冰川红叶小檗和紫叶小檗抗寒性对比分析
2021-02-03李昕蔓苏安然李玉言金卓颖冀伟王进茂
李昕蔓,苏安然,李玉言,金卓颖,冀伟,王进茂
(1 河北农业大学 林学院,河北 保定 071000;2 石家庄市藁城区绿都市政园林工程有限公司,河北 石家庄 052160)
植物的抗寒性是指植物忍受、抵御并适应低温的能力[1]。温度是限制植物生长发育及分布范围的重要生态因子[2]。低温会影响植物的生长发育,严重时会导致整个植株死亡,所以研究植物的抗寒性对指导农业生产及寻找其适种区域具有重要的指导意义[3]。抗寒性强的植物应具备:体内生物膜的正常液晶相不会发生相变[4];为防止冰晶对膜的破坏,必须要避免细胞内水分结冰;必须要具备抗冰冻脱水的性能,但不同的植物具有不同的抗性[5]。植物抗寒性的评价方法很多,包括田间调查法、解剖结构观察法、生理生化测定法、电阻抗图谱法、电导法、差热分解法、叶绿素荧光法、褐变法、恢复生长法、全株冰冻测试法、干物质含量法等,其中应用最为广泛的是电导法。随着现代科技的不断进步,用电阻抗图谱法测定植物的抗寒性也得到了广泛的应用[6-8]。
冰川红叶小檗(Berberisthunbergii‘Bingchuanhongye’)是小檗科、小檗属彩叶红果落叶灌木,小枝多红褐色,枝条丛生,有沟槽。单叶互生,红色叶片簇生,倒卵形,全缘叶,光滑无毛。花黄色,花两性,花梗长。红色浆果,成串挂在红色枝条中间。花、果、叶具有极高的观赏价值、适应性强、抗寒、抗旱、抗病虫、耐瘠薄,获得了国家林业植物新品种保护权并通过了河北省林木良种审定,是园林绿化中不可多得的彩叶树种[9]。紫叶小檗(Berberisthunbergiivar.atropurpurea),又叫红叶小檗,是小檗科、小檗属的落叶灌木,幼枝淡红带绿色,老枝灰棕色或紫褐色且具条棱。叶菱状卵形、全缘,紫红色到鲜红色,叶背面带灰白色。4月开花,花黄色。果实椭圆形,果熟后艳红美丽,并且具有枝叶繁茂、适应性强、病虫害少、抗逆性强、观赏价值高等优良特性,是园林绿化的优良树种[10]。
为了研究冰川红叶小檗是否适合栽植在东北寒冷地区,以冰川红叶小檗、紫叶小檗为研究对照,探讨2种小檗在不同低温处理下相对电导率、电阻抗参数值以及萌芽率的变化,并根据相对电导率、电阻抗参数值配合Logistic方程拟合其半致死温度,运用电导法、电阻抗图谱法和恢复生长法,综合评价其抗寒性,为寻找其适种的栽培区域及推广应用提供科学的理论依据。
1 材料与方法
1.1 试验地概况
试验地位于河北省石家庄市藁城区南董镇南董村的绿都市政园林工程有限公司苗圃基地。该地属于暖温带大陆性季风气候,夏季高温多雨,冬季寒冷干燥,季风性显著,夏秋常受热带气旋影响。年平均气温12.8 ℃,年极端高温43.2 ℃,年极端低温-23.4 ℃。1月份最冷,月平均气温-3.5 ℃;7月份最热,月平均气温26.4 ℃。年平均降水量445.8 mm,年最大降水量1 007.1 mm,年最小降水量224.6 mm。降水主要集中在夏季6至9月,其中以7、8月降水量最多,年日照时数2 513.3 h,无霜期203.5 d。
1.2 试验材料
供试材料为冰川红叶小檗和紫叶小檗的当年生枝条,均采自绿都市政园林工程有限公司苗圃基地。试验苗木为2 a生嫩枝扦插苗,2个品种隔行栽植,株行距为30 cm×30 cm,常规管理,苗木在生长季节不施肥追肥,在干旱情况下浇水。
1.3 样品处理
2018年12月上旬和2019年3月上旬分别随机采集生长良好、无病虫害、无破损、粗细均匀一致的当年生枝条带回实验室,用去离子水冲洗3次,之后用吸水纸擦干,然后剪成80段5 cm长的枝条,充分混匀后随机选择11段枝条为1份置于塑封袋中(电导法设定3次重复,电阻抗图谱法设定8次重复),一共7份,利用冰箱设置7个温度(4 ℃、-10 ℃、-20 ℃、-30 ℃、-35 ℃、-40 ℃、-50 ℃)处理,样品处理好以后,统一放入低温冰柜中,以4 ℃/h的速率降温,到达设定温度后保持24 h,取出该温度处理材料,以4 ℃/h的速率升温至 4 ℃,解冻24 h,测定各项抗寒性指标;其他材料继续以4 ℃/h的速率降温,到达下一设定低温处理温度,取出该温度处理材料,采用相同的方法升温并测定抗寒指标,依次进行低温处理,直到最后1个处理(-50 ℃)。
1.4 试验方法
1.4.1 电导法(EL)测定枝条抗寒性 在枝条中部避开芽眼,剪取1 cm粗细均匀一致的茎段,充分混匀后随机挑选3段,再把每段平均分成4份,放到10 mL的试管中,加入10 mL去离子水,用保鲜膜把试管口封紧,THZ-92A型台式恒温振荡仪振荡24 h,然后利用DDS-307A型电导率仪测定其初电导率,测定完成后再用封口膜将试管口封紧,沸水浴30 min,再振荡24 h后测定其终电导率,根据初电导率和终电导率计算相对电导率。
1.4.2 电阻抗图谱法(EIS)测定枝条抗寒性 样品的处理方法同电导法。低温处理后,避开芽眼剪取枝条中部、粗细均匀一致的1.5 cm长茎段,充分混匀后随机挑选8段,用测厚仪测定每个茎段的直径,用阻抗分析仪测定每个茎段(80~1 MHz)的电阻抗值。求得的电阻抗值与其处理温度作图,并拟合Logistic曲线函数。
1.4.3 恢复生长法测定枝条抗寒性 随机选择2个品种外围中部的生长良好、无病虫害、无破损、粗细均匀一致、芽体饱满、20 cm左右的当年生枝条,共20个枝条,充分混匀后,随机选择6个枝条为1份放入自封袋中,每个品种3次重复,放到冰箱里进行低温处理。低温处理设置3个(-10 ℃、-20 ℃、-40 ℃)温度梯度,以4 ℃/h的速率降温,到达设定温度后保持24 h,再以4 ℃/h的速率升温至4 ℃,解冻24 h,然后放到人工气候室(温度27 ℃,相对湿度80%)进行水培,每天及时换水并记录萌芽数量,直到萌芽数量不再增加,统计萌芽数及总芽数,计算萌芽率。
1.5 数据处理
用Excel和SPSS 11.5对冰川红叶小檗和紫叶小檗的相对电导率、萌芽率进行方差分析和显著性分析。根据相对电导率、电阻抗参数值,用SPSS 11.5 软件拟合Logistic方程y(x)=a/(1+eb(c-x)) +
d,根据方程拟合半致死温度并计算半致死温度和萌芽率之间的相关性。
2 结果与分析
2.1 不同温度处理下2个小檗品种电导率
不同低温处理下2种小檗相对电导率的变化见表1。
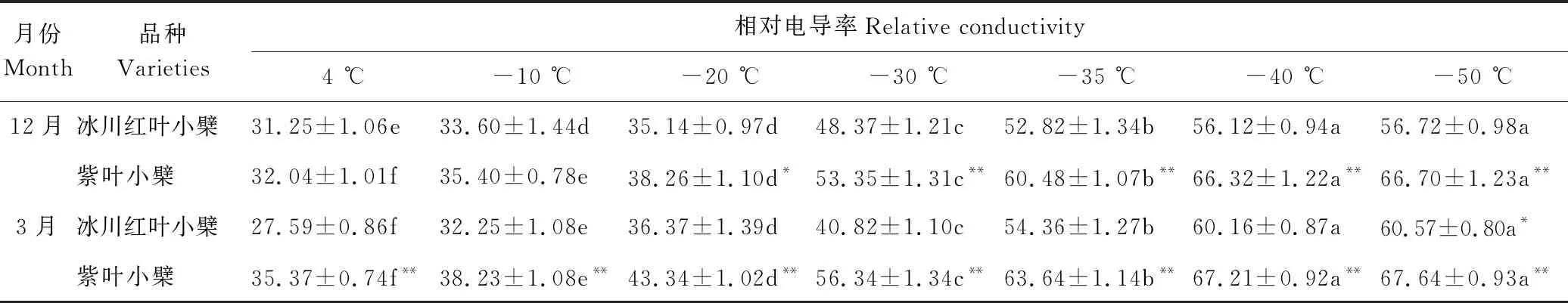
表1 不同低温处理下2种小檗相对电导率的变化Table 1 Changes of relative conductivity of two species of Berberis under different low temperature treatments %
由表1可知,在-50 ℃~4 ℃处理温度范围内,2种小檗随着处理温度逐渐降低,其相对电导率逐渐增加,但增加幅度有所不同。当处理温度在-20 ℃~4 ℃时,相对电导率增加较为缓慢;当处理温度在-40 ℃~-20 ℃时,相对电导率急剧增加;当处理温度为-40 ℃以下时,相对电导率又趋于平稳。12月份,当处理温度为4 ℃和-10 ℃时,2种小檗的相对电导率不存在显著差异;当处理温度为-20 ℃和-30 ℃时,2种小檗的相对电导率存在显著差异;当处理温度为-35 ℃、-40 ℃、-50 ℃时,2种小檗的相对电导率存在极显著差异。3月份2种小檗在不同低温处理下的相对电导率均存在极显著差异。
2.2 不同低温处理下2个小檗品种半致死温度
不同抗寒性测定方法2种小檗的半致死温度见表2。
由表2可知,12月份冰川红叶小檗当年生枝条用相对电导率,胞外电阻率(re)及低频电阻率(r1)拟合的半致死温度分别为-34.84 ℃、-31.46 ℃和-29.47 ℃,紫叶小檗拟合的半致死温度分别为-28.37 ℃、-27.12 ℃和-26.07 ℃。3月份冰川红叶小檗当年生枝条用相对电导率,胞外电阻率(re)及低频电阻率(r1)拟合的半致死温度分别为-31.94 ℃、-29.94 ℃和-27.98 ℃,紫叶小檗拟合的半致死温度分别为-26.84 ℃、-26.08 ℃和-25.43 ℃。用电导法拟合的半致死温度和用胞外电阻率(re)、低频电阻率(r1)拟合的半致死温度的相关系数分别为0.997、0.999,达到了极显著相关水平。12月和3月用电导法拟合的半致死温度比用胞外电阻率(re)、低频电阻率(r1)拟合的半致死温度低。12月和3月用2种抗寒性测定方法拟合的冰川红叶小檗的半致死温度均低于紫叶小檗的半致死温度,说明冰川红叶小檗的抗寒性高于紫叶小檗。12月用2种抗寒性测定方法拟合的半致死温度均低于3月用2种抗寒性测定方法拟合的半致死温度,说明2种小檗12月的抗寒性高于3月份的抗寒性。
2.3 不同低温处理下2个小檗品种萌芽率
不同低温处理下2种小檗萌芽率的变化见表3。
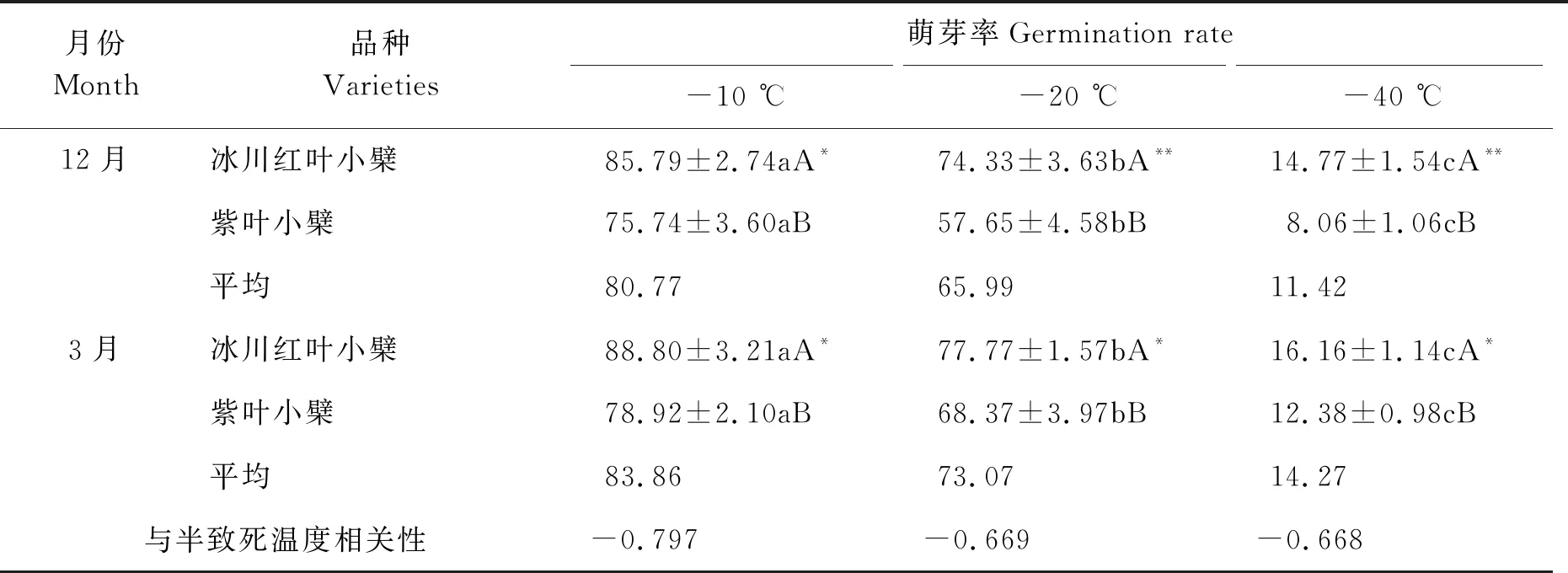
表3 不同低温处理下2种小檗萌芽率对比
由表3可知,12月份不同低温处理下的当年生枝条冰川红叶小檗的萌芽率分别为85.79%、74.33%和14.77%;紫叶小檗的萌芽率分别为75.74%、57.65%和8.06%,3月份不同低温处理下的当年生枝条冰川红叶小檗的萌芽率分别为88.80%、77.77%和16.16%,紫叶小檗的萌芽率分别为78.92%、68.37%和12.38%。随着处理温度的降低,冰川红叶和紫叶小檗的萌芽率逐渐降低,但变化幅度有所不同。12月份,在-10 ℃、-20 ℃、-40 ℃低温胁迫条件下,冰川红叶和紫叶小檗的萌芽率存在显著或极显著差异。3月份,在-10 ℃、-20 ℃、-40 ℃低温胁迫条件下,冰川红叶和紫叶小檗的萌芽率存在显著差异。3月份冰川红叶小檗和紫叶小檗的萌芽率普遍高于其12月份的萌芽率。冰川红叶小檗和紫叶小檗半致死温度的平均值与萌芽率之间的相关系数分别为-0.797、-0.669、-0.668,呈负相关关系,但均未达到显著或极显著相关水平。
3 结论与讨论
用3种抗寒性测定方法对冰川红叶小檗和紫叶小檗的抗寒性进行测定和对比分析,得到如下结论:(1)冰川红叶小檗的抗寒性高于紫叶小檗。(2)2个小檗品种12月的抗寒性高于3月的抗寒性。(3)用电导法和电阻抗图谱法测定的抗寒性一致,但用电导法拟合的半致死温度略低于电阻抗图谱法拟合的半致死温度。
本研究用3种不同的抗寒性测定方法探究2个小檗品种抗寒性的差异以及不同抗寒性方法测定结果的相关性,试验结果表明,用相对电导率拟合的半致死温度[11]以及用胞内电阻率(r1)和胞外电阻率(re)拟合的半致死温度[12]的相关性达到了极显著相关,主要原因是因为植物在低温胁迫条件下,细胞膜会受到不同程度的伤害,使其透性增加,电解质大量外渗,进而导致其相对电导率增加,电阻抗值减小[13],这2种植物抗寒性测定方法的原理基本一致。
用电导法和电阻抗图谱法拟合的半致死温度[14]都是根据植物受到低温胁迫后电解质的外渗情况经过数理统计拟合出来的,而枝条受到低温胁迫之后的萌芽率更能真实准确地反映其抗寒性的差异。本研究结果显示,2种抗寒性测定方法拟合的半致死温度的平均值与不同低温处理下的萌芽率呈负相关关系,这与于立洋等对新疆野苹果抗寒性的研究结果一致[15],也侧面说明了综合这3种方法能够准确测定2种小檗的抗寒性。本试验结果显示,用电阻抗图谱法拟合的半致死温度比用电导法拟合的半致死温度相对偏高,其抗寒性普遍偏低,这一研究结果与张军、于立洋等对新疆野苹果抗寒性的研究结果一致[15-16],也与张振东等对枣树枝条抗寒性的研究结果一致[17]。但董军生等对杨树抗寒性的研究结果是用电阻抗图谱法拟合的半致死温度比用电导法拟合的半致死温度相对较低,抗寒性较高,与该研究结果相反[18]。以上说明本研究所使用方法仅能反应在设定的试验条件下不同品种的抗寒性差异,但测定的半致死温度不能反映实际低温。
电导率法和半致死温度所得的结果为3月份冰川红叶小檗和紫叶小檗的抗寒性低于12月份其抗寒性,而萌芽率得出结果则相反,其主要原因是3月份枝条开始萌动,各种生理活动已开始,其含水量高,所以3月份以相同低温方法处理的枝条要比12月份枝条的萌芽率高。
本研究只对2种小檗2个时期(初冬开始休眠、初春打破休眠)的当年生枝条做了抗寒性测定,所设处理温度的梯度也相对较大,在今后的试验中,我们要对小檗属植物的抗寒性进行更加全面的研究,对不同时期枝条设置更多的处理温度,对经过低温处理之后当年生枝条的丙二醛含量、可溶性糖、可溶性蛋白、游离脯氨酸、超氧化物歧化酶、过氧化物酶、过氧化氢酶等生理指标进行测定,探索小檗属不同处理温度及不同时期抗寒性的变化规律以及各生理指标与抗寒性之间的相关性,为筛选小檗属优良抗寒种质资源提供科学的理论依据以及寻找其适种区域提供科学的指导[19-20]。
