谷子MRP蛋白家族序列特征、分子进化及表达模式分析
2021-02-03高豪满夏夏孙朝霞韩渊怀李红英侯思宇郭数进
高豪 满夏夏 孙朝霞 韩渊怀 李红英 侯思宇 郭数进
摘要:本研究旨在深入了解谷子叶酸转运家族成员的基因结构和表达模式,为谷子体内叶酸转运分子机制的研究奠定基础。使用Phytozome、Clustal X、MEGA7.0等在线工具和软件,对谷子MRP家族成员进行生物信息学分析;基于谷子谷穗有参转录组测序数据,分析谷子MRP家族成员表达模式。结果显示,谷子MRP蛋白家族成员共21个,在7号染色体分布最多,有8个基因;17个谷子MRP蛋白偏碱性和4个偏酸性,根据疏水值判断其均为亲水性蛋白;成员SiMRP7、SiMRP12基因的表达量与谷子组织中的总叶酸含量表现出协同降低的趋势,推测这两个蛋白可能对谷子叶酸的积累起到调控作用。本研究有助于进一步鉴定叶酸转运相关蛋白,为后续研究叶酸代谢途径及谷子基因挖掘提供了理论基础。
关键词:谷子;MRP蛋白家族;谷子叶酸;生物信息学分析;表达模式
中图分类号:S515文献标志码:A论文编号:cjas2020-0059
MRP Protein Family of Foxtail Millet: Analysis of Sequence Characteristics, Molecular Evolution and Expression Pattern
Gao Hao, Man Xiaxia, Sun Zhaoxia, Han Yuanhuai, Li Hongying, Hou Siyu, Guo Shujin
(College of Agriculture, Shanxi Agricultural University, Taigu 030801, Shanxi, China)
Abstract: The aims are to study the gene structure and expression patterns of folate transport family members, and establish the foundation for exploring the molecular mechanism of folate transport in foxtail millet. Bioinformatics analysis of MRP family members was carried out with Phytozome, Clustal X, MEGA7.0 and other online tools and software. Based on the transcriptome sequencing data of foxtail millet, the expression patterns of foxtail millet MRP family members were analyzed. The results show that there are 21 foxtail millet MRP protein family members, most of which are distributed on chromosome 7, with 8 genes; 17 MRP proteins are alkaline and 4 proteins are acidic, and they are all hydrophilic proteins according to the hydrophobic value. The expression of SiMRP7 and SiMRP12 genes and the total folate content in foxtail millet tissue have a synergistic decrease trend, and these two proteins might play a regulatory role in the accumulation in foxtail millet. This study further identifies folate transport-related proteins, and could provide a theoretical basis for future research on folate metabolism pathway and gene mining.
Keywords: Foxtail Millet; MRP Protein Family; Folate of Foxtail Millet; Bioinformatics Analysis; Expression Pattern
0引言
谷子(Setaria italic)为禾本科植物,籽粒脱壳后被称为小米,产于中国北方地区[1]。作为农耕文明的一部分,曾是中国北方人民主要的粮食作物,在某种程度上决定了先民们的生产与生活方式[2]。随着生活水平的提高,人们开始追求健康和营养均衡的饮食,小米的营养保健和食疗价值都非常高,可以充分满足消费者对食品营养的需求。叶酸存在于各种动物性食品、粮食作物和绿色蔬菜中[3],是生物体内不可缺少的有机物质[4-5],也是食物成分表中有待补充的重要营养成分之一[6-7],人体不能自身合成叶酸,只有从膳食中摄取,粮食和蔬菜是人类叶酸的主要来源[8-9]。叶酸还具有补血,促进婴幼儿大脑的发育等功能,缺乏叶酸会导致神经衰弱、贫血、癌症等多种疾病[10-12]。
目前,全世界都广泛存在叶酸缺乏问题,中国也面临着严重的叶酸缺乏问题。据上海市闵行区妇幼保健院孕前门诊在备孕夫妻中进行的横断面调查,结果显示,总体人群膳食叶酸平均水平为296.3μg/d,男性为327.3μg/d,女性为267.2μg/d,低于成人400μg/d的推薦摄入量[13]。侯思宇等[14]建立了一种高效测定谷子籽粒叶酸的HPLC方法,同时测定了45份种质的叶酸含量,其平均叶酸含量最高达1.84μg/g,含量最高的种质的叶酸含量达到了2.86μg/g。邵丽华[15]测定了山西281份谷子种质资源的小米和36种主要粮食作物、常见水果和蔬菜的总叶酸含量,发现小米叶酸含量在粮食和蔬菜作物中属于较高水平,只略低于新鲜水果。
MRP蛋白(Multidrug resistance-associated protein)家族是跨膜转运蛋白超家族ABC家族的成员之一,其转运底物包括多糖、多肽、重金属螯合物、生物碱和药物等[16]。研究表明,玉米bronze-2 (bz2)突变体由于缺失bz2基因编码的一种谷胱甘肽S-转移酶,导致花色素苷不能在液泡中积累。因谷胱甘肽S-转移酶对MRP转运蛋白结合底物的活性有很重要影响,推测在玉米bz-2突变体中,MRP型ABC转运蛋白很可能参与花色素苷转运过程[17]。ZmMrp3是从玉米中克隆的一个编码ABC转运蛋白,定位在液泡膜上,是花色素苷转运至液泡中积累所必须的,而ZmMrp4转运蛋白也参与了花色素苷的积累过程,但在糊粉层组织大量表达[18]。王璐[19]以瓜叶菊蓝色、洋红色、黄色、粉色、白色5个色系植株为试验材料,利用RT-PCR技术分析瓜叶菊MRP亚家族14个不同成员在舌状花发育最初的2个阶段的表达模式,发现ScMRP1在不含花青素苷的黄色、白色系瓜叶菊中低表达,而在含有花青素苷的蓝色、洋红色、粉色系瓜叶菊中高表达,推测ScMRP1可能与瓜叶菊花青素苷转运相关。Garza等[20- 21]和Storozhenko等[22]发现,AtMRP1参与四氢叶酸的转运过程,在叶酸生物合成中起重要作用。
小米与其他大宗粮食比较,不仅营养丰富全面,而且具有特殊的医疗保健功能,所以深受国民尤其是中老年人的喜爱,更是产妇补身催乳的必备食品[23]。随着生活水平和习惯的变化,人们对健康饮食的需求越来越高,小米是优秀的营养保健和食疗食品,富含叶酸等营养物质,可以满足消费者均衡营养的需求。因全世界普遍存在的葉酸缺乏问题,叶酸的生物强化已成为一个新的研究热点,然而当前关于谷子富集叶酸的分子机制尚不清楚,尤其是叶酸代谢途径的关键基因的序列特征和表达模式尚无人报道。本研究利用生物信息学相关技术鉴定谷子的MRP基因家族并对该家族的序列特征、分子进化及表达模式进行分析,以期为谷子叶酸转运相关基因的研究和作物的叶酸生物强化研究提供一定的参考。
1材料与方法
1.1谷子MRP家族成员的序列提取及理化性质分析
在Pfam网站(http://pfam.xfam.org/)中查找MRP家族的保守域序列号(PF00005,PF00664)。在Phytozome网站(https://phytozome.jgi.doe.gov/pz/portal.html)下载基因位置信息。使用ProtParam网站(https://web. expasy.org/protparam/)计算氨基酸理化参数。
1.2谷子MRP家族蛋白系统进化树的构建
使用Clustal X进行序列比对,在TAIR网站(https://www.arabidopsis.org/index.jsp)下载拟南芥(Arabidopsis thaliana)MRP蛋白序列;在RGAP数据库(http://rice.plantbiology.msu.edu/)下载水稻(Oryza sativa)MRP蛋白序列;在MEGA7.0中使用邻接法(Neighbor-Joining,NJ)构建谷子、拟南芥和水稻的系统进化树。
1.3谷子MRP家族的蛋白质保守序列
利用MEME在线工具(http://meme-suite.org/)对谷子的MRP家族蛋白质保守基序进行分析,基序长度范围设为10~50个,基序最大发现数目设为10个,其他设为默认值。
1.4谷子MRP家族的蛋白质的二级结构、三级结构
使用SOPMA网站(https://npsa- prabi.ibcp.fr/cgibin/npsa_automat.pl?page=npsa_sopma.html)进行蛋白质二级结构预测,SWISS-MODLE服务器(https:// swissmodel.expasy.org/)进行蛋白质三级结构预测。
1.5谷子MRP家族成员表达模式分析
对谷子S1、S3、S5时期的穗部组织样品进行有参转录组测序,结合谷子S1、S3、S5时期的穗部组织总叶酸含量,根据谷子MRP家族成员在各时期的表达量,对谷子MRP家族成员的表达模式进行分析。
2结果与分析
2.1谷子MRP家族基因信息及理化性分析
鉴定出21个谷子MRP家族的基因,根据基因在染色体上的位置分布,按顺序对基因进行了命名,为SiMRP1-SiMRP21(表1)。基因序列长度范围在4413(SiMRP6)~17930(SiMRP10),蛋白序列长度范围在635(SiMRP5)~1609(SiMRP10)。其中氨基酸数目大于1300和小于1300的蛋白数目各为11个和10个,除了编码氨基酸数目最小的SiMRP5,其余MRP氨基酸数目跨度较小。对MRP蛋白理论等电点进行分析,结果表明,其中SiMRP14编码蛋白的等电点最小,为5.38,表现为酸性;SiMRP7编码蛋白的等电点最大,为9.32,表现为碱性。表现为碱性的蛋白有17个,表现为酸性的蛋白有4个,说明谷子MRP蛋白含有较多的碱性氨基酸和较少的酸性氨基酸。
对MRP蛋白的不稳定系数进行分析,结果表明, 21个蛋白的不稳定系数介于35.65~45.28,其中不稳定系数大于40的蛋白数目为9个,小于40的蛋白数目为12个,表现为不稳定蛋白的数目超过百分之五十。氨基酸的疏水性数值介于0.026~0.276,均为正值,表明谷子MRP蛋白均表现为亲水性(表1)。
MRP家族基因分别位于7条染色体上。其中有8个基因分布在7号染色体(Chr7),数目最多,其次是Chr1,有7个基因分布其上。剩余基因分布在其他6条染色体上(图1)。
2.2谷子MRP家族蛋白系统进化树分析
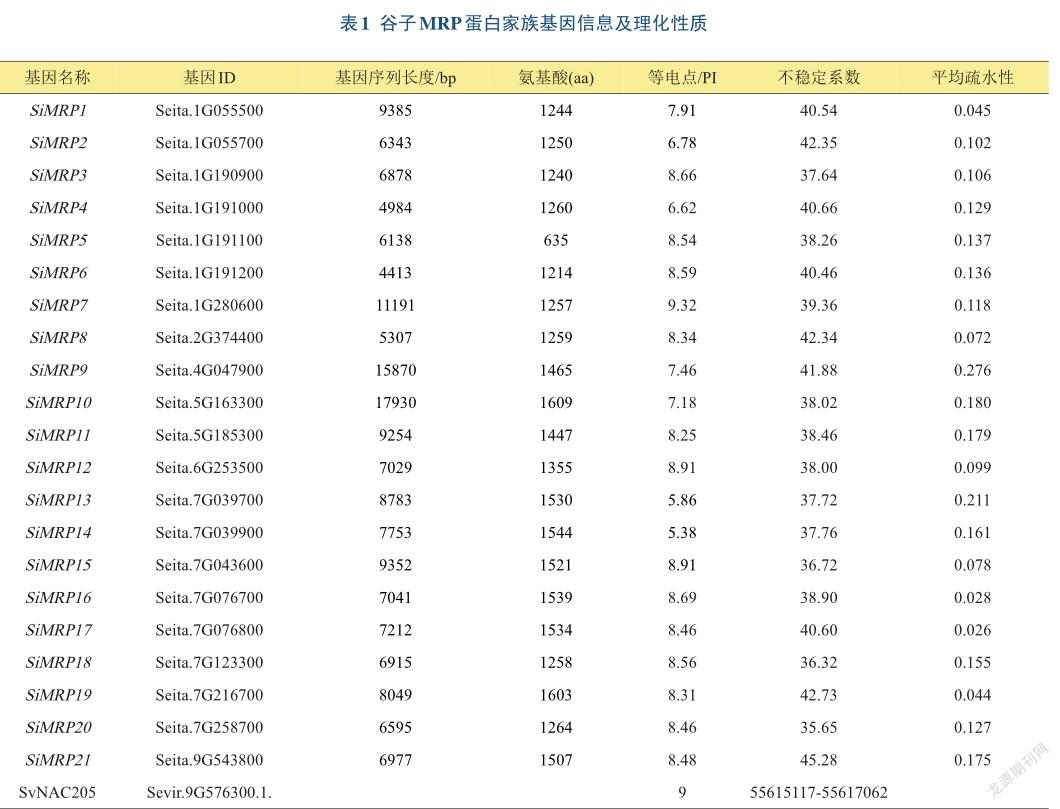
为了研究谷子MRP基因家族蛋白之间的亲缘关系,使用邻接法对谷子与拟南芥、水稻,3个物种共52个MRP蛋白序列进行分析,构建了系统进化树(图2)。根据谷子MRP家族系统进化树的分支情况,可以将谷子MRP蛋白成员分为两个进化分支,即ClassⅠ和ClassⅡ。进化树显示2个分支之间的蛋白质数量差距不大,其中ClassⅠ包含11个蛋白,成员为SiMRP1、SiMRP2、SiMRP3、SiMRP4、SiMRP5、SiMRP6、SiMRP7、SiMRP8、SiMRP12、SiMRP18、SiMRP20;ClassⅡ包含10个蛋白,分别为SiMRP9、SiMRP10、SiMRP11、SiMRP13、SiMRP14、SiMRP15、SiMRP16、SiMRP17、SiMRP19、SiMRP21。所有拟南芥和水稻MRP家族成员都聚类到了ClassⅡ中。从进化树的聚类分析可见,蛋白结构域相似的以及同源度高的蛋白聚在一起,如SiMRP1和SiMRP2,SiMRP16和SiMRP17,SiMRP18和SiMRP20,SiMRP21和OsMRP13,SiMRP11和OsMRP2聚类到了一起,通过蛋白序列比对发现SiMRP1和SiMRP2的同源性为82.51%,SiMRP16和SiMRP17的同源性为96.87%,SiMRP18和SiMRP20的同源性为86.73%,推测它们有较相似的功能。
2.3谷子MRP家族蛋白質结构域及保守基序分析
利用MEME对谷子的MRP家族21个蛋白质保守基序进行分析,结果表明,在MRP家族蛋白中,一共发现了10个保守基序(motif)(图3)。其中,SiMRP1、SiMRP2、SiMRP3具有完全相同的motif排列,SiMRP19、SiMRP20具有完全相同的motif排列,SiMRP16、SiMRP17具有完全相同的motif排列;有15个MRP蛋白的最后4个motif排列顺序是相同的,顺序为motif 10、motif 2、motif 3、motif 6;除了SiMRP5,所有成员的蛋白保守序列的倒数第3、4个motif都为motif 10、motif 2,这两个motif的排列顺序是固定不变的;图中显示这10个保守基序的氨基酸序列(图4)。

2.4谷子MRP家族蛋白质的二级结构、三级结构分析
利用SOPMA对谷子MRP家族的二级结构进行预测。蛋白质的二级结构包括无规则卷曲,延伸链,α螺旋,β折叠。谷子MRP家族在这4项中差别不大(表2),α螺旋最大为55.70%,最小为47.36%;延伸链最大为17.34%,最小为13.86%;β折叠最大为9.92%,最小为4.30%;无规则卷曲最大为30.82%,最小为 22.36%。根据MRP家族系统进化树结果,选择SiMRP1、SiMRP2、SiMRP8三个成员绘制蛋白质的二级结构和三级结构图,蛋白质的二级结构(图5)不但在某些方面上决定了蛋白质的三维结构,而且对蛋白质功能的预测有一定的帮助。

SiMRP1的α螺旋为52.33%,β折叠为5.71%,延伸链为17.20%和无规则卷曲为24.76%;SiMRP2的α螺旋为50.56%,β折叠为6.00%,延伸链为17.04%和无规则卷曲为26.40%;SiMRP8的α螺旋为50.28%,β折叠为6.04%,延伸链为17.24%和无规则卷曲为26.45%。这3个蛋白质的α螺旋,β折叠,延伸链和无规则卷曲的数值相差不大,蛋白的三维结构有一些的差别(图6)。
2.5谷子MRP家族成员表达模式分析
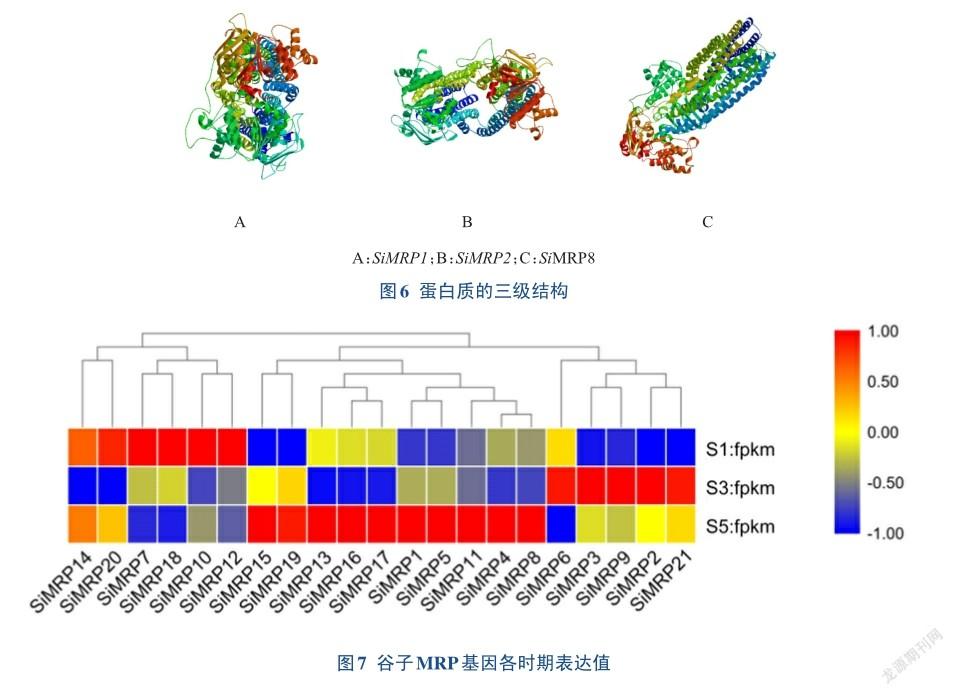
为进一步了解MRP家族成员的特性和功能作用,对其组织特异表达情况进行分析。所有的成员在谷子的S1、S3、S5时期组织中均有表达,SiMRP2在S1时期表达量最低,fpkm值为0.014,SiMRP10在S1时期表达量最高,fpkm值为46.013,SiMRP3在S3时期表达量最低,fpkm值为0.021,SiMRP21在S3时期表达量最高,fpkm值为38.654,SiMRP3在S5时期表达量最低,fpkm值为0.017,SiMRP21在S5时期表达量最低,fpkm值为33.164(图7)。其中SiMRP7、SiMRP9、SiMRP10、SiMRP12、SiMRP13、SiMRP18、SiMRP19、SiMRP20和SiMRP21在每个时期的表达量都很高,暗示着这9个基因有可能发挥多方面作用。不同的基因在不同时期表达量差异很大,如SiMRP7在S1时期的表达值是其在S5时期的2.42倍,SiMRP12在S1时期的表达值是其在S5时期的3.55倍,SiMRP10在谷子S1时期表达值最高,SiMRP21在S3、S5时期表达值都最高。结合谷子S1、S3、S5时期的谷子穗部组织总叶酸含量(图8),发现SiMRP7、SiMRP12基因的表达量与谷子组织中的总叶酸含量表现出协同降低的趋势,其基因在谷子不同时期的表达差异及功能机制还需要进一步研究。


3讨论与结论
谷子在中国具有长久的种植历史,是中国人民的传统粮食作物,在古代,它曾被作为中国北方主要粮食作物来种植,是中国北方人民喜爱的食物。随着社会的发展,谷子由于它丰富的营养、独特的风味,正日渐成为全国人民都喜爱的健康食物。谷子籽粒脱皮后被成为小米,其富含丰富的营养物质,包括多种维生素、蛋白质、脂肪、糖类及钙、磷、铁等人体所必需的营养物质,同时还具有一定的药用价值[24]。因谷子营养丰富,尤其富含叶酸,所以对谷子营养成分的富集机制的研究逐渐引起科学家的注意,谷子叶酸富集机制的探究更是成为一个新的研究热点。
目前谷子MRP蛋白家族的鉴定和系统分析尚未见报道,本研究对谷子MRP蛋白家族進行鉴定,并进行了一系列生物信息学分析。得到谷子的MRP家族一共有21个成员,并对该族蛋白序列进行了基序分析,发现谷子MRP蛋白家族具有保守性较高的特点。其一级结构之间和二级结构之间较为相似,三级结构之间的差异说明在进化的过程中这些蛋白序列发生了功能上的差异。谷子中亲缘关系较近的MRP蛋白一般具有相似的基因结构和蛋白结构域,如SiMRP18和SiMRP20蛋白的亲缘关系很近,其蛋白结构域也很相似,推测这对MRP基因很有可能来源于同一祖先基因,是基因复制的结果。SiMRP21和OsMRP13,SiMRP11和OsMRP2聚类到了一起,推断它们可能具有较高的同源性和较为相似的功能。在21个成员中共发现的10个保守基序,其中Motif 6和Motif 3非常相似,这可能会对结果造成一定的偏差。
研究表明,由MRP家族介导的氨甲喋呤(MTX)能通过竞争性抑制DHFR,阻断二氢叶酸(FH2)合成四氢叶酸(FH4),导致细胞内四氢叶酸不足[25- 26]。同时,AtMRP1也参与了到叶酸生物合成途径中的四氢叶酸的转运过程中[20-22],而SiMRP7、SiMRP12基因的表达量与谷子组织中的总叶酸含量表现出协同降低的趋势,推测这两个蛋白可能对谷子叶酸的积累起到调控作用。由此可以推断,谷子MRP蛋白家族很可能参与到了谷子叶酸的合成途径中,它可能在谷子叶酸合成途径中扮演了重要的角色。目前对MRP蛋白家族的研究主要集中在动物、癌症、药物领域,在植物领域中的研究比较少见,与谷子叶酸的合成途径的关系也未探明,因此本研究针对谷子MRP蛋白家族序列特征、分子进化及表达模式的分析有助于进一步探究谷子叶酸的合成途径,为挖掘谷子叶酸调控相关基因资源提供理论依据,为后续分子育种奠定基础。
参考文献
[1]刘宇杰,陈银焕,杨修仕,任贵兴.小米营养及功能成分研究进展[J].粮食与油脂,2020,33(5):1-3.
[2]张大众,刘佳佳.中国谷子种植利用史及其演进启示[J].草业学报, 2018,27(3):173-186.
[3]Cabalier O B, Allen L H. Encyclopedia of Human Nutrition[M]. Academic Press, 2012:262-269.
[4]Lin H L, An Q Z, Wang Q Z, et al. Folate In take and Pancreatic Cancer Risk:an Overall and Dose-response Meta-analysis[J].Public Health, 2013, 127(7):607-613.
[5]李松珍,李乐.浅谈孕早期叶酸合理应用的临床意义[J].中国保健, 2009(16):743-744.
[6]吴国芳,冯志坚,马炜梁,等.植物学[M].第二版.北京:高等教育出版社,1992:190.
[7]Packer L, Rimbach G, Virgili F. Antioxidant activity and biologic properties of a procyanidin-rich extract from pine (Pinus maritime) bark, pycnogenol[J]. Free Radical Biology and Medicine, 1999,27(5-6):704-724.
[8]Chen Y, Xie M Y, Gong X F. Microwave-assisted extraction used for the isolation of total triterpenoid saponins from Ganoderma atrum[J]. Journal of Food Engineering, 2007,81(1):162-170.
[9]傅荣杰,冯怡.微波萃取技术在中药及天然产物提取中的应用[J].中国中药杂志,2003(9):804-807.
[10]Koikeh. Clinicopathologic Features of Folate-deficiency Neuropathy[J].Neurology, 2015, 84(10):1026-1033.
[11]Duthies J. Folic Acid Deficiency and Cancer:Mechanisms of DNA Instability[J]. British Medical Bulletin, 1999, 55(3):578-592.
[12]Baggott Jje, Osterrva. Meta-analysis of Cancer Riskin Folic Acid Supplementation Trials[J].Cancer Epidemiology, 2012,36(1):78-81.
[13]季米,金龙妹,李春娟,等.备孕人群膳食叶酸营养状况横断面调查[J].中国循证儿科杂志,2018,13(6):401-405.
[14]侯思宇,宋敏,闫陆飞,等.HPLC法测定谷子籽粒叶酸含量及种质资源评价[J].土壤,2018,50(6):1235-1240.
[15]邵丽华.山西省小米叶酸含量的研究[D].临汾:山西师范大学, 2014.
[16]Dean M, Rzhetsky A, Allikmets R. The human ATPbinding cassette(ABC) transporter superfamily[J]. Genome Res, 2001,11:1156-1166.
[17]Marrs K A, Alfenito M R, Lloyd A M, et al. A glutathione S-transferase involved in vacuolar transfer encoded by the maize gene Bronze-2. Nature, 1995,375:397-400.
[18]Goodman C D, Casati P, Walbot V A. A multidrug resistanceassociated protein involved in anthocyanin transport in Zea mays[J]. Plant Cell,2004,16:1812-1826.
[19]王璐.瓜叶菊MRP基因家族的表达分析[C].中国园艺学会观赏园艺专业委员会.中国观赏园艺研究进展(2014).中国园艺学会观赏园艺专业委员会:中国园艺学会,2014:281-287.
[20]de la Garza, R I D, Gregory, J F, Hanson A D. Folate biofortification of tomato fruit[A]. Proceedings of the National Academy of Sciences, 2007,104(10):4218-4222.
[21]de la Garza, R I D,Quinlivan E P, Klaus S M J, et al. Folate biofortifi cation in tomatoes by engineering the pteridine branch of folate synthesis. Proceedings of the National Academy of Sciences of the United States ofAmerica, 2004,101(38):13720-13725.
[22]Storozhenko S, De Brouwer V, Volckaert M, et al. Folate fortifi cation of rice by metabolic engineering[J]. Nature Biotechnology, 2007,25(11):1277-1279.
[23]許寅生,郭亚丽,王玉祥,等.谷子的营养价值及产品开发[J].农业科技通讯,2018(3):152-155.
[24]张超,张晖,李冀新.小米的营养及应用研究进展[J].中国粮油学报,2007,22(1): 51-55,78.
[25]Banerjee D, Mayer-Kuckuk P, Capiaux G, et al. Novel aspects of resistance to drugs targeted to dihydrofolate reductase and thymidilate synthase[J]. Biochim Biophys Acta, 2002,1587(2- 3): 164-173.
[26]Gorlick R, Goker E, Trippett T, et al. Intrinsic and acquired resistance to methotrexate in acute leukemia[J]. N Engl J Med, 1996,335(14):1041-1048.
