鱼类血栓细胞免疫功能的研究现状
2021-02-03高迎莉韩高尚姚海静陈香凝周谦青
高迎莉, 韩高尚, 姚海静, 陈香凝, 周谦青
鱼类血栓细胞免疫功能的研究现状
高迎莉1, 2, 3, 韩高尚1, 姚海静1, 陈香凝1, 2, 3, 周谦青1
(1. 江苏海洋大学 海洋生命与水产学院, 江苏 连云港 222000; 2. 江苏省海洋生物技术重点实验室, 江苏 连云港 222000; 3. 江苏省海洋生物资源与环境重点实验室, 江苏 连云港 222000)
血细胞研究是鱼类基础研究的重中之重。鱼类血细胞的前期研究主要聚焦于淋巴细胞、巨噬细胞和嗜中性粒细胞等细胞的功能特性, 而忽略了血栓细胞。血栓细胞是低等脊椎动物具有的类似于哺乳动物血小板的一类细胞, 该细胞的经典功能是参与凝血和血栓形成。近年来的研究表明, 血栓细胞是一类特殊的免疫细胞, 在鱼体免疫应答中发挥重要作用。作者将对鱼类血栓细胞免疫功能相关的形态学、免疫学功能和研究前景等内容进行综述, 为鱼类血栓细胞功能的多样性提供基础资料。
形态学; 免疫功能; 血栓细胞; 鱼类
1882年, Giulio[1]描述血液中一种新的“形态学成分”, 命名为血小板(Platelets), 并发现血小板在机体出血和血栓形成中发挥作用, 因此Giulio被称为“血小板之父”。后来的研究发现, 哺乳动物无核的血小板来源于骨髓中的巨核细胞, 在非特异性和特异性免疫反应中发挥重要作用[2]; 低等脊椎动物如鸟类[3]、爬行类[4]、两栖类[5]和鱼类[6]中, 类似于血小板的细胞是有核的血栓细胞。近年来的研究表明, 血栓细胞参与鱼类的非特异性免疫反应以及炎症反应[7-9]。此外, 血栓细胞参与鱼类的特异性免疫应答的研究引起众多研究者的兴趣[7-8, 10-12]。鱼类的血栓细胞是否具有功能多样性, 有待于进一步的研究。作者将对鱼类血栓细胞免疫功能相关的形态学和细胞发生、免疫学功能等研究现状进行综述, 以全面阐述鱼类血细胞的新功能, 为鱼类血栓细胞功能的多样性提供基础资料。
1 血栓细胞的形态学特征
硬骨鱼血液中, 血栓细胞在数量上仅次于红细胞, 这些数量庞大的血栓细胞参与鱼体的血栓形成和凝血过程[6]。血栓细胞成群分布, 呈泪滴型、纺锤型、卵圆型或孤核型等多种形态, 具有指状突起、丝状伪足、囊泡、液泡、微管系统、颗粒、线粒体、高尔基体以及核糖体等成分[7, 13-23]。Jagadeeswaran[24]等发现斑马鱼()血栓细胞具有开放的管状系统, 血栓细胞活化后, 出现丝状伪足, 细胞发生凝集反应。斑点叉尾鮰()血栓细胞呈梭状, 胞内具有富含吞噬物质的液泡, 胞质中含有弥漫性的糖原颗粒[25](图1a)。聚盖鱼() 血栓细胞有梭形和球形两种, 细胞核有中心常染色质, 细胞质有液泡、管状系统以及吞噬体[26](图1b)。Azevedo[27]等的研究发现, 白纹笛鲷()血栓细胞被红细胞围着, 可形成多个胞质突起而呈现不规则细胞膜, 同时血栓细胞通过投影接近多个红细胞的细胞膜; 血栓细胞核内的异染色质和常染色质中包含部分颗粒原纤维团块及部分发育的液泡, 这些液泡有的位于胞质突起的顶端, 有的在细胞质中随机分布。最新研究表明, 人类血小板的细胞膜具有伪足和突起, 细胞内部具有液泡、α-颗粒、致密颗粒和溶酶体颗粒、管状系统、线粒体和自噬体。此外, 人类血小板的形态结构与年龄相关, 年龄增大, 细胞膜的规整性降低, 光滑性减少, 伪足和突起增多且更细长, 细胞内部的α-颗粒显著减少[28]。综上所述, 不同种鱼类的血栓细胞在形态结构上存在差异, 但如突起、伪足、囊泡、溶酶体等跟免疫相关的形态结构均出现在血栓细胞中, 这为血栓细胞的功能多样性奠定了结构基础。而人类血小板的形态结构是伴随时间发生变化, 因此不同鱼类血栓细胞的结构差异是否与鱼体的年龄、性别、健康状态等相关, 仍需要进一步探究。
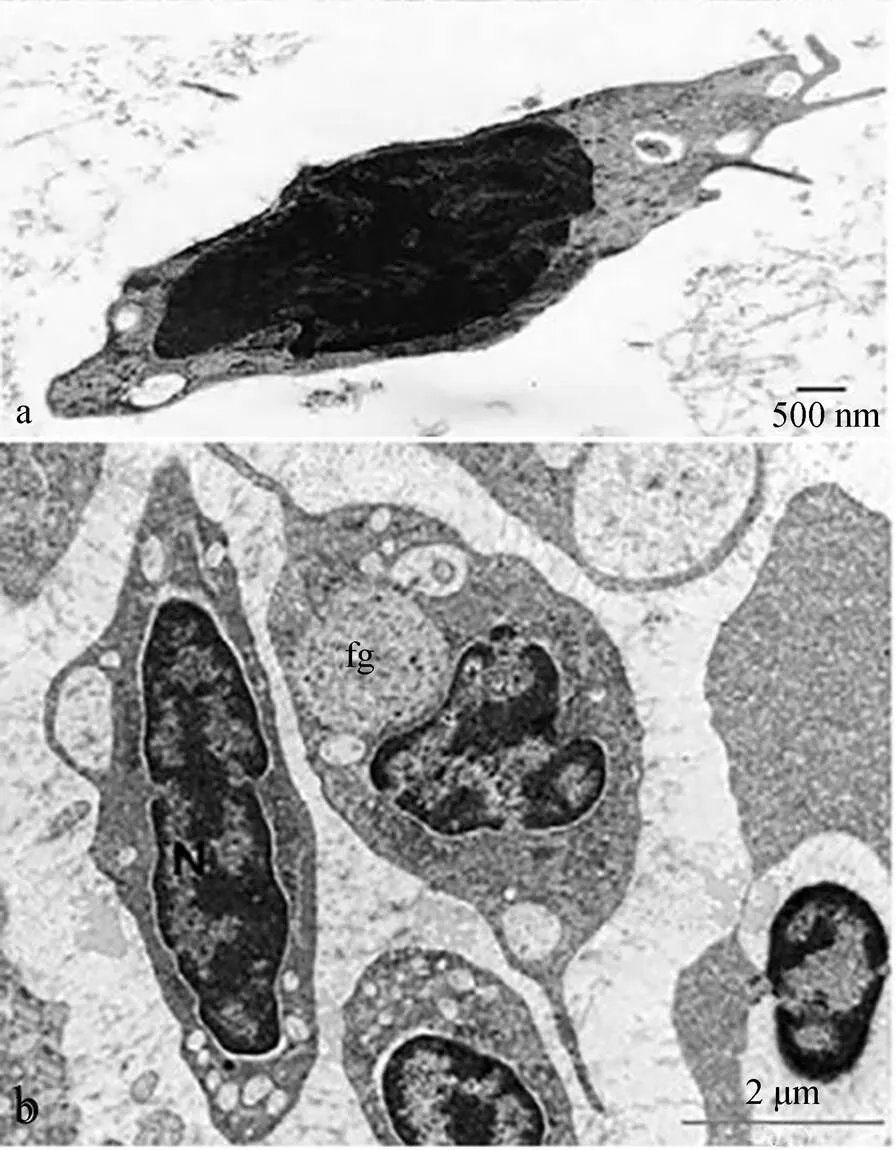
图1 血栓细胞的超微结构观察

a. Ultrastructure of thrombocytes in channel catfish. Fusiform thrombocyte with vacuoles, 1 of which contains phagocytosed material, bar=500 nm[25]; b. Ultrastructure of thrombocytes in fat snook. A fusiform thrombocyte to the left with its nucleus (N), to the right containing large phagosomes (fg), bar=2 μm[26]
细胞化学染色是以细胞形态学为基础, 根据某一特定胞内组分的化学反应原理, 在显微镜下观察细胞化学成分及其变化的显微研究方法。常用的有酶类(酸性磷酸酶ACP、碱性磷酸酶ALP、过氧化物酶Peroxidase、酯酶等)、脂类(苏丹黑B染色SBB)、糖原(过碘酸-希夫反应PAS)等细胞化学染色, 且常联合吉姆萨Giemsa染色用于区别不同细胞亚群或者用于细胞功能的研究(图2)。硬骨鱼类种类不同, 血栓细胞的细胞化学特性有差别, 如莫瑞鳕鱼(, Mitchell)[29](图3)等鱼类的血栓细胞呈现不同程度的细胞化学特性的相似性与差异性(表1)。除了蓝旗金枪鱼()[30], 所有鱼类的血栓细胞都是PAS阳性, 糖原是中性粒细胞进行胞内杀伤以及清除外来物质时所需的能量来源, 这从形态学角度证明鱼类血栓细胞的吞噬潜能。蓝旗金枪鱼是大洋洄游性鱼类, 其血栓细胞的功能与其他鱼类存在差异, 表明不同生境鱼类的血栓细胞会进化出与其生境或者生理活动相匹配的功能, 蓝旗金枪鱼血栓细胞是否具有吞噬功能需要进一步的研究。人类血小板的吞噬溶酶体中含有一些酶类, 如ACP, 这是血小板具有吞噬能力的证据[31]。由表1可知, 除了莫瑞鳕鱼、花斑裸鲤[32]和虹鳟[33], 其他鱼类血栓细胞都是ACP阳性, 表明这些鱼类的血栓细胞具有吞噬活性。但是, 莫瑞鳕鱼、花斑裸鲤和虹鳟的血栓细胞超微结果显示,细胞表面具有管状系统, 胞内有吞噬小泡和吞噬物, 说明这3种鱼类血栓细胞的吞噬活性是机械式而非酶活式。其他的细胞化学染色结果差别较大, 呈现明显的种属特异性, 如大盖具脂鲤和条纹鲮脂鲤的ALP染色结果相反; 铲吻油鲶[34]是苏丹黑B阴性; 热带鱼[35]是过氧化氢阴性, 但黄鲇鱼[36]和大口黑鲈[37]是过氧化氢阳性; 罗非鱼[38]仅仅检测了糖原特性。鱼类血栓细胞的形态学研究不仅可用于区分鱼类的不同血细胞类群, 而且可以直接观察血栓细胞的外部细胞膜、突起等结构以及内部细胞器等组成, 这为进一步研究血栓细胞的功能特性奠定结构基础。此外, 细胞化学特性为鱼类血栓细胞的种属差异性、年龄相关性、健康状态相关性、吞噬活性、免疫功能相关性等的研究提供直接证据。
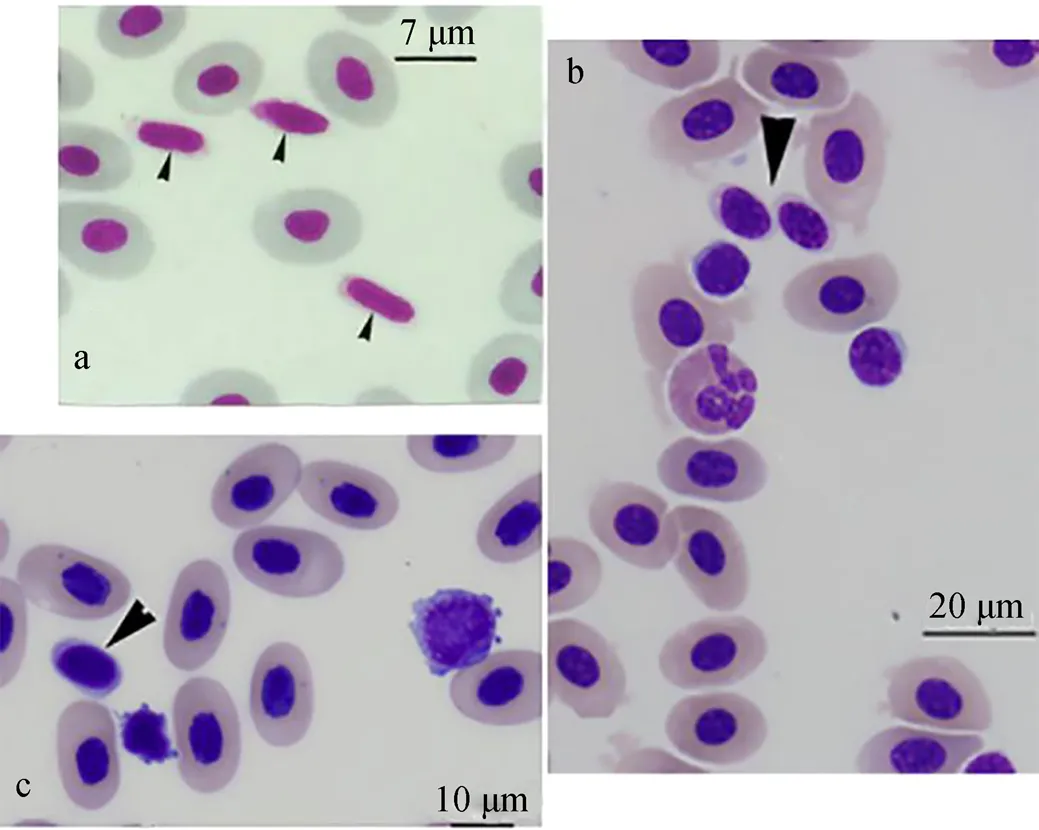
图2 鱼类血栓细胞的形态图
a. The thrombocytes of channel catfish (arrowhead) are smaller than the erythrocytes, with a spindle-shaped nucleus[25]; b. The thrombocyte of southern stingray (arrowhead)[23]; c. The thrombocyte of white sturgeon (arrowhead)[23]
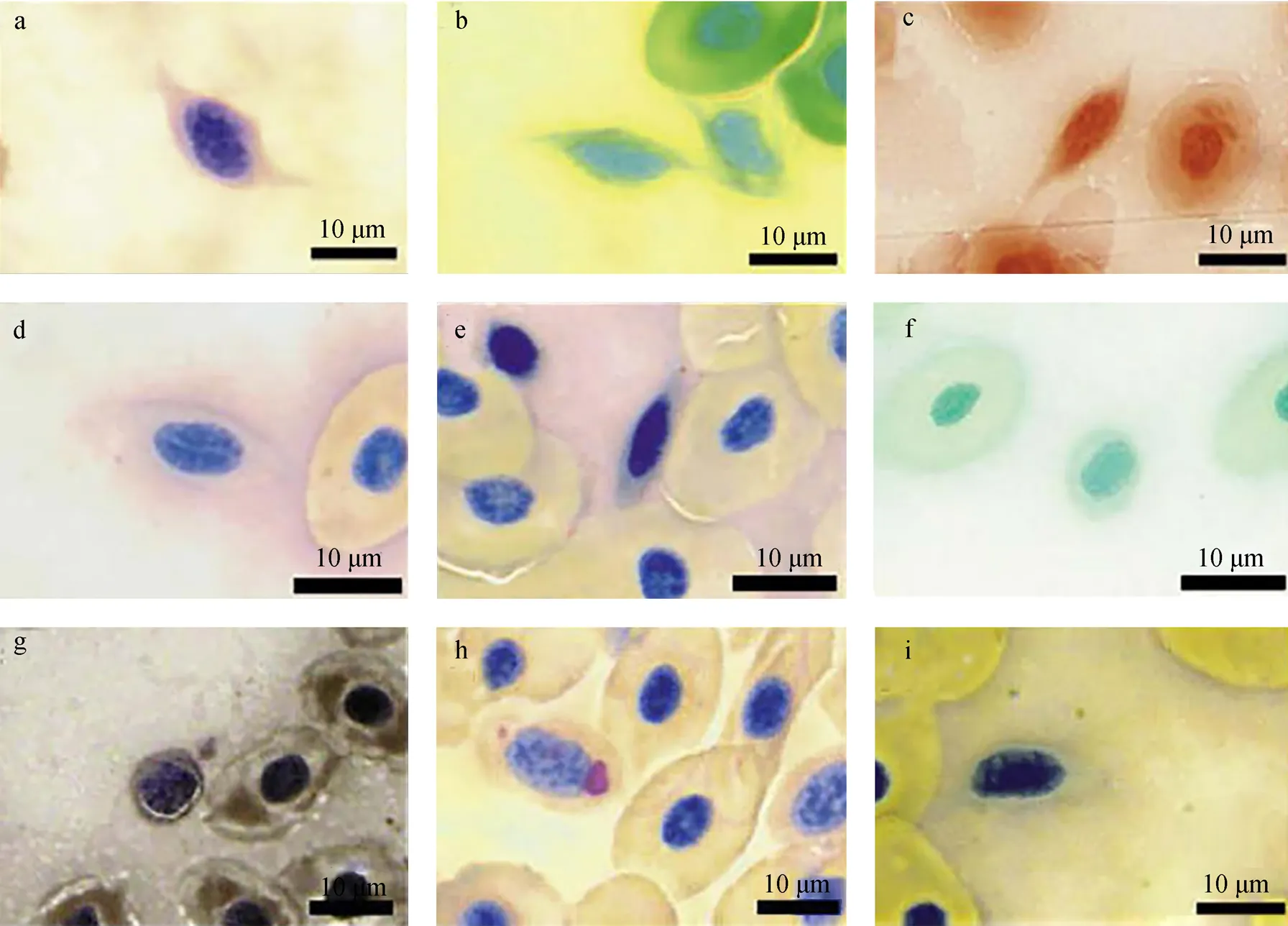
图3 莫瑞鳕鱼血栓细胞的细胞化学染色
a. 过氧化氢染色; b. 酸性磷酸酶(ACP)染色; c. 碱性磷酸酶(ALP)染色; d. 氯乙酸AS萘酚酯酶(NCE)染色; e. 乙酸萘酯酶(NAE)染色; f.α-丁酸萘酯酶(NBE)染色; g. 苏丹黑B(SBB)染色; h. 糖原(PAS)染色; i. β-葡糖醛酸糖苷酶染色。放大倍数×100, 标尺=10 μm[29]
a. peroxidase staining; b. acid phosphatase (ACP) staining; c. alkaline phosphatase (ALP) staining; d. naphthol AS chloroacetate esterase (NCE) staining; e. naphthyl acetate esterase (NAE) staining; f. α-naphthyl butyrate esterase (NBE) staining; g. Sudan black B (SBB) staining; h. periodic acid Shiff”s (PAS) staining; i. β-glucuronidase staining. Magnification × 100, Bar=10 μm[29]

表1 血栓细胞和血小板的细胞化学特性

续表
注: “+”. 阳性; “–”. 阴性; 鲤鱼[8, 10](); 胭脂鱼[17](); 条纹鲮脂鲤[25](); 大盖具脂鲤[25](); 聚盖鱼[26](); 莫瑞鳕鱼[29](, Mitchell); 蓝鳍金枪鱼[30](); 花斑裸鲤[32](); 虹鳟[33](); 铲吻油鲶[34](); 热带鱼[35](); 黄鲇鱼[36](); 大口黑鲈[37](); 罗非鱼[38]()
2 血栓细胞的免疫学功能
2.1 吞噬作用
吞噬作用是内吞作用的特殊形式, 包括颗粒的水泡内化, 形成内部囊泡即吞噬溶酶体, 是非特异性防御病原微生物的重要机制[39]。鱼类专业的吞噬细胞有单核/巨噬细胞[40]、嗜中性粒细胞[41]和树突状细胞[42], 此外, 淋巴细胞也具有吞噬作用[43-45]。关于上述各类细胞吞噬功能的研究多采用细胞系培养或细胞分离方法, 将胞内寄生虫、酵母菌、荧光微球和胞内寄生细菌等作为被吞噬的颗粒物, 应用细胞化学染色、光学显微镜、荧光显微镜、透射电镜以及流式细胞术等技术, 证明细胞的吞噬作用。
近年来的研究结果表明, 鱼类血栓细胞也具有吞噬作用, 但是血栓细胞在病原菌吞噬和清除上存在争议, 这可能是实验技术的不同造成的。细胞形态学结果显示, 血栓细胞胞内存在具有吞噬能力的颗粒[46], 然而, 细胞化学分析表明, 血栓细胞ACP呈阴性[37]。但人类血小板吞噬体酶类鉴定结果表示, ACP阳性是吞噬体不可或缺的成分。因此, 这种不一致是由于研究方法的局限性, 还是由于鱼类血栓细胞的细胞化学特殊性, 有待于进一步研究。脊椎动物的血小板吞噬作用具有4个特征, 且在鱼类血栓细胞中也有体现: (1) 血栓细胞与颗粒性物质(非生物颗粒、病原体等)相互作用。鲤鱼血栓细胞可以摄入并杀伤金黄色酿脓葡萄球菌()[46]以及大肠埃希氏菌()[8]; Burrows[47]等发现, 瘤棘鲆()血栓细胞能够在体外主动吞噬碳颗粒, 具有较强的吞噬能力; 牙鲆()和鲤鱼血栓细胞(图4)能够在体外吞噬荧光微球[8]; (2) 抗原刺激后, 血栓细胞胞质的溶酶体产物被释放, 导致胞内杀菌作用。白纹笛鲷血栓细胞的超微结构显示, 抗原刺激后, 胞内微管释放空泡物质, 而该物质是血栓细胞吞噬作用的产物, 从细胞结构上印证血栓细胞的吞噬作用[27]; 鲤鱼血栓细胞吞噬溶酶体融合实验表明, 血栓细胞可以内化细菌且具有杀菌能力[8]; (3)吞噬作用的代谢产物能作为炎症反应的媒介。Ferdous[3]研究了鸡血栓细胞吞噬作用, 证明其代谢产物介导炎症反应。而Nagasawa[12]等通过研究血栓细胞吞噬作用与白细胞分泌的激活因子之间的关系, 发现血栓细胞的吞噬活性会因白细胞分泌激活因子的增加而增加, 从而使其代谢产物在急性炎症反应中发挥作用; (4)利用补体, 通过调理素作用增强吞噬消化功能。研究表明, 鲤鱼血清对血栓细胞的吞噬活力起到增强的作用[8]。以上关于鱼类血栓细胞吞噬作用研究主要集中于体外吞噬, 血栓细胞体内吞噬研究涉及较少, 研究方法单一, 缺乏多种方法的联合使用以及方法间的相互佐证, 且血栓细胞吞噬作用介导的胞内杀伤作用等仍然不够明确, 这在很大程度上限制对鱼类血栓细胞抗菌功能的认知与探索。
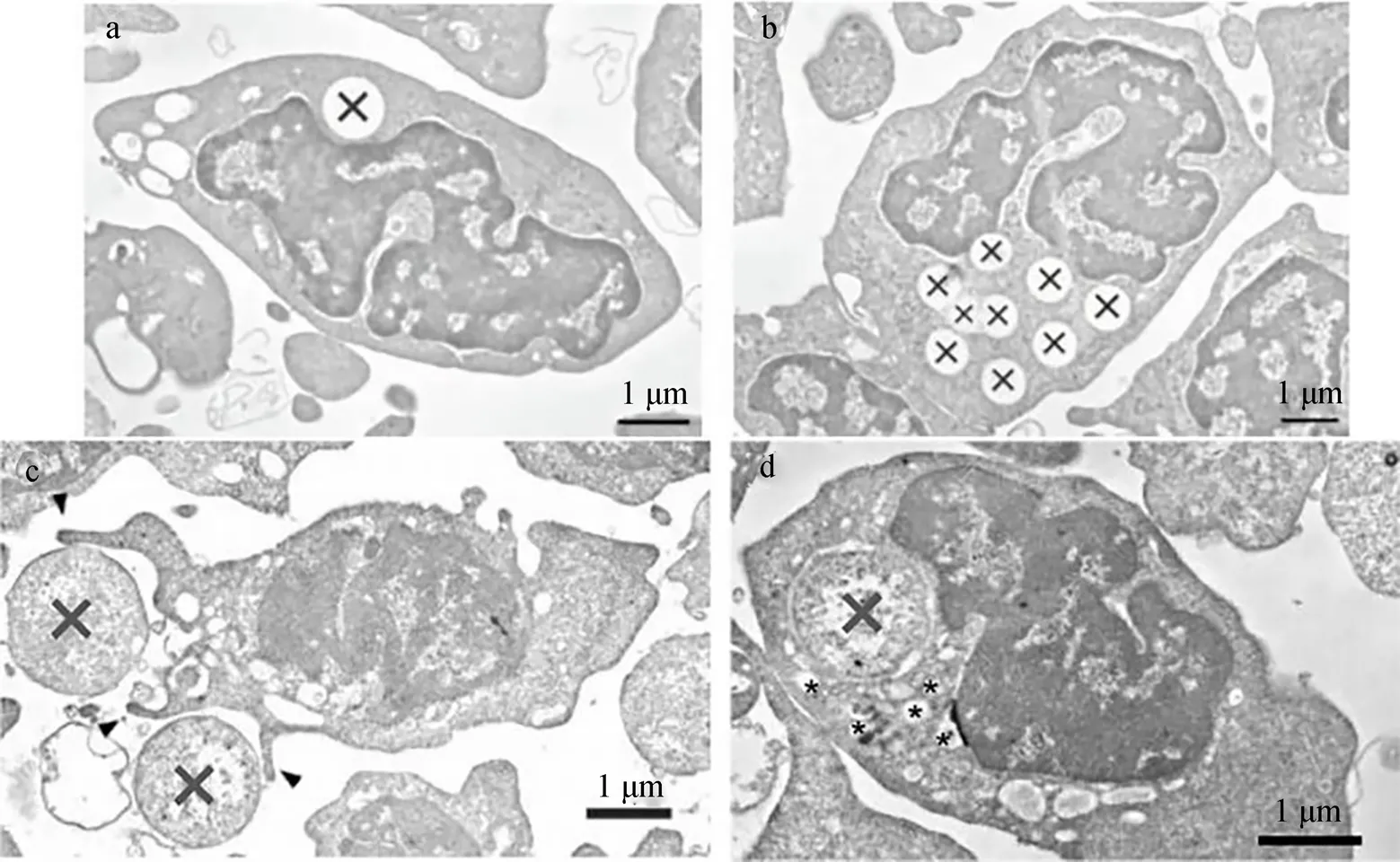
图4 鲤鱼血栓细胞吞噬荧光微球和细菌
a.b. 血栓细胞吞噬荧光微球(X为被吞噬的微球); c. 血栓细胞通过延伸伪足吞噬细菌, 箭头所示伪足, X.细菌; d. 小囊泡围绕着被摄入的细菌, *. 囊泡, 标尺=1 μm[8]
a.b. Phagocytosis with fluorescent beads by thrombocytes (Ingested beads are indicated as X); c. Thrombocytes engulfed bacteria (X) via the extension of pseudopods (arrowhead); d. Small vesicles surrounding the internalized bacteria are indicated by asterisks (*), Bar=1 μm[8]
2.2 非特异性免疫反应
机体组织损伤后, 血小板/血栓细胞会参与机体的稳态以及血栓形成, 进而参与炎症反应。血小板/血栓细胞会在第一时间聚集于损伤处, 除了聚集和参与炎症反应, 血小板/血栓细胞表达和释放有利于伤口组织修复的蛋白质和物质[48]。不同动物的血栓细胞分泌不同的细胞因子参与炎症反应和伤口修复。鸟类血栓细胞产生一些生物活性物质, 即趋化因子(巨噬细胞炎症蛋白-1β和一氧化氮)以及炎症反应的媒介物质, 如诱导型一氧化氮合酶(iNOS)等; 鸡血栓细胞能够表达抗炎细胞因子(转化生长因子TGF和IL-10)和前炎症因子(IL-1β, IL-6, IL-8和 IL-12)以及利于伤口修复的细胞因子[3]。鲤鱼和虹鳟血栓细胞能表达IL-1β、iNOS、肿瘤坏死因子TNFα、TGFβ和一些趋化因子[7-8]。
在哺乳动物中, 血小板可以表达一些非特异性免疫相关分子, 如Toll样受体(Toll-like receptors, TLR)和抗原呈递相关基因。鲤鱼、大西洋鳕鱼()、三刺鱼()和虎河豚()的血栓细胞可以表达辅助分子(TLR4 interactor with leucine-rich repeats, TRIL)[9], 这表明鱼类血栓细胞会像其他白细胞一样, 在非特异性免疫反应中直接发挥作用。
2.3 特异性免疫反应
抗原呈递是激活特异性免疫反应过程中的最后一个步骤, 主要经历: (1) 摄取抗原; (2) 将抗原呈递给特异性的胞内组分; (3) 将抗原肽释放到抗原结合位点-主要组织性相容性复合体Ⅱ分子(MHC II)上; (4) 激活CD4辅助性T细胞。鱼类的单核/巨噬细胞、树突状细胞和B淋巴细胞除了具有直接抗菌作用外, 还表达Ⅱ型组织相容性复合体, 且具有抗原处理和呈递的分子机制。低等脊椎动物血栓细胞表面具有MHC Ⅱ分子, 如鸟类[49]、虹鳟和鲤鱼[7-8, 10-11]的血栓细胞能够表达MHC Ⅱ分子mRNA, 参与抗原呈递过程。此外, 吞噬异物的血栓细胞以及具有抗原承载力的血栓细胞在鲤鱼的头肾和脾脏中都有所发现, 也说明血栓细胞具有运载抗原到淋巴组织并进入特异性免疫阶段的能力[8]。以上研究仅局限于定量PCR的方法, 鉴定血栓细胞中免疫相关基因的表达水平变化, 缺少不同层次(基因水平、蛋白水平、细胞水平)的科学证据来论证血栓细胞的免疫功能。目前, 鱼类血栓细胞非特异性和特异性免疫反应的分子水平研究与哺乳动物血小板还存在很大的差距。
3 血栓细胞的未来研究方向
鱼类血栓细胞的免疫功能研究具有较大的发展空间。目前, 鱼类血栓细胞的研究主要集中于鱼类血栓细胞特性研究, 这将仍是鱼类血栓细胞研究的方向。此外, 鱼类血栓细胞特性是否和人类血小板类似, 与自身因子(年龄)等因素有关[28], 仍需深入研究。因此, 鱼类血栓细胞特性与外界环境因素(栖息地、溶氧、pH等)以及内部因子(种类、年龄、性别、生理阶段等)的关系将会是研究的重点。血栓细胞表面特异性标记物及受体[49-52]发掘, 可以研发血栓细胞的特异性单克隆抗体, 如虹鳟[7]、斑马鱼[53]、鲤鱼[54]和牙鲆[55], 结合免疫磁珠、流式细胞术等方法, 提高鱼类血栓细胞的分离纯化效率[56-57]。利用分离纯化的血栓细胞, 在前期研究的基础上[7-12], 研究血栓细胞与其他免疫细胞的相互作用, 比如草鱼血栓细胞与白细胞的作用[12], 血小板与树突状细胞的相互作用[58]; 借鉴血小板[59]和鸡血栓细胞[60]的研究方法, 开展鱼类血栓细胞的组学研究和免疫调控网络分析, 将会是未来研究的热点。此外, 树蛙()血栓细胞系已经建立[5], 能否建立鱼类血栓细胞的细胞系, 开展胞外囊泡[61]等热点问题研究, 仍需要进一步探究。
[1] Giulio B. Rights and permissions[J]. Nature 157: 331- 332.
[2] Michelson A D, Coller B S. Platelets[M]. Amsterdam: Elsevier, 2007: 3-22.
[3] Ferdous F. The avian thrombocyte is a specialized immune cell[D]. US: Clemson University, 2014.
[4] David P, Carmen S. Morphologic and cytochemical characteristics of blood cells from the European Pond Turtle () and the Mediterranean Pond Turtle ()[J]. Journal of Herpetological Medicine and Surgery, 2009, 19(4): 119-127.
[5] Sugimoto K. Establishment of a sticky, large, oval-shaped thrombocyte cell line from tree frog as an ancestor of mammalian megakaryocytes[J]. Springerplus, 2015, 4: 447.
[6] Ferdous F, Scott T R. A comparative examination of thrombocyte/platelet immunity[J]. Immunology Letters, 2015, 163: 32-39.
[7] Köllner B, Fischer U, Rombout J H W M, et al. Potential involvement of rainbow trout thrombocytes in immune functions: a study using a panel of monoclonal antibodies and RT-PCR[J]. Developmental and Comparative Immunology, 2004, 28: 1049-1062.
[8] Nagasawa T, Nakayasu C, Rieger A M, et al. Phagocytosis by thrombocytes is a conserved innate immune mechanism in lower vertebrates[J]. Front Immunology, 2014, 5: 445.
[9] Pietretti D, Spaink H P, Falco A, et al. Accessory molecules for Toll-like receptors in teleost fish. Identification of TLR4 interactor with leucine-rich repeats (TRIL)[J]. Molecular Immunology, 2013, 56: 745-756.
[10] Fink I R, Ribeiro C M S, Forlenza M, et al. Immune- relevant thrombocytes of common carp undergo parasite-induced nitricoxide-mediated apoptosis[J]. Developmental and Comparative Immunology, 2015, 50: 146-154.
[11] Jaros J, Korytar T, Huong D T, et al. Rainbow trout () thrombocytes are involved in MHC II dependent antigen presentation[J]. Fish and Shellfish Immunology, 2013, 34: 1635-1691.
[12] Nagasawa T, Somamoto T, Nakao M. Carp thrombocyte phagocytosis requires activation factors secreted from other leukocytes[J]. Developmental and Comparative Immunology, 2015, 52: 107-111.
[13] 袁仕取, 张永安, 姚卫建, 等. 鳜鱼外周血细胞显微和亚显微结构的观察[J]. 水生生物学报, 1998, 22(1): 39-47. Yuan Shiqu, Zhang Yongan, Yao Weijian, et al. Micro and Ultra-structure of peripheral blood cells of the mandarin fish,(Basilewsky)[J]. Acta Hydrobiologica Sinica, 1998, 22(1): 39-47.
[14] 周永灿, 邢玉娜, 冯全英. 鱼类血细胞研究进展[J]. 海南大学学报(自然科学版), 2003, 21: 171-176. Zhou Yongcan, Xing Yuna, Feng Quanying. Research advance in the haemocytes of fishes[J].Natural Science Journal of Hainan University, 2003, 21: 171-176.
[15] 梅景良, 王寿昆, 祁保民, 等. 黑鲷外周血细胞的显微和超微结构观察[J]. 热带海洋学报, 2008, 27(2): 45-53. Mei Jingliang, Wang Shoukun, Qin Baomin, et al. Observation on microstructure and ultrastructure of peripheral blood cells of black sea bream ()[J]. Jounal of Tropical Oceanography, 2008, 27(2): 45-53.
[16] 胡玲玲, 李加儿, 区又君, 等. 条石鲷外周血细胞的显微结构[J]. 福建农林大学学报(自然科学版), 2009, 38(4): 384-387.Hu Lingling, Li Jiaer, OU Youjun, et al. Microstructure of peripheral blood cells in striped beak Perch[J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 2009, 38(4): 384-387.
[17] 金丽, 赵娜, 黄林, 等. 胭脂鱼外周血细胞的显微、超微结构与细胞化学观察[J]. 水生生物学报, 2011, 35(3): 550-556. Jin Li, Zhao Na, Huang Lin, et al. Structure and cytochemical characteristics of blood cells in[J]. Acta Hydrobiologica Sinica, 2011, 35(3): 550-556.
[18] 黄小铭, 张耀光, 江星, 等. 长薄鳅外周血细胞的显微结构和细胞化学特征研究[J]. 四川动物, 2012, 31(1): 59-63. Huang Xiaoming, Zhang Yaoguang, Jiang Xing, et al. Microstructure and cytochemical characteristics of peripheral blood cells in[J]. Sichuan Journal of Zoology, 2012, 31(1): 59-63.
[19] 师尚丽, 曹伏君, 陈思, 等. 长鳍裸颊鲷外周血细胞的显微结构[J]. 海洋渔业, 2014, 36(5): 418-423. Shi Shangli, Cao Fujun, Chen Si, et al. Microstructure of peripheral blood cells in[J]. Marine Fisheries, 2014, 36(5): 418-423.
[20] 毛海涛, 孙敬锋, 王庆奎, 等. 点带石斑鱼血液指标、血细胞分类与发生的观察[J]. 水产科学, 2015, 34(4): 214-219. Mao Haitao, Sun Jingfeng, Wang Qingkui, et al. Hematological indices, classification and hematopoiesis of blood cells in Malabar group[J]. Fisheries Science, 2015, 34(4): 214-219.
[21] 铁槐茂, 石丹, 银龙, 等. 瓦氏黄颡鱼血液生理指标与外周血细胞组成及显微结构[J]. 四川农业大学学报, 2015, 3(1): 78-83. Tie Huaimao, Shi Dan, Yin Long, et al. Blood physiological indexes, constitute and microstructure of peripheral blood cells in[J]. Journal of Sichuan Agriculture University, 2015, 3(1): 78-83.
[22] 张艳珍, 王彦鹏, 危起伟, 等. 中华鲟外周血细胞组成及形态观察[J]. 水生生物学报, 2018, 42(2): 323-332. Zhang Yanzhen, Wang Yanpeng, Wei Qiwei, et al. Composition and morphology of peripheral blood cells of[J]. Acta Hydrobiologica Sinica, 2018, 42(2): 323-332.
[23] Grant K R. Fish hematology and associated disorders[J]. Clinics in Laboratory Medicine, 2015, 35(3): 681-701.
[24] Jagadeeswaran J P, Sheehan F E, Craig, et al. Identification and characterization of zebrafish thrombocytes[J]. British Journal of Haematology, 1999, 107(4): 731-738.
[25] Tavares-Dias M, Ruas de M F. Leukocyte and thrombocyte reference values for channel catfish (Raf), with an assessment of morphologic, cytochemical, and ultrastructural features[J]. Veterinary Clinical Pathology, 2007, 36(1): 49-54.
[26] Silva W F, Egami M I, Santos A A, et al. Cytochemical, immunocytochemical and ultrastructural observations on leukocytes and thrombocytes of fat snook ()[J]. Fish and Shellfish Immunology, 2011, 31: 571-577.
[27] Azevedo C, Matos P, Rocha S, et al. Ultrastructure of novel thrombocytes in the dog snapper[J]. Journal of Fish Biology, 2014, 84: 865-871.
[28] Tian Jucheng, Liu Hanghang, Cui Xiao, et al. Investigating the effect of age on platelet ultrastructure using transmission electron microscopy[J]. International Wound Journal, 2019, 1-7.
[29] Shigdar S, Harford A, Ward A C. Cytochemical characterisation of the leucocytes and thrombocytes from Murray cod (, Mitchell)[J]. Fish and Shellfish Immunology, 2009, 26: 731-736.
[30] Bianchi M, Jerônimo G, Pádua S, et al. The hematological profile of farmed: Reference intervals, cell morphology and cytochemistry[J]. Veterinarski Arhiv, 2014, 84: 677-690.
[31] Lewis J C, Maldonado J E, Mann K G. Phagocytosis in human platelets: localization of acid phosphatase positive phagosomes following latex uptake[J]. Blood, 1976, 47(5): 833.
[32] 唐银. 养殖花斑裸鲤外周血液生理指标及血细胞形态参数的研究[D]. 成都: 四川农业大学, 2015. Tang Yin. Study on the Characterization of hematological parameters and morphology parameters of the peripheral blood cells of piebald naked carp ()[D]. Chengdu: Sichuan Agriculture University, 2015.
[33] Passantino L, Cianciotta A, Patruno R, et al. Do fish thrombocytes play an immunological role?Their cytoenzymatic profiles and function during an accidental piscine candidiasis in aquarium[J]. Immunopharmacology and Immunotoxicology, 2005, 27(2): 345-356.
[34] Rough K M, Nowak B F, Reuter R E. Haematology and leukocyte morphology of wild caught[J]. Journal of Fish Biology, 2005, 66: 1649-1659.
[35] Tavares-Dias M, Moraes F. Morphological, cytochemical, and ultrastructural study of thrombocytes and leukocytes in neotropical fish,Valenciennes, 1850 (Characidae, Bryconinae)[J]. Journal of Submicroscopic Cytology and Pathology, 2006, 38: 209-215.
[36] Prasad G, Charles S. Haematology and leucocyte enzyme cytochemistry of a threatened yellow catfish(Gunther 1864)[J]. Fish Physiology and Biochemistry, 2010, 36: 435-443.
[37] 赵柳兰, 陈侨兰, 杨淞, 等.“优鲈1号”大口黑鲈血细胞的形态特征及吞噬功能的研究[J]. 南方水产科学, 2016, 12(3): 43-50.Zhao Liulan, Chen Qiaolan, Yang Song, et al. Morphological characterization and phagocytosis of blood cells in “Youlu No.1” largemouth bass ()[J]. South China Fisheries Science, 2016, 12(3): 43-50.
[38] Ueda I K, Egami M I, Sasso W S, et al. Cytochemical aspects of the peripheral blood cells of Oreochromis (. Linnaeus, 1758) (Cichlidae, Teleostei)-Part Ⅱ[J]. Brazilian Journal of Veterinary Research Animal Science, 2001, 38: 273-277.
[39] Meenakshi B, Sidney W W. The ins and outs of endocytic trafficking in platelet functions[J]. Current Opinion in Hematology, 2017, 24(5): 467-474.
[40] Kordon A O, Abdelhamed H, Ahmed H, et al. Phagocytic and bactericidal properties of Channel catfish peritoneal macrophages exposed to live attenuated vaccine and wild-type strains[J]. Frontiers in Microbiology, 2018, 8: 2638.
[41] Øverland H S, Pettersen E F, Rønneseth A, et al. Phagocytosis by B-cells and neutrophils in Atlantic salmon (L.) and Atlantic cod (L.)[J]. Fish and Shellfish Immunology, 2010, 28: 193-204.
[42] Esteban M Á, Cuesta A, Chaves-Pozo E, et al. Implications of the new cells involved[J]. Biology, 2015, 4: 907-922.
[43] Ronneseth D B, Ghebretnsae H I, Wergeland G T, et al. Functional characterization of IgM+ B cells and adaptive immunity in lumpfish (L.)[J]. Developmental and Comparative Immunology, 2015, 52: 132-143.
[44] Zhang X J, Wang P, Zhang N, et al. B cell functions can be modulated by antimicrobial peptides in Rainbow trout: novel insights into the innate nature of B cells in Fish[J]. Frontiers in Immunology, 2017, 8: 388.
[45] Yang S, Tang X Q, Sheng X Z, et al. Development of monoclonal antibodies against IgM of half-smooth tongue sole () and analysis of phagocytosis of fluorescence microspheres by mIgM+ lymphocytes[J]. Fish and Shellfish Immunology, 2017, 66: 280-288.
[46] Stosik M, Deptuła W, Travnicek M, et al. Phagocytic and bactericidal activity of blood thrombocytes in carps ()[J]. Veterinarni Medicina, 2002, 47: 21-25.
[47] Burrows A S, Fletcher T C, Manning M J. Haematology of the turbot,(L): ultrastructural, cytochemical and morphological properties of peripheral blood leucocytes[J]. Journal of Applied Ichthyology, 2001, 17(2): 77-84.
[48] Koupenova M, Clancy L, Corkrey H A, et al. Circulating platelets as mediators of immunity, inflammation, and thrombosis[J]. Circulation Research, 2018, 122: 337-351.
[49] Paul M S, Paolucci S, Barjesteh N, et al. Characterization of chicken thrombocyte responses to Toll-Like receptor ligands[J]. Plos One, 2012, 7(1): e43381.
[50] Ohashi K, Takizawa F, Tokumaru N, et al. A molecule in teleost fish, related with human MHC-encoded G6F, has a cytoplasmic tail with ITAM and marks the surface of thrombocytes and in some fishes also of erythrocytes[J]. Immunogenetics, 2010, 62: 543-559.
[51] Hughes C E, Radhakrishnan U P, Lordkipanidze M, et al. G6f-Like is an ITAM-containing collagen receptor in thrombocytes[J]. Plos One, 2012, 7(12): e52622.
[52] Carrillo M, Kim S, Rajpurohit S K, et al. Zebrafish von Willebrand factor[J]. Blood Cells, Molecules, and Diseases, 2010, 45(4): 326-333.
[53] Ma D, Zhang J, Lin H F, et al. The identification and characterization of zebrafish hematopoietic stem cells[J]. Blood, 2011, 118(2): 289-297.
[54] Nakayasu C, Yoshitomi T, Oyamatsu T, et al. Separation of carp (L.) thrombocytes by using a monoclonal antibody, and their aggregation by collagen[J]. Veterinary Immunology and Immunopathology, 1997, 57(3-4): 337-346.
[55] Matsuyama T, Nakayasu C, Sano M. Immunocytochemical studies of the ontogeny of peripheral blood leucocyte subpopulations in Japanese flounder ()[J]. Fish and Shellfish Immunology, 2010, 29(2): 362-365.
[56] Kulkarni V, Kim S, Zafreen L, et al. Separation of young and mature thrombocytes by a novel immuno- selection method[J]. Blood Cells, Molecules, and Diseases, 2012, 48: 183-187.
[57] Lulijwa R, Alfaro A C, Merien F, et al. Advances in salmonid fish immunology: A review of methods and techniques for lymphoid tissue and peripheral blood leucocyte isolation and application[J]. Fish and Shellfish Immunology, 2019, 95: 44-80.
[58] Sharmeen N, Wuescher L M, Worth R G. Platelets enhance dendritic cell responses againstthrough CD40-CD40L[J]. Infection and Immunity, 2018, 86(9): e00186.
[59] 刘丹. 趋化因子CCL2对血小板功能的调控及其机制研究[D]. 沈阳: 第三军医大学学报, 2015. Liu Dan. Roles and mechanisms of chemokine CCL2 in platelet functions[D]. Shenyang: Journal of Third Military Medical University, 2015.
[60] Ferdous F, Saski C, Bridges W, et al. Transcriptome profile of the chicken thrombocyte: New implications as an advanced immune effector cell[J]. PloS One, 2016, 11(10): e0163890.
[61] Hill A. Methods in Molecular Biology[M]. New York: Humana Press, 2017.
Current research on the immune function of thrombocytes in fish
GAO Ying-li1, 2, 3, HAN Gao-shang1, YAO Hai-jing1, CHEN Xiang-ning1, 2, 3, ZHOU Qian-qing1
(1. College of Marine Life and Fisheries, Jiangsu Oean University, Lianyungang, 222000, China; 2. Jiangsu Key Laboratory of Marine Biotechnology, Lianyungang 222000, China; 3. Jiangsu Key Laboratory of Marine Bioresources and Environment, Lianyungang 222000, China)
The research on blood cells is very crucial to the research on fish. Research is basically focused on the immune functions of lymphocytes, macrophages, and neutrophils. There is limited focus on thrombocytes. Platelets in mammals and thrombocytes in lower vertebrates are implicated in thrombotic and hemostatic processes. The thrombocytes are regarded as special immune cells that play important roles in the immune response of fish based on current reports. This paper systemically reviews the immune-associated morphological features, immune functions, and future research perspectives on thrombocytes, with the aim of providing basic insight into the multi- functional nature of thrombocytes.
Morphology; immune function; thrombocyte; fish
Nov. 6, 2019
Q-1
A
1000-3096(2021)01-0120-09
10.11759/hykx20191106001
2019-11-06;
2020-05-25
江苏省自然科学基金青年基金项目(BK20170450); 江苏省省政策引导类计划-苏北科技专项项目(SZ-LYG2017020); 江苏省高等学校自然科学研究面上项目(17KJB240001); 江苏省生物技术重点实验室开放基金项目(HS2017003)和江苏省海洋资源与环境重点实验室开放基金项目(CXKT20180112); 江苏省大学生创新创业项目(SY201811641105003)
[Natural Science Foundation of Jiangsu Province, No. BK20170450; Policy Guidance Program of Jiangsu Province, No. SZ-LYG2017020; Natural Science Foundation of the Jiangsu Higher Education Institutions of China, No. 17KJB240001; The Open Research Fund of Jiangsu Key Laboratory of Marine Biotechnology, No.HS2017003; The Open Research Fund of Jiangsu Key Laboratory of Marine Bioresources and Environment, No. CXKT20180112; Student Innovation and Entrepreneurship Program of Jiangsu Province, No. SY201811641105003]
高迎莉(1988- ), 女, 江苏徐州人, 讲师, 博士, 主要从事水产动物病害与免疫学研究, E-mail: yingligao0127@126.com
(本文编辑: 谭雪静)
