Ultrastructural variation and key ER chaperones response induced by heat stress in intestinal cells of sea cucumber Apostichopus japonic u s*
2021-02-22ShashaWANGYingqiuZHENGMuyanCHENKennethSTOREY
Shasha WANG , , Yingqiu ZHENG , , Muyan CHEN , , Kenneth B. STOREY
1 Key Laboratory of Mariculture (Ocean University of China), Ministry of Education, Qingdao 266003, China
2 Institute of Biochemistry, Carleton University, Ottawa ON K1S 5B6, Canada
Abstract The unfolded protein response (UPR) is an important protective and compensatory strategy used during endoplasmic reticulum stress caused by factors including glucose starvation, low pH, or heat shock. However, there is very little information on the possible role(s) of the UPR under adverse conditions experienced by marine invertebrates. We observed that rough endoplasmic reticulum (ER) was dramatically expanded and numerous autophagosomes were accumulated in the intestinal cells of sea cucumbers, Apostichopus japonicus, under heat stress (4 h at 25 °C compared with 15 °C controls). Moreover, heat stress led to sharp increases in the relative transcript and protein expression levels of two primary ER chaperones: the endoplasmic reticulum resident protein 29-like (ERP29) and protein disulfide-isomerase A6-like (PDIA6). These results suggest a potential adaptive mechanism to deal with heat-induced stress in sea cucumber intestine.
Keyword: Apostichopus japonicus; endoplasmic reticulum (ER); unfolded protein response (UPR); endoplasmic reticulum resident protein 29-like (ERP29); protein disulfide-isomerase A6-like (PDIA6)
1 INTRODUCTION
In all eukaryotic cells, the endoplasmic reticulum (ER) is the central site for synthesis and folding of membrane and secretory proteins, and also participates in several important cellular functions including Ca2+storage and cell signaling (Bánhegyi et al., 2007; Fonseca et al., 2011). The lumen of the ER provides an oxidative environment that is specialized for the formation of disulfide bonds and proper protein folding for secretion or transportation. It has been proposed that 30% newly synthesized proteins are rapidly degraded, possibly as a result of improper protein folding (Schubert et al., 2000; Yewdell and Nicchitta, 2006). Thus, the situation becomes even more critical if there are any perturbations of these processes (Lai et al., 2007). The accumulation of unfolded or misfolded proteins in the ER induces the unfolded protein response (UPR), which is the major protective and compensatory mechanism used by the ER to adapt to changing environmental or stress conditions and, thereby, re-establish normal ER function (Schröder, 2006). These adaptive mechanisms of the UPR involve transcriptional programs that promote ER-associated protein degradation to remove misfolded proteins, or induce up-regulation of genes such as those coding for resident chaperones and folding enzymes (Ma and Hendershot, 2004) that enhance the protein folding capacity of the ER (Xu et al., 2005). Autophagy, as an intracellular degradative system, plays important roles in regulating protein homeostasis, and is essential for survival when cells are faced with various physiological or pathological stresses (Levine and Klionsky, 2004; Yorimitsu and Klionsky, 2005). During autophagy, double membrane vesicles termed autophagosomes are formed, which sequester damaged cytosolic proteins or organelles as cargoes and then fuse with vacuoles or lysosomes (Yorimitsu et al., 2006). Mounting evidence shows that the autophagic response can be induced by ER stress in eukaryotes (ranging from yeast to mammals) and is essential for normal cellular development and differentiation (McCullough et al., 2001; Back et al., 2005; Iwata et al., 2005). In addition to act in the repair of unfolded or misfolded proteins that accumulate under ER stress, ER chaperones are also primary sensors of ER stress in response to hypoxia, virus infection, starvation and heat shock in many animals including sea cucumbers (Ma and Hendershot, 2004; Wang et al., 2015). Previous studies have shown that these chaperones such as glucose regulated protein 94 (GRP94), endoplasmic reticulum resident protein 29-like (ERP29) and protein disulfideisomerase A6-like (PDIA6) act to fold new proteins eき ciently and refold stress-damaged proteins into their proper conformation (Hendrick and Hartl, 1995). However, very few studies have focused on the possible role of the UPR under adverse conditions in marine invertebrates (Chen et al., 2019).
The sea cucumber Apostichopus japonicus is a common species with high economic and medicinal values along the coasts of eastern Asia (Liao, 1997; Saito et al., 2002). As a typical temperate species, sea cucumbers are highly sensitive to changing variables of the marine environment (e.g. temperature, pH, salinity, etc.) (Asha and Muthiah, 2005; Wang et al., 2008). For example, temperature, as one of the most important abiotic factors, significantly affects the normal growth and metabolism of A. japonicus (Dong et al., 2007; Dong and Dong, 2008; Shao et al., 2015). In recent years, rapid temperature increasing and extreme high temperatures in the summer months has led to mass mortality of A. japonicus along the northern coasts of China, resulting in a dramatic resource decline and enormous economic loss, particularly for the aquaculture industry (Xu et al., 2017). Therefore, it is very important to understand the underlying molecular responses to heat stress in A. japonicus and identify regulatory mechanisms that can act to counteract effects of heat stress. The intestine of A. japonicus, the major tissue involved in food digestion and absorption of nutrients, has been reported to be highly sensitive to environmental change and to be a very important candidate organ for studies of the molecular responses to stress (Liao, 1997; Xu et al., 2016; Wang et al., 2018; Xu et al., 2018). To date, research on the molecular responses to heat stress in A. japonicus has mainly focused on expression of specific genes/proteins such as the heat shock protein (HSP) family and on epigenetic modifications such as microRNAs at an organismal level (Zhao et al., 2011; Chen et al., 2013; Xu et al., 2014; Wang et al., 2016; Chen et al., 2018; Gao et al., 2019). Due to the diき culty and immaturity of sea cucumber intestinal cell isolation and culture techniques, few studies have been conducted for endoplasmic reticulum stress response at the cellular and molecular levels (Wang et al., 2018), which will allow us to better understand the cytoprotective response to acute heat stress in sea cucumber intestinal cells. In the present study, we isolated sea cucumber intestinal cells by collagenase digestion and subjected them to heat stress. Ultrastructural changes in the intestinal cells were observed and changes in the expression levels of two key chaperones involved in ER stress were detected. We hope our current study will provide a candidate cytoprotective mechanism acting to counteract effects of heat stress in this marine invertebrate and help for heat-resistant variety selection of this valued economic species.
2 MATERIAL AND METHOD
2.1 Animal protocols
Twenty-one adult sea cucumbers A. japonicus (100±8 g) were collected from the coast of Qingdao (Jiaozhou Bay of the Yellow Sea, China) in May when seawater temperature was about 15 °C. The animals were acclimated in seawater aquarium system at 15 °C for one week at the Ocean University of China and fed with an artificial food compound (including kelp, scallop skirt, brown alga ( Sargassum thunbergii), gulfweed, Ulva and yeast) every morning. Subsequently, intestine samples from the mid-section of sea cucumbers were isolated and digested to release cells for experimental analysis. All animal procedures and experimental protocols were in compliance with the guidelines of the Experimental Animal Ethics Committee of Ocean University of China.
2.2 Cell culture and heat treatment
The cell culture and heat treatment were performed as previously described by Wang et al. (2018). Briefly, isolated intestines of A. japonicus from 12 individualswere treated with 0.125% collagenase IA (c9891, Sigma, USA) at 22 °C for 2-3 h as described previously (Odintsova et al., 2005). Cell suspension was obtained after centrifugation at the speed of 300× g for 2 min at 4 °C and filtered through cell strainer with a 70-μm mesh size. The eluted cell suspension was washed three times for 10 min each time in the artificial seawater suspending (CMFSS) dissolved with neomycin (10 mg/mL, N8090, Solarbio, Beijing, China) and amphotericin B (2.5 mg/mL, A8250, Solarbio, Beijing, China), followed by centrifugation for 10 min at 900× g. Then the cell pellets was resuspended in a modified Leibovitz’s L-15 medium (GNAM1300, Genom, Hangzhou, China) in 1.5-mL polyethylene tubes (Bello et al., 2015), and stabilized at 15 °C for half an hour for following experiments. Then all cells were then randomly divided into either control or experimental groups. The cells in the experimental group were heated at 25 °C for 4 h and were sampled at the same time with control group (15 °C).

Table 1 Primer information
2.3 Transmission electron microscopy
Samples used for transmission electron microscopy (TEM) were processed using standard techniques (Ullman et al., 2008). Briefly, the intestinal cells (from 15 °C and 25 °C groups) were centrifuged and fixed with 2.0% glutaraldehyde in seawater (pH 7.4, 1.1 Osm) overnight at 4 °C. After fixation, samples were placed in 1% osmium tetroxide with seawater for 2 h at room temperature and rapidly washed twice with sterile distilled water. Then, the samples were dehydrated in a graded series of ethyl alcohol and embedded in a mixture of Araldite and Epon 812. Ultrathin sections of 70-80 nm were cut with a Reichert-Jung Ultracut E ultramicrotome (Reichert, Austria) and placed on the Formvar coated slot copper grids. Sections were then counterstained with uranyl acetate followed by aqueous lead citrate and examined with a Jeol JEM 1200 transmission electron microscope (Japan). Digital images were acquired with an AMT XR-60 CCD Digital Camera System.
2.4 RNA isolation, cDNA synthesis, qRT-PCR and gene structure analysis
RNeasy mini kit (74104, Qiagen, Germany) with RNase-free DNase-treatment (79254, Qiagen, Germany) was applied to extract total RNA from isolated intestinal cells following the manufacturer’s instructions. For qRT-PCR analysis of intestinal cells in experiment group (25 °C) and control group (15 °C), first-strand cDNA was synthesized using PrimerScript™ RT reagent Kit (RR047A, TaKaRa, China) as the qPCR template, and the expression levels were measured in StepOnePlus (ABI Inc., USA) using SYBR®Premix Ex TaqTM(RR420, TaKaRa, Japan). Six biological replicates were detected for each group and cells from two individual intestines pooled as a biological replicate. Each sample was run in triplicate. The specific primers for AjERP29 (PIK36082.1) and AjPDIA6 (PIK49878.1) were listed in Table 1. β-tubulin (TUBB, PIK51093) and β-Actin (ACTB, PIK61412.1) were selected as the internal controls, according to previous studies (Zhao et al., 2014; Ono et al., 2018). Melting-curve analysis of the amplification products was performed at the end of the PCR reaction to confirm specificity.
The complete sequences of AjERP29 and AjPDIA6 were obtained from the A. japonicus genome reported by Zhang et al. (2017), which is publicly available from GenBank with accession number GCA_002754855.1. The promoter region sequence was predicted by Promoter 2.0 Prediction Server (http://www.cbs.dtu.dk/services/Promoter/). The transcription factor binding sites in the 5′-flanking region were predicted using the AiBaba 2.1 programs (http://gene-regulation.com/pub/programs/alibaba2/index.html). Multiple sequence alignments and phylogenetic-tree construction applying the neighbor joining (NJ) method were conducted using the ClustalW program and MEGA 6.0 software.
2.5 Preparation of cell protein extracts
Three biological replicates (cells from three individuals pooled as a biological replicate) were used for total protein extraction in treatment (25 °C) and control group (15 °C), respectively. Cell lysis buffer (phenylmethylsulfonyl fluoride added) (P0013, Beyotime, Beijing, China) was mixed gently with intestinal cells on ice after centrifuging at the speed of 100× g for 10 min. Then centrifuging (14 000× g, 5 min, 4 °C) to collect supernatant. Enhanced BCA protein assay kit (P0010S, Beyotime, Beijing, China) was used to measure soluble protein concentration in SpectraMax Plus (Molecular Devices, USA) system. The qualified protein was stored at -80 °C for use.
2.6 Protein Expression level of AjERP29 and AjPDIA6
The Western blot was applied to analyze the protein expression levels following the protocol as previously described (Chen et al., 2016). In brief, the protein samples (15-25 μg) were mixed with 4×Laemmli buffer (1610747.0, Beyotime, Beijing, China) and heated at 95 °C for 5 min. For sodium dodecyl sulfate polyacrylamide gels electrophoresis (SDS-PAGE), equal protein amounts were loaded into adjacent lanes on 12% gels and run for 2 h at 120V. Proteins were then transferred to polyvinylidene fluoride membrane (PVDF) (IPVH00010, Bedford, MA, USA) for 30 or 45 min at 280 mA following a wet transfer procedure (BioRad, USA). The membranes were blocked with 5% non-fat powdered milk in TBST (Tris Buffered Saline with Tween 20) for 2 h at room temperature and then washed five times for 10 min each on a gently shaking platform. Membranes were then incubated with one of the following primary antibodies: rabbit polyclonal antibody against AjERP29 (0.464 mg/mL, diluted 1:450 for cells in TBS with 5% non-fat powdered milk and 0.05% Tween-20, prepared by Genscipt, Nanjing, China) and rabbit monoclonal antibody against AjPDIA6 (0.364 mg/mL, diluted 1:330 in TBS with 5% non-fat powdered milk and 0.05% Tween-20, prepared by Genscipt, Nanjing, China) overnight at 4 °C with gentle shaking. The membranes were then washed with TBST five times for 10 min each and incubated with goat anti-rabbit IgG conjugated to horseradish peroxidase (1:10 000, 2578S, CST, Danvers, MA, USA) for 1 h at room temperature. β-Tubulin was chosen as the internal protein standard and incubated with rabbit polyclonal β-Tubulin antibody (1:1 000, 2146S, CST, Danvers, MA, USA) and goat anti-rabbit lgG, HRP-linked antibody (1:10 000, 7074S, CST, Danvers, MA, USA). The membranes were then washed with TBST five times for 10 min each and bound antibodies were detected by an ECL detection reagent (P0018, Beyotime, Beijing, China) according to the manufacturer’s instructions, followed by semiquantification analysis using the Image-Pro Plus 6.0 software (Media Cybernetics Inc., USA). Individual blots were repeated 3 times.
2.7 Statistical analysis
Relative expression levels of AjERP29 and AjPDIA6 at the level of transcription were evaluated by 2-ΔΔCtmethod. Values were normalized against the β-Tubulin and β-Actin transcripts, respectively, and then mean values were calculated. The results were statistically analyzed using the Student’s t-test with P <0.05 accepted as statistically significant (unless otherwise stated). All data are presented as mean±SE ( n=6 separate isolations of intestinal cells).
Relative protein abundance of AjERP29 and AjPDIA6 was normalized by β-Tubulin (Wang et al., 2018; Wang et al., 2019) and analyzed using the Student’s t-test with P <0.05 accepted as statistically significant. Results are presented as mean±SE ( n=3 independent protein isolations from different animals).
3 RESULT AND DISCUSSION
3.1 Homology and phylogenetic analyses of AjERP29 and AjPDIA6
Alignment of the deduced amino acid sequences of AjERP29 from A. japonicus with the amino acid sequences of ERP29 from other species showed the highest homology with the following marine invertebrates: Acanthaster planci (starfish, 51% identity), Lingula anatina (lingula, 48% identity), Branchiostoma belcheri (lancelet, 47% identity) and Crassostrea gigas (oyster, 47% identity) (Fig.1a). A phylogenetic tree based on the NJ method was constructed by analyzing the predicted amino acid sequences of ERP29 from different species (Fig.1b). The predicted AjERP29 sequence from A. japonicus clustered into a clade with another echinoderm species (starfish), and distinct from other invertebrate clades (Fig.1b).
The deduced amino acid sequence of AjPDIA6 from A. japonicus showed the highest identities of 59%, 59%, and 58% with those of Strongylocentrotus purpuratus (sea urchin), Acanthaster planci (starfish), and Lingula anatina (lingula), respectively (Fig.2a). A phylogenetic tree based on the NJ method was constructed by analyzing the predicted amino acid sequences of PDIA6 from different species (Fig.2b). The amino acid sequence of AjPDIA6 was also clustered into one clade with other echinoderms (sea urchin and starfish) (Fig.2b) and distincted from the vertebrate clade. The results for both proteins indicated that the evolution of ERP29 and PDIA6 are consistent with the known evolutionary position of echinoderm species. Furthermore, the high conservation of these two proteins across different animal phyla suggests that they may share a common important function across the animal kingdom.

Fig.1 Homology and phylogenetic analyses of the deduced amino acid sequence of AjERP29 compared with ERP29 from other invertebrate species

Fig.2 Homology and phylogenetic analyses of the deduced amino acid sequence of AjPDIA6 with PDIA6 from other species

Fig.3 Changes in the ultrastructure of sea cucumber intestinal cells under heat stress
We also observed a four-residue ER-retention motif (KEEL for AjERP29 and KDEL for AjPDIA6) at the carboxyl-terminus of the translated sequence (Fig.1a & b). These are typical characteristics of reticuloplasmins residing in the ER lumen (Scheel and Pelham, 1996), suggesting that both proteins are reticuloplasmins.
3.2 Ultrastructural observations of sea cucumber intestinal cells in response to heat stress
Obvious structural changes in intestine tissue under high temperature stress have been reported and studied previously in A. japonicus (Czechowski, 1996; DeSesso and Williams, 2008). Cell apoptosis after 192 h of heat stress and intensified ultrastructural damage with increasing time was also reported in A. japonicus (Xu et al., 2015). The present study provided the first ultrastructural analysis of isolated intestinal cells in A. japonicus after heat stress (Fig.3a & b). The most obvious structural change in the intestinal cells under heat stress was the expansion of the rough endoplasmic reticulum (Fig.3b, white arrowhead), which means that the diameter of the endoplasmic reticulum increased, but still kept its original structure and a mesh shape. It is interesting to note that we also observed numerous autophagosomes in the lumen of heat-treated cells (Fig.3b, black arrowhead) in comparison with control conditions. The images also show that the numbers of mitochondria and intracellular vesicles (lysosomes, residual bodies) decreased in stressed cells compared with the control group, suggesting that metabolic capacity and endocytosis/exocytosis functions were weakened in heat-stressed cells. Previous studies have indicated that ER stress is a potent inducer of autophagy, which resulted either in enhancing cell survival or in committing the cell to non-apoptotic death (Yorimitsu et al., 2006; Senft and Ronai, 2015). Thus, we deduced that an activation of autophagy may be induced through ER stress in sea cucumber intestinal cells under heat stress. Noticeably, the direct link between ER stress and autophagy has been rarely reported so far, and a series of questions such as the signal transduction pathway connecting ER stress and the role of autophagy induced by ER stress, remain unknown (Kabir et al., 2018). Ultrastructural changes caused by heat shock have been observed in various organelles inside the cells of other species. In spermatid cells of rats, mitochondrial degeneration, dilatation of smooth endoplasmic reticulum (SER) and enlarged intercellular spaces were recorded after hyperthermia (Kanter et al., 2013). Concentric stacking of rough endoplasmic reticulum (RER) and swelling of mitochondrial saccules were also reported under heat stress in tobacco Nicotiana sylvestris (Kandasamy and Kristen, 1989).
3.3 Promoter structure and the expression profiles in the intestinal cells against heat stress
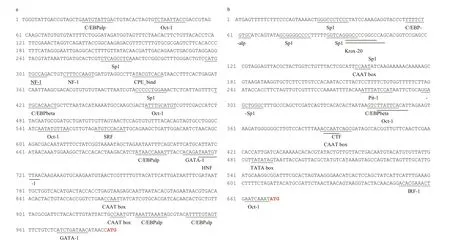
Fig.4 The putative transcriptional factor binding sites in the promoter sequence of AjERP29 (a) and AjPDIA6 (b) genes
Promoters of AjERP29 and AjPDIA6 were predicted and analyzed. Interestingly, in our present study, C/EBPbeta binding site was observed in both of AjERP29 and AjPDIA6 promoter regions (Fig.4) and it has been reported that the C/EBPbeta gene is induced in response to ER stress (Chen et al., 2004), which suggested that the variation of these two chaperones are potentially responsible for ER stress by binding with the activated C/EBPbeta gene. Growing evidence has suggested that reticuloplasmins might share common functions as a molecular chaperones during protein assembly and degradation and undergo up-regulated expression in response to cellular stresses such as improper protein folding or disrupted calcium homeostasis in the ER induced by biotic and abiotic factors (Kozutsumi et al., 1988; Harding et al., 2003; Ma and Hendershot, 2004; Schröder and Kaufman, 2005; Yin et al., 2017). The ER chaperones in these circumstances such as GRP94, ERP29 and PDI are primary sensors of ER stress and assist in repairing unfolded or misfolded proteins that accumulate in the ER under stress (Mkrtchian et al., 1998; Ma and Hendershot, 2002). In present study, we focused on the relative transcriptional and translational expression levels of EPR29 and PDIA6 in intestinal cells in response to heat stress. ERP29 has been shown to interact with and enhance the function of other ER chaperones, such as GRP94, GRP78, ERP72, and calnexin (Park et al., 2005), which suggested the important role of ERP29 in the regulation of cell survival and apoptosis. Protein disulfide isomerase A6-like (PDIA6) can also be considered as a key ER chaperone and is a multifunctional protein with a modular domain structure (Noiva and Lennarz, 1992). PDIA6 has been reported to participate in protein folding, assembly and post-translational modification by catalyzing native disulfide bond formation and rearrangement (Freedman et al., 1994; Tu and Weissman, 2004). Our present study showed that expression levels of AjERP29 mRNA transcripts increased significantly by 6.00 fold ( P <0.05) in intestinal cells in response to heat stress at 25 °C (Fig.5) and that AjERP29 protein expression levels also rose significantly by 2.88 fold under heat stress, as compared with controls ( P <0.05) (Fig.6). In addition, the transcriptional expression of AjPDIA6 was strongly increased by 3.66 fold at 25 °C ( P <0.05) (Fig.5) and the protein expression levels of AjPDIA6 rose by 2.09 fold in response to heat stress, compared to controls ( P <0.05) (Fig.6). Thus, we propose that AjERP29 and AjPDIA6 play important roles in the regulation of cell survival by alleviating ER stress caused by high temperatures in sea cucumbers. Previous studies have reported the overexpression of ERP29 and PDIA6 during heat stress associated with aestivation in sea cucumber intestinal tissue and coelomocytes (Zhang et al., 2013; Chen et al., 2016), which is supportive of our present study.
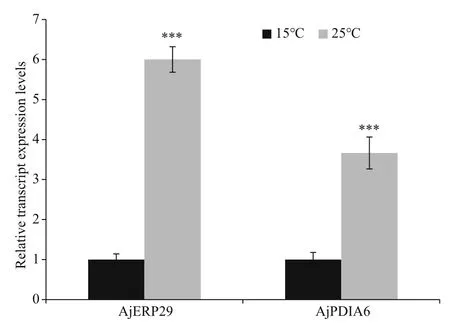
Fig.5 Relative transcript expression levels of AjERP29 and AjPDIA6 in intestinal cells in control (15 °C) and heat treatment (25 °C) groups, respectively
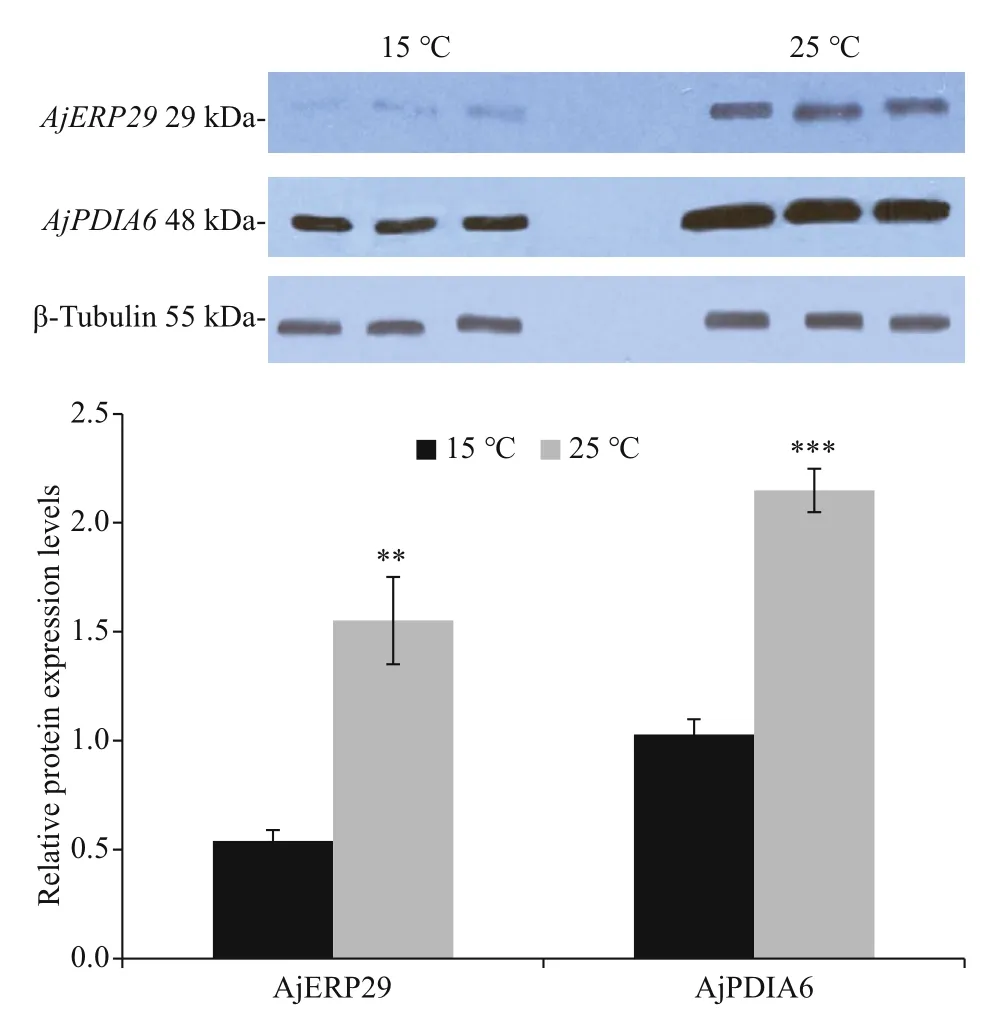
Fig.6 Relative protein expression levels of AjERP29 and AjPDIA6 in control group (15 °C) and experiment group (25 °C) using western blot
4 CONCLUSION
Heat stress may greatly influence the survival of sea cucumbers in summer. To our knowledge, this study provides the first demonstration of an endoplasmic reticulum stress response at the cellular and molecular levels in sea cucumber intestinal cells and allows us to better understand the potential cytoprotective mechanism in response to heat stress in sea cucumber, Which may fill the knowledge of thermal adaptive mechanism in sea cucumber and provide support for heat-resistant variety selection of this valued economic species. Further studies will also be needed to gain additional details about the regulatory pathway of the ER stress response in this species.
5 DATA AVAILABILITY STATEMENT
All data generated and/or analyzed during this study are included in this published article and its supplementary information files.
6 COMPETING INTERESTS STATEMENT
The authors declare no competing interests.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Erratum to: Arctic multiyear sea ice variability observed from satellites: a review*
- Application of an ice-ocean coupled model to Bohai Sea ice simulation*
- Expression and function of WNT4 involved in larvae development and limb regeneration in Portunus trituberculatus*
- Prevalence and risk assessment of antibiotics in riverine estuarine waters of Larut and Sangga Besar River, Perak*
- Relationships between sagittal otolith size and body size of Terapon jarbua (Teleostei, Terapontidae) in Malaysian waters*
- Effects of temperature on photosynthetic performance and nitrate reductase activity in vivo assay in Gracilariopsis lemaneiformis (Rhodophyta)*