蜂Theocolax elegans有强烈的吸引作用[27]。
与含蛹球果内的虫粪不同,3龄幼虫虫粪气味对云南派姬小蜂并无引诱活性,说明两种幼虫虫粪的活性组分间存在较大差异,其原因有 3种可能:1)与球果成分的变化有关。食物成分的差异对幼虫虫粪气味有重要影响,取食咖啡豆的小蠹虫Hypothenemus hampe幼虫虫粪,对肿腿蜂Prorops nasuta有强烈的吸引作用,而饲喂人工饲料的幼虫虫粪则无引诱作用。分析发现,幼虫虫粪中的引诱物质大部分来源于咖啡豆[28]。本研究中,健康球果和幼虫为害的球果无引诱活性,而含蛹球果的引诱作用显著(图1),表明随着幼虫的连续取食,球果成分发生了改变,产生了引诱物质,这些组分很可能通过取食进入4龄和5龄幼虫虫粪中,从而使含蛹球果内的虫粪表现出引诱作用。2)与幼虫虫龄有关。虫粪中的引诱组分很可能是幼虫的代谢产物,虫龄不同,其虫粪中的代谢产物也可能存在差异,从而导致引诱活性的变化。Chiu-Alvarado等[28]研究了咖啡果小蠹虫的幼虫虫粪对肿腿蜂Prorops nasuta趋性行为的影响,发现只有二龄及其以上虫龄的幼虫虫粪对寄生蜂才有引诱作用,且随着虫龄的增大引诱作用显著提高。对于微红梢斑螟而言,可能只有4~5龄幼虫才能合成引诱组分并进入到虫粪中。3)与虫粪发酵有关。多种昆虫虫粪发酵所产生的气味(如乙酸,乙醛)对寄生蜂有引诱作用[8,29]。球果内的幼虫虫粪在蛀道内存在时间较长,很可能在微生物的作用下,产生对云南派姬小蜂有吸引作用的组分。
微红梢斑螟老熟幼虫爬行痕迹对云南派姬小蜂的行为无明显影响,表明其爬行痕迹中不存在该寄生蜂用于寄主定位的信息物质,这可能与老熟幼虫并非该寄生蜂适宜的寄主虫态有关,在其他研究中也发现了类似的现象[8]。然而,核桃绕实蝇Rhagoletis completa和地中海实蝇Ceratitis capitata幼虫在爬行过程中留下的水溶性化合物能够明显延长蛹寄生蜂Coptera occidentalis的停留时间[30]。
在培养皿中,云南派姬小蜂首次接触到茧浸提液所需要的时间,均明显短于对照,且停留时间明显长于对照(表 1),表明微红梢斑螟的茧中存在对该寄生蜂具有引诱和滞留作用的物质。同样,苜蓿切叶蜂Megachile rotundata茧的正己烷浸提液在近距离对蜾蠃巨柄姬小蜂Melittobia digitata有极强的引诱作用[7]。
与茧类似,寄主蛹的体表也存在着调控寄生蜂寄主定位的信息物质,如美国白蛾蛹体表的烷烃类化合物在近距离对白蛾周氏啮小蜂有强烈的吸引作用[31]。微红梢斑螟蛹的二氯甲烷或正己烷浸提液对云南派姬小蜂有引诱和滞留作用(表2),说明在蛹的体表存在着对该寄生蜂具有短程吸引和接触识别的化学线索。笔者在显微镜下观察发现,该寄生蜂在接触到球果中完整的茧(含蛹)后,会不断撕咬茧的表面,直至咬出孔洞进入茧中为止,这很可能与茧所释放的信息物质有关,也可能是茧和蛹所释放的短距离信息物质共同作用的结果。
总之,云南派姬小蜂利用其寄主(含蛹球果内的虫粪、茧和蛹)及寄主植物(含蛹球果、含蛹松枝)相关的多种化学线索定位其寄主和寄主栖境。那么,这些化学信息物质是什么?有待进一步研究。
