基于“华占”的强优势杂交组合的株高与主穗长遗传分析
2021-02-22谢展文程红颂李忠金郑国源程祖锌林荔辉
谢展文 程红颂 李忠金 郑国源 程祖锌 林荔辉


摘 要:株高与穗长是水稻品种的关键性状,有效利用水稻性状间的杂种优势是培育高产品种的关键技术手段。以“华占”配制的‘天优华占‘恒丰优华占‘五丰优华占3个杂交组合为供试材料,构建了包含P1、P2、F1、F2、BC1-1、BC1-2等6个遗传世代群体。采用植物数量性状“主基因+多基因”混合模型分析了株高和主穗长的遗传方式。结果表明:3个杂交组合的株高和主穗长遗传均符合MX2-ADI-ADI遗传模型。在“华占”为遗传背景中,3个群体F2、BC1-1、BC1-2的株高均以多基因控制为主,多基因遗传率变幅分别为72.11%~75.42%、76.92%~86.28%和68.13%~75.13%,育种后代中应在高世代选择。3个群体F2代中的主穗长主基因遗传率分别为81.99%、68.95%和90.01%,多基因遗传率分别为12.16%、23.78%和3.52%;在回交群体中,3个组合的主穗长主基因和多基因遗传率与回交亲本密切相关,育种上应根据群体遗传特性进行相应的选择策略。
关键词:水稻;株高;主穗长;遗传模型;遗传分析
中图分类号:S511 文献标识码:A
Abstract: Plant height and panicle length are the key traits of rice varieties and the effective use of the heterosis of the traits is the key technical means to cultivate high-yield varieties. In this study, three hybrid combinations of ‘Tianyou Huazhan ‘Hengfengyou Huazhan and ‘Wufengyou Huazhan were selected as the test materials. Six genetic generations including P1, P2, F1, F2, BC1-1 and BC1-2 were constructed.The “main gene + polygene hybrid genetic model” was adopted for genetic analysis of plant height traits and panicle length in each generation. The plant height and panicle length inheritance of the three hybrid combinations were all consistent with the MX2-ADI-ADI genetic model. In the Huazhan genetic background, the plant height of F2, BC1-1 and BC1-2 was dominated by polygenic control. The heritability rate of the plant height traits ranged from 72.11% to 75.42%, 76.92% to 86.28%, and 68.13% to 75.13%. Breeding offspring should be selected in the higher generation.The main gene heritability of main panicle length in F2 generation of the three populations was 81.99%, 68.95% and 90.01%, and the polygenic heritability was 12.16%, 23.78% and 3.52%, respectively. In the backcross population, the main genes and polygenic heritability of panicle length in the three combinations were closely related to the backcross parents, so the selection strategies should be carried out according to the genetic characteristics of the population in breeding.
Keywords: rice (Oryza sativa L.); plant height; panicle length; genetic model; genetic analysis
DOI: 10.3969/j.issn.1000-2561.2021.01.009
水稻是重要的糧食作物之一,强优势杂交水稻组合的选育与利用是满足人民生活需求和保证粮食安全的重要途径。“华占”是当前我国水稻育种中一个重要且广泛应用的强恢复系之一[1]。该恢复系的一般配合力强,适应性好,配组并通过各级审定的组合多达133个,为我国配组组合最多的恢复系[2]。“华占”及其陆续培育系列衍生恢复系配制的杂交组合的推广应用对我国的水稻粮食生产起着举足轻重的作用。因此,剖析以“华占”为遗传背景的强优势杂交组合的关键性状遗传基础,对有效利用其杂种优势具有一定的参考价值。
水稻株高与穗长是水稻品种的主要关键性状之一。前者直接影响水稻品种的抗倒性和丰产性,决定了品种的地域适应性和应用前景[3]。主穗长也是水稻产量性状的主要构成因素,与穗粒数密切相关,直接影响水稻的产量[4-6]。因此,适宜的株高与穗长是水稻育种重要的目标之一,尤其在当前轻简化直播生产中,这2个性状是决定一个水稻品种能否大面积推广的关键因素,合适的株高和理想的穗部结构可有效提高水稻产量与适应性[7-9]。
水稻株高与主穗长是典型的数量性状,主要受主基因和微效多基因共同控制[10]。在水稻株高性状上,约有60多个基因陆续被克隆验证,如d1、d61、sd-1等基因[11-16];与水稻穗长相关的QTL也有大量的相关报道,如qPL6、qPL8、qPL1-1等基因[17-19],上述基因的功能定位与功能诠释对水稻的株高与穗长性状遗传改良具有重要价值。而在水稻实践育种中,这些基因在强优势杂交水稻的杂种优势贡献也是复杂的。因此,剖析以“华占”为遗传背景的强优势杂交组合的株高及主穗长的遗传基础及其遗传效应,评价了“华占”在配组的杂种一代株高与主穗长的遗传潜力和育种价值,对有效利用杂种优势以及后代的选择具有重要意义。
1 材料与方法
1.1 材料
2017年9月,采用3个籼型保持系‘天丰B‘五丰B‘恒丰B为母本,以“华占”为父本。配制‘天丰B/“华占”、‘恒丰B/“华占”、‘五丰B/“华占”3个杂交组合的F1代。2018年3月,在海南三亚分别采用相应恢复系和保持系与F1代回交,构建BC1-1、BC1-2世代。同时在F1代成熟时,收获F2种子,构建上述6个世代的遗传群体。上述材料均由福建农林大学农学院提供。
1.2 方法
1.2.1 田间种植 2018年5月,在福建省南平市水吉镇福建农林大学育种基地种植上述包括3个组合的双亲P1、P2、F1、F2,以及2个回交群体。其中F1各种植14行,每行7株,共计98株;各组合的回交组合BC1-1、BC1-2种植200株,F2种植800株,试验材料四周设置两行保护行。全部试验材料均单本插植,株行距为23.3 cm23.3 cm,田间栽培管理一致,具体措施如一般大田。
1.2.2 株高与主穗长的性状调查 在水稻植株成熟时,采用直尺测量水稻茎基部(齐泥)至穗顶部(不连芒)的高度,并同时测量主穗长,即穗茎节至穗顶部的长度。其中双亲P和杂种一代F1各调查30株,F2调查500株;回交组合BC1-1、BC1-2各调查150株。
1.3 遗传模型分析
采用植物数量性状“主基因+多基因”混合模型分析方法[20],利用R studio软件对3个杂交组合各世代的株高和主穗长性状进行分分析统计。运用极大似然法、IECM算法、通过AIC值比较和适合性检验,确定各群体株高和主穗长性状的最优遗传模型,估算2个性状的主基因和多基因遗传参数[20]。
2 结果与分析
2.1 不同世代的株高和主穗长的表型分析
3个供试杂交组合的6个世代的株高和主穗长均值与标准差见表1。在株高性状,“华占”平均株高为120.27 cm,‘天丰B‘恒丰B‘五丰B的株高较矮,分别为111.73、108.15和96.36 cm。3个组合的F2、BC1-1、BC1-2的株高和主穗长的群体频次分布图均呈连续分布,为典型的数量性状(图1)。在6个世代中,除了‘天优华占的BC1-2的株高均值低于双亲,‘天优华占和‘恒丰优华占2个组合的F2株高均值高于相应的双亲外,其他组合的F1、F2、BC1-1、BC1-2的群体的株高均值都居于双亲中间。在主穗长性状上,“华占”主穗长为20.74 cm;‘天丰B‘恒丰B以及‘五丰B的平均主穗长分别为20.95、20.34、和20.84 cm,亲本间的主穗长无明显差异。3个组合的F1代主穗长均高于双亲,变幅为21.01~21.25 cm,呈现出明显的杂种优势;在回交群体中,除了‘五丰优华占BC1-1的主穗长均值低于低值亲本外,其他5个回交群体均高于高值亲本,仍具备一定的杂种优势。F2的主穗长均值明显降低,杂种优势明显衰退。
2.2 主穗长和株高的最优遗传模型的确定
采用“主基因+多基因”混合遗传模型对以“华占”配制的3个强优势组合后代遗传群体的株高和主穗长性状进行联合分析,在48种遗传模型的极大似然值和AIC值中,取AIC值最小的3个值对应的模型列为表2。由表2可知,在株高性状上,‘天优华占株高的遗传模型MX2-ADI-ADI、PG-ADI、MX1-AD-ADI的3种模型AIC值分别为6483.363、6488.763、6490.445;‘恒丰优华占株高的遗传模型PG-ADI、MX1-AD-ADI、MX2- ADI-ADI的3种模型AIC值分别为6244.685、6248.302、6248.956;‘五丰优华占株高的遗传模型PG-ADI、MX1-AD-ADI、MX2-ADI-ADI的3种模型AIC值分别为6383.078、6386.421、6390.212。分别对上述3个模型分别进行适合性检验。结果表明,3个组合6个世代在均匀性检验(U12、U22、U32)、Smirnov检验(nW2)和Kolmogorov检验(Dn)中,‘天优华占的株高遗传模型3个统计量均达到显著性水平(P<0.05),根据AIC值最小准则,最佳模型为MX2-ADI-ADI(即2对加性?显性?上位性主基因+加性?显性?上位性多基因遗传模型,下同);而‘五丰优华占的3个模型中,PG-ADI、MX1-AD-ADI各有3个统计量达到显著性水平(P<0.05),仅MX2- ADI-ADI模型只有1个统计量达到显著性水平(P<0.05),故选择统计量达到显著水平最少的MX2-ADI-ADI作为最优模型。依此类推断可得,‘恒丰优华占的株高最佳模型亦为MX2- ADI-ADI。
在主穗长的遗传模型分析中,‘天优华占的MX2-ADI-ADI、MX2-A-AD、MX2-ADI-AD的3种遗传模型AIC值分别为3858.762、3875.995、3897.553;‘恒丰优华占的MX2-ADI-ADI、MX2-ADI-AD、MX2-A-AD的3种遗传模型AIC值分别为3877.552、3900.686、3913.751;‘五丰优华占的MX2-ADI-AD、MX2-ADI-ADI、MX2-A-AD的3种遗传模型AIC值分别为4286.086、4291.484、4310.029。与上述株高模型分析的方法相似,依据上述AIC值、相应的適合性检验及统计量显著性分析,3个杂交组合的主穗长最优遗传模型均为MX2-ADI-ADI。
综合上述分析结果,以“华占”配组的3个组合的株高与主穗长的最优遗传模型均为MX2- ADI-ADI,即2对加性?显性?上位性主基因+加性?显性?上位性多基因遗传模型,具有明显的遗传共性。
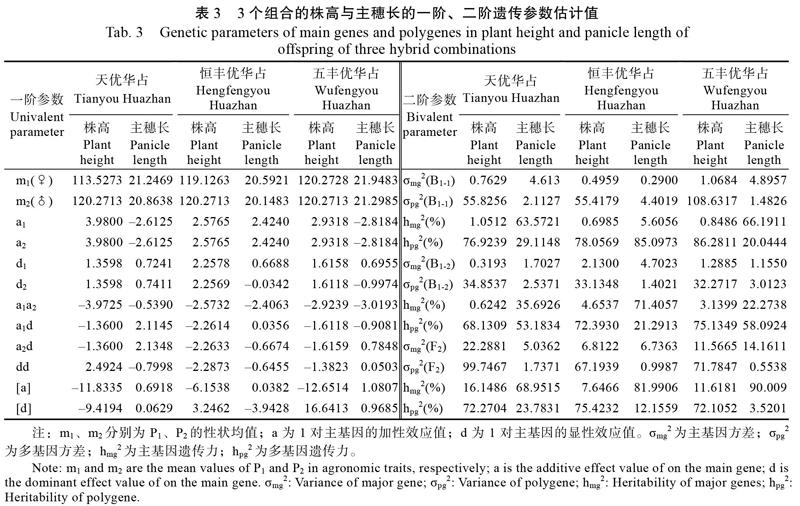
2.3 株高和主穗长的遗传模型及遗传参数
3个杂交组合的2个性状一阶、二阶参数分析见表3。由表3可知,3个组合后代的株高遗传均具有2对主基因的加性、显性、上位性效应。在一阶参数中,‘天优华占组合的2对主基因(基因1和基因2)加性效应与显性效应均相等,即a1=a2,d1=d2;加性效应为3.9800,显性效应为1.3598。2对纯合位点互作效应(a1a2)为?3.9725;纯合位点与杂合位点的互作效应值均为?1.3600,即a1d=a2d;2对杂合位点互作效应(dd)为2.4924;多基因加性效应[a]为?11.8335,显性效应[d]为?9.4194。而‘恒丰优华占和‘五丰优华占的株高2对主基因加性效应相等(即a1=a2),加性效应值2.5765和2.9318;显性效应值也近似相同(即d1≈d2);显性效应值分别为2.2578与2.2569和1.6158与1.6118;2对纯合位点互作效应值(a1a2)分别为?2.5732和?2.9239;2个组合基因1纯合位点与杂合位点的互作效应a1d分别为?2.2614和?1.6118;基因2纯合位点与杂合位点的互作效应的a2d分别为?2.2633和?1.6159,效应值相似;2对杂合位点互作效应(dd)分别为?2.2873和?1.3823。多基因加性效应[a]分别为?6.1538和?12.6514,显性效应分别为3.2462和16.6413。在株高性状,以“华占”配组的不同组合中,除了‘天优华占的2对杂合位点互作效应值(dd)为正值不同外,其他遗传参数正负赋值均相同,但不同组合间的遗传效应值存在一定的差异。
在主穗长性状遗传上,‘天优华占‘恒丰优华占和‘五丰优华占3个组合的2对主基因(基因1和基因2)加性效应与显性效应均相等(即a1=a2),分别为?2.6125、2.4240和?2.8184。其中,‘天优华占和‘五丰优华占组合加性效应为负值,为偏向母本(低值亲本)。在主基因显性效应上,基因1显性效应(d1)分别为0.7241、0.6688和0.6955;基因2的显性效应(d2)分别为0.7411、?0.0342和?0.9974;3个组合2对纯合位点互作效应值(a1a2)分别为?0.5390、?2.4063和?3.0193;纯合位点与杂合位点的互作效应值(a1d)分别为2.1145、0.0356和?0.9081,a2d值分别为2.1348、?0.6674和0.7848;杂合位点互作效应值(dd)分别为?0.7988、-0.6455和0.0503;多基因加性效应[a]分别为0.6918、0.0382和1.0807,显性效应[d]分别为0.0629、?3.9428和0.9685。綜合上述结果表明,在“华占”配组的杂交组合中,遗传背景不同组合的主穗长性状的最优遗传模型虽然一致,但各个遗传参数的效应值有所不同。
各性状的最优遗传模型二阶参数见表3。在株高性状上,‘天优华占组合的F2、BC1-1、BC1-2主要由多基因遗传为主,分别为72.27%、76.92%和68.13%,而主基因遗传分别为16.15%、1.05%和0.62%;‘恒丰优华占组合的F2、BC1-1、BC1-2的多基因遗传传率分别为75.42%、78.06%、72.39%,主基因的遗传率分别为7.65%、0.70%、4.65%;在‘五丰优华占组合中,F2、BC1-1、BC1-2的多基因遗传传率分别为72.11%、86.28%、75.13%,主基因的遗传率分别为11.62%、0.85%、3.14%。由此可知,3个组合各个群体后代的株高遗传上均以受多基因控制为主,且遗传率大小相近,遗传基础复杂,因此,在F2代或回交育种世代中,株高性状上应在高世代选择。
在主穗长性状上,‘天优华占组合的BC1-1群体中主穗长的主基因遗传率63.57%,多基因遗传率29.11%;BC1-2群体中主穗长的主基因遗传率35.69%,多基因遗传率53.69%,以多基因控制为主。F2代群体的主穗长主基因遗传率为68.95%,多基因遗传率为23.78%。但在‘恒丰优华占组合中,BC1-1群体中主穗长主基因遗传率为5.61%,多基因遗传率为85.10%;BC1-2的主穗长主基因遗传率为71.41%,多基因遗传率为21.29%,F2代群体主穗长性状的主基因遗传率为81.99%,多基因遗传率为12.16%。在‘五丰优华占组合中,BC1-1群体中主穗长主基因遗传率66.19%,多基因遗传率20.04%;BC1-2群体中主穗长的主基因遗传率22.27%,多基因遗传率58.09%,以多基因控制为主;F2代群体的主穗长主基因遗传率90.01%,而多基因遗传率仅3.52%,以主基因控制为主。综合上述结果,3个组合的F2代群体主穗长性状均由主基因遗传控制为主,但在不同组合的不同回交群体中,主穗长的遗传因回交亲本的差异,呈现不同的遗传方式,其中,‘天优华占和‘五丰优华占的遗传趋势相同,BC1-1以主基因控制为主,BC1-2则受多基因影响。而‘恒丰优华占的遗传则与上述2个组合相反。因而,在F2群体中的主穗长选择可在低代选择,而在不同回交群体则应根据亲本的不同而在不同世代进行选择。
3 讨论
3.1 株高和主穗長的遗传模型分析
水稻产量是由复杂数量性状构成[10]。本研究对3个以“华占”为骨干亲本的强优势杂交组合的株高和主穗长性状进行遗传模型分析,对在以“华占”为亲本的杂交组合的育种选择具有重要的指导意义。本研究表明,不同遗传群体株高或主穗长2个性状的遗传均符合“MX2-ADI-ADI”遗传模型(即2对加性?显性?上位性主基因+加性?显性?上位性多基因模型),具有明显的遗传共性。在株高的遗传模型上,这一结果与刘金波等[21]和陈献功等[22]采用不同组合和不同遗传群体的分析结果一致。同时,在主穗长的遗传模型上,陈志彬等[23]、陈献功等[22]、江建华等[24]认为主穗长符合2对“主基因+多基因”混合遗传模型,这与本研究结果也相似。但方兆伟等[25]以9311S 和WHR2 构建与本研究相同的遗传群体进行分析的结果则发现,株高和主穗长性状均受1对“主基因+多基因”的遗传,且主要受多基因控制。且在2个回交群体的主穗长以主基因遗传为主,而F2代的主穗长以多基因遗传为主。郑燕等[5]的研究结果则与方兆伟等[25]的研究结果相似。上述分析结果的差异可能与供试亲本组合及统计方法差异有关。
3.2 性状的遗传率与遗传效应
在3个组合F2、BC1-1、BC1-2的不同遗传群体中,株高均表现多基因遗传,遗传率变幅分别达72.11%~75.42%、76.92%~86.28%和68.13%~ 75.13%,且2对主基因与多基因的显性效应与加性效应值都具有同等重要。熊振民等[26]认为,不同的水稻品种株高是由多基因控制。方兆伟等[25]的研究也表明,株高主要受多基因控制,在F2群体中呈现株高主基因和多基因遗传同等重要,且表现加性效应遗传为主。樊叶杨等[27]利用协青早/密阳46的F2遗传群体也揭示控制株高性状的基因呈部分显性,并以加性?显性为主。因此,在水稻株高的遗传上是极其复杂的,在杂交组合的配组上,只有大量的测交筛选方能获得符合育种目标的杂交稻优势组合,同时在育种后代理想株型选择宜在高世代进行。3个杂交组合的F2代中主穗长的主基因遗传率分别为68.95%、81.99%和90.01%,多基因遗传率分别为23.78%、12.16%和3.52%,且以加性效应为主。在遗传贡献率上,这与蔡英杰等[28]的研究结果相近。而遗传效应上,邹文广等[29]则采用半双列杂交遗传群体的研究认为,主穗长同时受基因的加性效应和显性效应共同影响,以加性效应为主,但该研究结果发现,主穗长的遗传率低,宜在高世代选择,这一结论与本研究的结论不同。此外,本研究也发现,采用不同的亲本进行回交时,受双亲影响,BC1-1、BC1-2的主穗长遗传方式有所不同。因此,在回交育种加速性状稳定的同时,应根据不同组合的遗传特点进行定向选择,方可实现育种目标,有效利用杂种优势。
参考文献
张祖普, 华 荣, 贾长敬, 等. 优质三系杂交中稻武香优华占的特征特性及制种技术[J]. 现代农业科技, 2019, 741(7): 38-39.
李黎红. 中国水稻研究所植物新品种权保护之实践[J]. 中国种业, 2019 (2): 41-43.
邢永忠, 徐才国, 华金平, 等. 水稻株高和抽穗期基因的定位和分离[J]. 植物学报(英文版), 2001, 43(7): 721- 06.
郭小蛟, 张 涛, 蒋开锋, 等. 水稻籼粳交F8、F2群体穗长QTL比较分析[J]. 中国农业科学, 2013, 46(23): 4849-4857.
郑 燕, 王小明, 梁康迳, 等. 水稻长穗大粒特异种质的遗传分析[J]. 分子植物育种, 2011, 9(6): 672-679.
任德勇, 何光华, 凌英华, 等. 基于单片段代换系的水稻穗长QTL加性及其上位性效应[J]. 植物学报, 2010, 45(6): 662-669.
许乃霞, 杨益花, 苏祖芳. 抽穗后水稻株型与高光效群体形成关系的研究[J]. 耕作与栽培, 2009(5): 17-19.
王雪仁, 梁康迳, 林文雄, 等. 籼型三系杂交水稻株高的发育遗传分析[J]. 福建农林大学学报(自然科学版), 2002, 31(3): 279-282.
王雪仁, 梁康迳, 黄荣华, 等. 籼型三系杂交水稻穗颖花数和茎秆特性的遗传分析[J]. 福建农业大学学报, 2001, 30(1): 9-15.
郭龙彪, 罗利军, 邢永忠, 等. 水稻重要农艺性状的两年QTL剖析[J]. 中国水稻科学, 2003, 17(3): 211-218.
王松文, 施利利, 王妮妮, 等. 水稻sd-1基因分子标记的建立[J]. 天津农学院学报, 2000, 7(2): 52.
Li X, Gu M H, Xu J F, et al. Chromosome location of semidwarf gene sd t and sd n in indica rice[J]. Chinese Rice Research Newsletter, 2002, 10(1): 12-13.
Ma J. Genetic Identification of a New D1-allelic mutant and analysis of its gene function in rice[J]. Acta Agronomica Sinica, 2016, 42(9): 1261-1272.
高雅楠. 一个水稻D61基因等位突变体的表型分析及分子鉴定[D]. 聊城: 聊城大学, 2014.
张云辉, 张所兵, 林 静, 等. 水稻株高基因克隆及功能分析的研究进展[J]. 中国农学通报, 2014, 30(12): 1-7.
马玉银, 王如平, 李 磊, 等. 水稻株高的遗传与育种研究进展[J]. 河南农业科学, 2008 (11): 12-17.
蒋 强. 水稻穗长QTL定位[D]. 重庆: 重庆大学, 2008.
刘 颖, 叶生鑫, 彭 强, 等. 水稻穗长和有效穗数的QTL定位分析[J]. 江苏农业科学, 2016, 44(4): 86-89.
Zhang L, Wang J, Wang J, et al. Quantitative trait locus analysis and fine mapping of the qPL6 locus for panicle length in rice[J]. Theoretical & Applied Genetics, 2015, 128(6): 1151-1161.
盖钧镒, 章元明, 王建康. 植物数量性状遗传体系[M]. 北京: 科学出版社, 2003.
刘金波, 徐 波, 李建红,等. 水稻株高和每穗颖花数的6个世代联合遗传分析[J]. 华北农学报, 2017, 32(S1): 88-94.
陈献功, 洪德林. 水稻杂种汕优63F2代产量表现及株高遗传分析[J]. 雜交水稻, 2006, 21(4): 64-68.
陈志彬, 陈小荣. 水稻若干产量性状主基因+多基因混合遗传分析[C]//2010中国作物学会学术年会论文摘要集, 2010: 24-25.
江建华, 张启武, 洪德林. 粳稻穗角与谷粒性状及品质性状相关性分析[J]. 江西农业学报, 2008, 20(1): 1-4.
方兆伟, 刘金波, 王宝祥, 等. 水稻株高、穗长和每穗颖花数的遗传研究[J]. 江西农业学报, 2016, 28(5): 14-19.
熊振民, 闵绍楷, 俞桂林, 等. 云南稻种矮源的进一步筛选及其矮生性的遗传研究[J]. 江苏农学院学报, 1989, 10(3): 1-5.
樊叶杨, 庄杰云, 李 强, 等. 水稻株高QTL分析及其与产量QTL的关系[J]. 作物学报, 2001, 27(6): 915-922.
蔡英杰. 水稻长穗大粒RIL群体产量相关性状的遗传分析[D]. 福州: 福建农林大学, 2006.
邹文广, 许旭明, 张受刚, 等. 水稻长穗大粒种质资源产量性状配合力及遗传力分析[J]. 福建农业学报, 2014, 29(4): 324-328.
责任编辑:白 净
