湖南黄精属新种湘黄精的形态和分子特征分析
2021-02-07刘辉华马玉申王本忠揭红东向世军杨佳宁揭雨成
刘辉华,马玉申,王本忠,揭红东,向世军,杨佳宁,揭雨成,5*
(1湘西民族职业技术学院,湖南吉首 416000;2湖南农业大学苎麻研究所,长沙 410128;3高望界国家级自然保护区管理局,湖南古丈 416300;4吉首大学,湖南吉首 416106;5湖南省种质资源创新与资源利用重点实验室,长沙 410128)
黄精属传统上属于百合科,但分子生物学证据支持将它从百合科分出,隶属现在的天门冬科[1]。陈心启等认为,全球约有60种黄精属植物,中国有39种[2]。我国作为其主要分布中心,近年来不断有黄精属新种发现[3-7]。1875年,Baker根据叶序将黄精属分为3个组,分别为互叶组、轮叶组和对叶组[8];在此基础上,唐进结合花和苞片等形态将该属分为8个系[9]。随着细胞学、分子系统学的应用,黄精属植物的分类研究不断深入。Tamura[10]根据染色体基数与花丝的表皮特征将该属分为轮叶组(sect.Verticillata)和黄精组(sect.Polygonatum)2组,并将黄精组分为3个系。Meng等[11]基于叶绿体rbcL、trnK、psbA-trnH和trnC-petN片段分析,根据系统进化关系和地理分布将黄精属分为3大类群,其结果与Tamura的分组基本一致,只是将黄精(Polygonatum sibiricum)作为一个新类群。Zhao等[12]的最新分子系统研究也支持Meng等的分类结果,并将黄精属分为3组:互叶组(sect.Alternifolia)、轮叶组(sect.Verticillata)和黄精组(sect.Sibirica)。
2017—2018年湖南省草地资源调查中,笔者在湖南省古丈县高望界国家级自然保护区发现了一种黄精属存疑植物。该植物花被大小与多花黄精(Polygonatum cyrtonema)相近,叶形、叶先端等特征与黄精相似,通常一年发2次苗。通过查询国家标本资源共享平台(http://www.nsii.org.cn/2017/home.php)、中国数字植物标本馆(http://www.cvh.ac.cn/)、物 种2000中国节点(http://www.sp2000.org.cn.)以及JSTOR Global Pant(https://plants.jstor.org),进行形态特征比较和系统发育分析,确定其为黄精新种。因首次发现于湖南,故命名为湘黄精(Polygonatum hunanense H.H.Liu&B.Z.Wang)。湘黄精的发现对丰富中国黄精物种资源具有重要意义。
1 材料与方法
1.1 植物材料
湘黄精4个种群样本均采自湘西地区,具体地理信息如表1。凭证标本存放于吉首大学植物标本馆(JIU)。

表1 湘黄精种群采集地点信息Table 1 Collection location information of Rhizoma Polygonatum population
1.2 形态学分析
通过实地考察、对所采集的标本(包括样本的根、茎、叶片、花序、花梗、苞片、花被及裂片、花药、花丝、子房、花柱、浆果等)进行解剖、观察和测量、查阅文献资料[2]进行比较分析与特征描述。
1.3 样品序列信息获得
采用改良的CTAB法提取湘黄精样本新鲜叶片总DNA[13],参照Meng等[11]和Yan等[14]的方法设计引物(表2),参照Huang等[15]的方法进行PCR扩增,PCR产物经纯化后送上海生工测序获得样本序列信息。

表2 PCR引物序列信息Table 2 Primer sequences used for PCR
1.4 系统发育分析
将湘黄精的测序结果在NCBI网站进行Blast比对,选取同源性较高的21个黄精属代表种和6个外类群:长叶竹根七(Disporopsis longifolia)、散斑竹根七(D.aspersa)、竹根七(D.fuscopicta)、金佛山异黄精(Heteropolygonatum ginfushanicum)、异黄精(H.roseolum)和 总序鹿药(Maianthemum racemosum),分别从GenBank中下载4种叶绿体基因(rbcL、trnK、psbA-trnH 和trnC-petN)序列(表3),利用MEGA7.0对下载序列进行复核和调整[16]后,采用Clustal X2进行多重序列比对[17]和系统发育分析,并按照Zhao等[12]的方法采用最大简约法(MP)构建系统发育树。
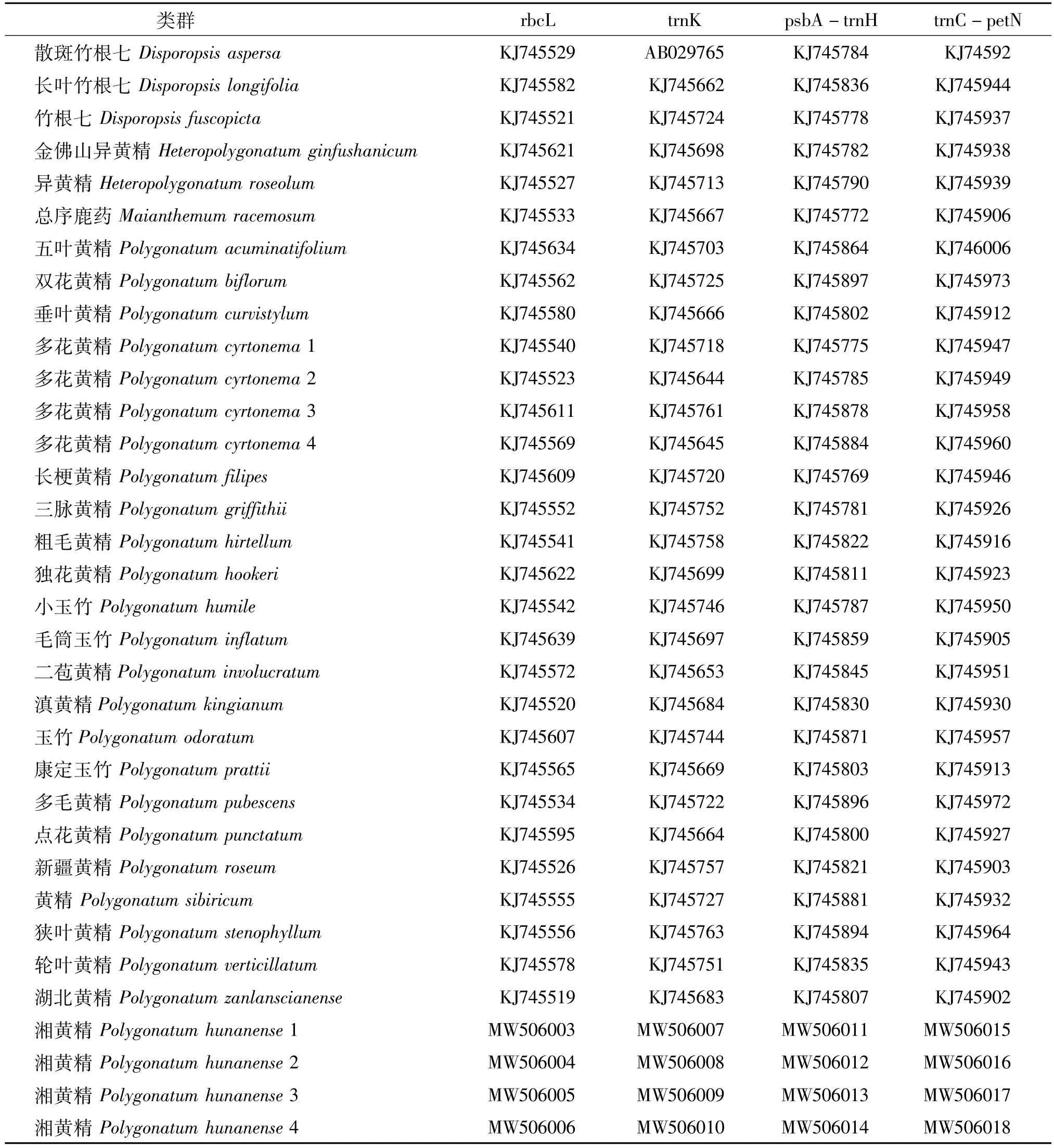
表3 各类群叶绿体基因的GenBank登录号Table 3 GenBank accession numbers of chloroplast genes in different populations
2 结果与分析
2.1 形态学描述
湘黄精:新种(图1、2)
Polygonatum hunanenseH.H.Liu & B.Z.Wang,sp.nov.(Fig.1&Fig.2)
The new species is closely related to P.cyrtonema and P.sibiricum,but it differs from the former by its leaves mostly whorled,linear-lanceolate or lanceolate,apex strongly cirrose.It differs from the latter by its peduncle 1.7-3.5 cm long,perianth 1.7-2.2 cm long,style 7-12 mm long.
Type:China,Hunan province,Guzhang county,Guanyin mountain,28°42′52.99″N,110°10′34.79″E,alt.349 m,9 June 2018,H.H.Liu& B.Z.Wang 2018060901a(Holotype:JIU!barcodes 83621).
根状茎近念珠状或姜块状,肉质肥厚,直径1.0~4.0 cm。全株光滑无毛,茎高1.0~3.5 m,顶端作攀缘状。叶多为轮生或近轮生,每轮3~6枚,栽培种达10枚,而上部和下部的叶片通常互生、对生或近对生,条状披针形或披针形,长5.0~20.0(~27.0)cm,宽5.0~25.0(~32.0)mm,背面有淡淡的白粉,先端通常强烈卷曲。花序具(1~)2~5朵花,伞形状,总花梗长1.7~3.5 cm,下垂,花梗长0.7~1.8 cm;苞片位于花梗基部,膜质,钻形至条状披针形,长3.0~4.0 mm;花被乳白色至淡黄绿色,圆柱状钟形,长1.7~2.2 cm,裂片长5.0~6.5 mm;花丝长2.0~3.0 mm,扁平,花药长约5.0 mm;子房椭圆状球形,长5.0~7.0 mm,花柱长9.0~12.0 mm,靠顶端通常弯曲。浆果,淡黄绿色或橙黄色,不规则球形,直径1.0~1.8 cm,具3~12粒种子。花期5至6月,果期8至12月(秋季发苗花期11至12月,果未见)。
该种生长于湖南西部河谷两岸海拔200~700 m的林下或灌丛中。由于长期大量采挖,现野生资源稀少,武陵山区有种植。

图1 湘黄精结构观察Fig.1 Structural Observation of Polygonatum hunanense H.H.Liu&B.Z.Wang
模式标本信息:湖南省古丈县观音山,北纬28°42′52.99″,东经110°10′34.79″,海拔349 m,2018年6月9日,刘辉华和王本忠2018060901a(JIU 83621)。
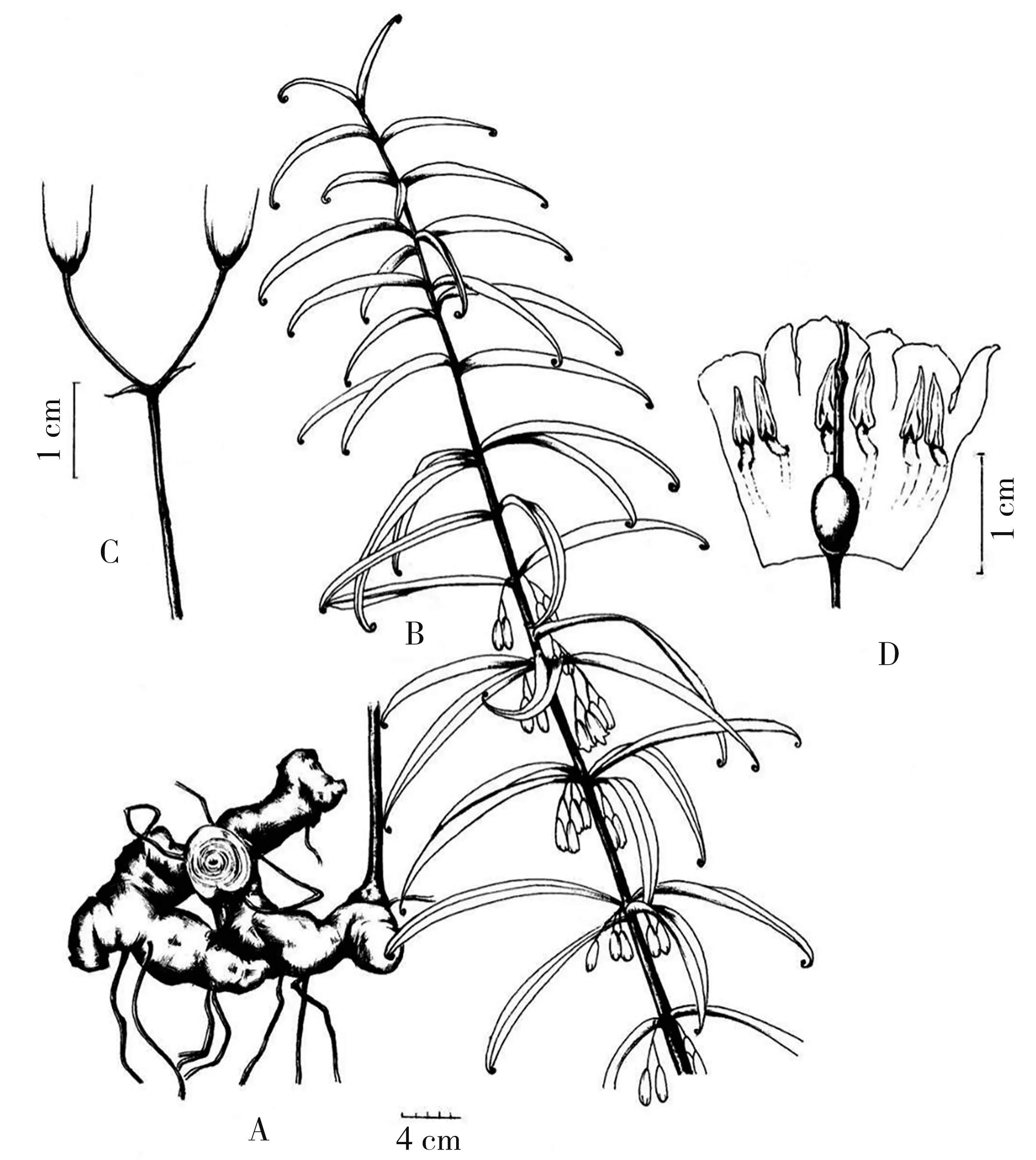
图2 湘黄精墨线描图Fig.2 The sketch of Polygonatum hunanense H.H.Liu&B.Z.Wang
2.2 系统发育分析
以长叶竹根七、散斑竹根七、竹根七、金佛山异黄精、异黄精和总序鹿药作为外群,以同源性较高的21个黄精属(多花黄精、双花黄精等代表种)采用最大简约法(MP)基于rbcL、trnK、psbA-trnH和trnC-petN序列(湘黄精各个片段序列Genbank号见表3)构建了系统发育树。结果显示,本研究构建的黄精属植物可分成3组(采用Zhao等[12]的分组名称),分别为互叶组、轮叶组及黄精组(图3)。湘黄精的4个株系可单独形成一个小分支且与多花黄精(P.cyrtonema)不同株系亲缘关系较近,但能明显区分,湘黄精的独立种的地位得到较好支持。
3 结论与讨论
系统发育树显示湘黄精单独形成一个小分支,与多花黄精亲缘关系较近。湘黄精与多花黄精的区别在于前者叶多为轮生、条状披针形或披针形,先端通常卷曲[2,9]。形态学比较和系统发育分析均支持湘黄精作为一个独立的种。湘黄精与多花黄精分布有重叠、花被大小相近等证据也表明亲缘关系较近,笔者将其划分在互叶组(sect.Alternifolia)[12],但其分组有待进一步探讨。
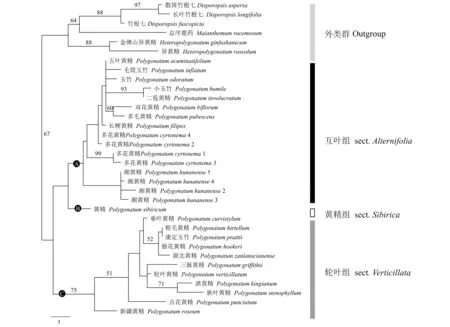
图3 基于rbcL,trnK,psbA-trnH和trnC-petN序列片段的湘黄精及其相关物种的最大简约法系统发育树Fig.3 Strict consensus tree illustrating the phylogeny of Polygonatum hunanense and related species generated by MP based on combined rbcL,trnK,psbA-trnH and trnC-petN sequences
