基于花器官特征的新疆杏品种亲缘关系及遗传多样性分析
2021-02-03王亚楠李雯雯周伟权樊国全章世奎王亚桐
王亚楠,李雯雯,周伟权,樊国全,章世奎,王亚桐,廖 康
(1.新疆农业大学特色果树研究中心,乌鲁木齐 830052,2.新疆农业科学院轮台果树资源圃,新疆轮台 841600)
0 引 言
【研究意义】杏(PrunusarmeniacaL.)为蔷薇科(Rosaceae)杏属(ArmeniacaScop)植物,是我国重要的经济生态型果树树种之一,其分布广泛[1]。杏原产中国和中亚[2],起源中心(多样化中心或基因中心)在我国新疆,种质资源极为丰富[3-4]。在长期的人为驯化和生态适应过程中,杏在亚欧大陆各地迥异的地理气候条件下形成了中亚(Central Asian)、伊朗-高加索(Irano-Caucasian)、欧洲(European)、准噶尔-外伊犁(Dzhungar-Zailij)和华北(Northern China)等5个不同的生态地理群[5-6],其中中国新疆南部环塔里木盆地周边绿洲的库车、喀什及和田3个不同亚群为中亚生态群的重要组成部分[7],新疆杏栽培历史悠久,形成了诸多优质的品种。【前人研究进展】李明[8]采用ISSR、SSR和SRAP分子标记技术,对20个以新疆南疆杏为代表的中亚生态群杏品种的遗传多样性和遗传结构进行了研究,这些品种大多显示出生长期长、中小型果实、果实毛少或无毛等相似的形态性状,新疆杏品种的亲缘关系近;苑兆和等[9]利用AFLP荧光标记技术研究发现,我国新疆南疆栽培杏的AFLP扩增多态性为72.70%,与世界其它地区的杏群体相比有较高的遗传多样性;张艳敏[10]研究得出,新疆南疆栽培杏遗传多样性较丰富,其中库车的最丰富;刘娟[11]采用ISSR分子标记技术共扩增出485条条带,其中478条为多态性条带,得出栽培杏多态性比率为98.56%,供试资源中遗传多样性最丰富的是轮台采样点。上述研究者通过表型多样性和分子标记的方法,对我国新疆栽培杏亲缘关系和遗传多样性进行了研究,喀什亚群、和田亚群、库车亚群间栽培杏表型特征相似程度高,亲缘关系近,亚群间多态性比率高且遗传多样性丰富,亚群间存在高度一致性。【本研究切入点】花期是杏生长期中重要的阶段之一,花粉形态特征受植物基因型控制而受外界条件影响小,具有较强的遗传保守性,是探讨植物起源、演化及亲缘关系的重要特征之一[12-14],花粉形态学中的花粉外壁纹饰具有稳定的遗传性,植物的孢粉因其外壁结构坚固独特,有极强的遗传保守性和稳定性,果树也不例外[15]。研究杏品种花器官表型特征及花粉微观特性差异。【拟解决的关键问题】研究选取喀什亚群、和田亚群、库车亚群3个不同亚群间的杏种质作为供试材料,通过花器官指标测定和扫描电镜观察分析,分析比较供试样本的花器官及花粉微观特征,利用系统聚类的方法对新疆杏品种间亲缘关系进行分析,计算21个表型性状的变异系数、Simpson多样性指数、Shannon-weaver多样性指数,研究其遗传多样性,为其种质资源的评价、鉴定与利用提供参考依据。
1 材料与方法
1.1 材 料
样品采样地点地位于新疆农业科学院轮台果树资源圃(该资源圃位于E 84°14′,N 41°47′),其中喀什亚群14个品种,和田亚群13个品种,库车亚群14个品种,共计41个品种,供试品种所属亚群参考苑兆和[9]与刘娟[14]的分类方法。表1
1.2 方 法
1.2.1 花器官指标测定
参照《杏种质资源描述规范和数据标准》[15],于2019年4月6~8日分别采集各品种盛花期10朵花,观测花瓣形状、花瓣颜色、花萼颜色、花瓣数量、花冠直径、雄蕊数目、单花重、花瓣纵径、花瓣横径。对3种非数值性状进行赋值,分别为花瓣形状、花瓣颜色、花萼颜色。选取品种树体健壮,砧木及管理条件一致。表2

表1 供试栽培杏品种及编号Table 1 Cultivated apricot varieties and number

表2 新疆杏品种花器官非数值性状的赋值Table 2 Assignment of non-numerical characters of apricot varieties in Xinjiang almond flower organs
1.2.2 花粉微观特征观测
各品种分别收集气球期花蕾60个左右,剥出花药平铺于硫酸纸盒内阴干散粉,待花粉完全散出后,收集至1.5 mL离心管中,标明单株编号并封口,至于硅胶中密封干燥避光4℃保存。在SEM下观察花粉微观特征,在500~1 000倍下观察花粉集群,2 000倍下观察花粉萌发沟,2 500倍下观察花粉赤道面,3 500倍下观察花粉极面,20 000倍下观察花粉外壁纹饰。
使用Image J图像分析软件,分别测量花粉的极轴长、赤道轴长、脊宽、脊距、孔径、孔频等指标,通过极轴长和赤道轴长的比值计算极赤比,通过面积和穿孔数量计算孔频。分析各品种的花粉形状、极面观、赤面观、外壁纹饰类型以及NPC系统分类地位等指标。每个品种设置10个重复,将数值型数据求出平均值后进行聚类分析。
1.3 数据处理
使用Excel 2010处理基础数据,对测量性指标进行平均值的计算,使用SPSS 19.0软件进行标准值的计算,X±Y,X为同一品种10个重复的平均值,Y为同一品种10个重复的标准差。使用SPSS 19.0软件采用UPGMA聚类法对花器官特征和花粉微观形态特征分别进行聚类分析。采用DPS7.05数据处理软件计算表型性状的变异系数、Simpson多样性指数、Shannon-weaver多样性指数。共有21个指标。
变异系数(CV)计算公式为:
CV=σ/μ.
式中σ为41个供试样本中同一测量指标的标准差,μ为41个供试样本中同一测量指标的平均值。
Shannon-weaver多样性指数(H')计算公式为:
式中41表示研究供试样本总量,pi表示某一项指标第i个品种的值与这项指标数据总和的比值,ln表示自然对数e,e≈2.718 28。
Simpson多样性指数(D)计算公式为:
式中41表示研究供试样本总量,pi表示某一项指标第i个品种的值与这项指标数据总和的比值。
2 结果与分析
2.1 杏品种间亲缘关系
2.1.1 花器官性状特征
研究表明,多数品种花瓣为卵圆、圆形、椭圆3个不同形状,花瓣为白、浅粉红、深粉红3个不同的颜色;而在花萼颜色指标中除了和田亚群的克尔基胡安娜花萼颜色为紫绿色,其余的40个品种都为紫红、红褐2个颜色。在花瓣数指标中除了皮乃孜花瓣数大部分为5片,也存在6和7片花瓣的花,其余40个品种均为5片花瓣;麻木达西杏为花冠直径值最大值30.98 mm,古木杏和卡拉阿藏为花冠直径值最小值20.92 mm;雄蕊数量最多的黄其力干为35.92个,雄蕊数量最少的克孜西米西为23.02个;单花重最大值为粗黑叶杏0.18 g,单花重最小值品种有奎克皮曼、托乎提和晚熟杏3个,最小值为0.06 g;花瓣纵径最大值为大五月杏12.93 mm,最小值为晚熟杏7.14 mm,花瓣横径最大值为冬杏22.25 mm,最小值为古木杏和卡巴克西米西9.96 mm;花瓣纵横径比值最大为大五月杏1.04,比值最小为冬杏0.41。花冠直径、花瓣纵径、花瓣横径是显示差异的重要指标。表3,表4
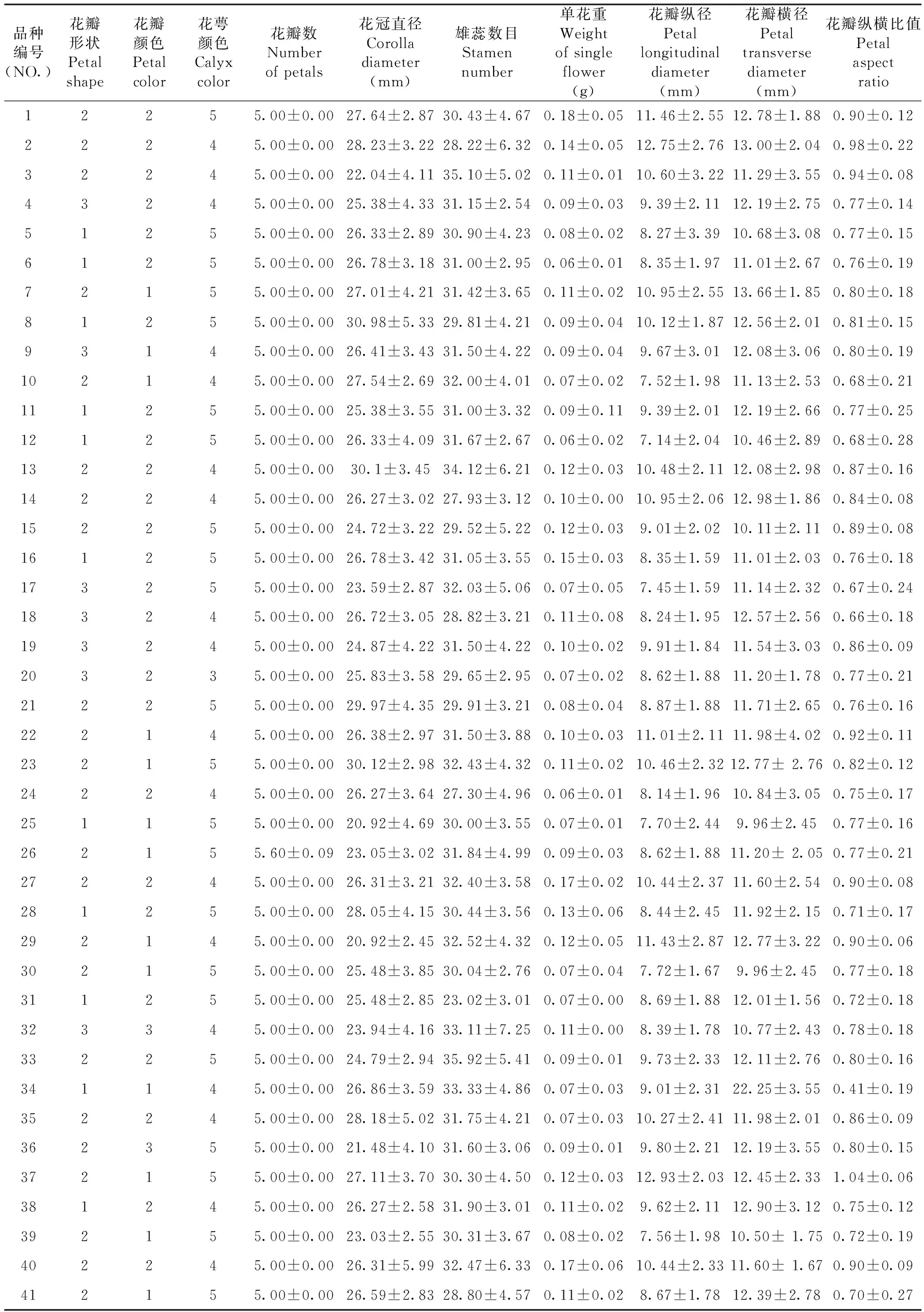
表3 41个新疆杏品种的花器官性状Table 3 Floral character of 41 apricot cultivars in Xinjiang

表4 41个新疆杏品种的花粉微观特征Table 4 Pollen morphology indexes of 41 apricot varieties in Xinjiang
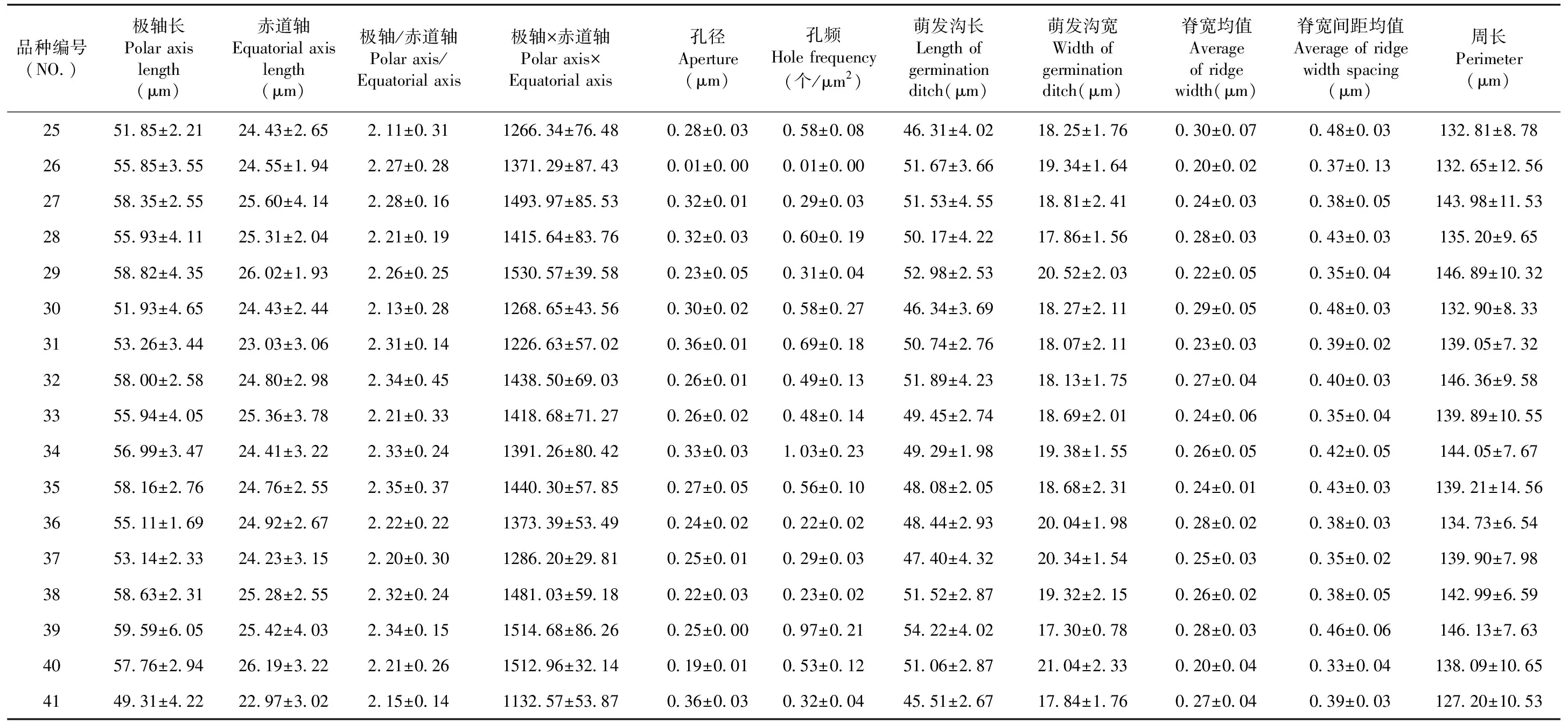
续表4 41个新疆杏品种的花粉微观特征Table 4 Pollen morphology indexes of 41 apricot varieties in Xinjiang
2.1.2 花粉微观形态特征
研究表明,41个品种的花粉粒辐射对称。根据埃尔特曼提出的NPC分类系统,供试品种均属于N3P4C5型,即3条沟都单环状等间距分布在赤道位置上。部分不同亚群品种花粉外部微观形态。
41个品种极赤比数值都大于2,形状都为超长球形;所有供试品种极轴值都大于赤道轴,赤面观均为窄椭圆形。供试样本的极轴长在50.26~60.28 μm,最大值为克孜佳娜丽,最小值为晚熟杏;赤道轴在22.65~26.13 μm,最大值为阿克牙格勒克,最小值为晚熟杏;极轴与赤道轴比值在2.09~2.46,最大值为克孜佳娜丽,最小值为白色买提;极轴与赤道轴乘积在1 132.57~1 530.57,最大值为卡拉阿藏,最小值为阿克达拉孜。
41个品种外壁纹饰都属于“条纹-穿孔”型,外壁纹饰的相似度极高。但在外壁纹饰的孔径、孔频、脊宽均值、脊宽间距均值这4项指标中略有不同,孔径值范围在0.01~0.44 μm,英吉沙杏的孔径值最大为0.44 μm,皮乃孜的孔径值最小为0.01 μm;孔频范围在0.01~0.69 个/μm,克孜西米西的孔频值最大为0.69 个/μm,皮乃孜的孔频值最小为0.01 个/μm;脊宽均值范围在0.20~0.34 μm,叶城黑叶杏的脊宽均值最大为0.34 μm,皮乃孜的脊宽均值最小为0.20 μm;脊宽间距在0.27~0.48,英吉沙杏的脊宽间距最大为0.48,晚熟杏的脊宽值最小为0.27。供试品种花粉微观形态特征差异较小,品种花粉微观形态特征相似度极高,品种间亲缘关系较近。图1
2.1.3 花器官表型及花粉微观特征聚类分析
研究表明,在遗传距离系数为15.0时供试的41个杏品种明显分为4大类:第I组有30个品种,分为A和B组2个小组,A组中包含和田亚群的木孜佳娜丽和阿克牙格勒克2个品种,喀什亚群的细黑叶杏,库车亚群的卡巴克西米西、大五月杏、克孜朗3个品种,B组中包含了和田亚群的图乃斯坦胡安娜、早大油杏、克孜佳娜丽、克尔基胡安娜、卡拉胡安娜、奎克皮曼、洛浦1号、皮乃孜8个品种;喀什亚群的色买提王132号、赛买提、中熟佳娜丽、叶城黑叶杏、托乎提、晚熟杏、英吉沙杏、贝新纳尔、胡安娜、白色买提这10个品种;库车亚群的苏陆克、洪待克、阿克达拉孜、卡拉阿藏、托乎提库都这5个品种。第II组包含来自和田亚群的洛浦洪待克、脆佳娜丽2个品种和来自库车亚群的黄其力干1个品种。第III组仅有来自库车亚群的克孜西米西1个品种。第IV组包含来自喀什亚群的粗黑叶杏、麻木达西杏、乔尔胖3个品种;来自和田亚群的古木杏1个品种;来自库车亚群的阿克阿依、冬杏、毛拉肖、佳娜丽4个品种。图2
供试的41个品种花粉微观特征的距离系数在0~25.0,第I组与第II组被分为1组,二者亲缘关系更近,而与第III组亲缘关系较远;为了能和花器官聚类在同一距离系数上形成对比,选择距离系数为15.0进行分析,41个品种明显聚为2大组:第I组有36个品种,将其分为A和B组,A组有13个品种,包含和田亚群的古木杏、脆佳娜丽2个品种;喀什亚群的乔尔胖、白色买提、贝新纳尔、英吉沙杏、色买提王132号、赛买提、大五月杏、胡安娜8个品种;库车亚群的卡巴克西米西、冬杏、托乎提库都3个品种;B组有23个品种,包含和田亚群的图乃斯坦胡安娜、克尔基胡安娜、洛浦洪待克、洛浦1号、奎克皮曼、阿克牙格勒克、早大油杏、木孜佳娜丽、皮乃孜、卡拉胡安娜、克孜佳娜丽11个品种;喀什亚群的中熟佳娜丽、麻木达西杏、托乎提、细黑叶杏4个品种;库车亚群的克孜朗、黄其力干、佳娜丽、洪待克、苏陆克、卡拉阿藏、毛拉肖、阿克阿依8个品种。第II组有5个品种,其中喀什亚群有叶城黑叶杏、粗黑叶杏、阿克达拉孜3个品种;库车亚群有克孜西米西、晚熟杏2个品种。图3
花粉的聚类中大部分品种是按照地域进行分组,而部分品种与地域划分所属亚群不吻合,如第II组中喀什亚群的叶城黑叶杏、粗黑叶杏、阿克达拉孜和库车亚群的克孜西米西、晚熟杏划分为1组。
2.2 新疆杏表型遗传多样性
研究表明,21个表型性状的变异系数幅度为3.07%~75.68%,孔频(75.68%)变异系数最大,花瓣形状(34.45%)、花瓣颜色(31.30%)、单花重(30.20%)、孔径(30.11%)这4个表性指标的变异系数较大,花瓣数(3.07%)变异系数最小;Shannon-weaver指数幅度为5.01~5.43,花瓣纵横比值Shannon-weaver指数最高,孔频Shannon-weaver指数最低,Simpson指数幅度为0.96~0.98,除了孔频的Simpson指数为0.96,其余40个品种的Simpson指数为都为0.97或者0.98。变异系数、Shannon-weaver指数和Simpson指数都是衡量种质资源遗传多样性的重要指标,变异系数表示品种多样性,值越高,品种多样性越高,遗传多样性越丰富[17],研究中41份供试样本间变异系数平均值为16.24%;Shannon-weaver指数表示群落多样性的高低,当其值大于3时,供试样本所属群落丰富度高,供试样本来自3个不同的亚群,且得出Shannon-weaver指数平均值为5.33>3,此供试样本所属类群丰富度较高;Simpson指数表示随机取样的2个供试样本不属于同一品种的概率,Simpson指数越接近1,供试样本间多样性程度越高,研究中Simpson指数平均值为0.97,很接近1,符合供试样本间多样性程度高的结论。供试新疆杏样本间多样性程度高,遗传多样性丰富。表5
3 讨 论
3.1 基于花器官表型及花粉微观特征的品种间亲缘关系
新疆杏的花器官性状差异主要表现在花冠直径、花瓣横径、雄蕊数目这3个指标,是分析新疆杏品种亲缘关系的重要参考指标。对南疆杏种质资源表型多样性的研究除了孙家正等[18]通过对南疆栽培杏各种化学成分组成的研究,在果肉香气成分、糖、酸组分及杏仁油脂肪酸组分上有报道。研究通过21个花器官指标测量和观察,得出供试样品花瓣形状大部分为卵圆、圆形、椭圆,花瓣颜色大部分为白、浅粉红、深粉红,花萼颜色大部分为紫绿、紫红、红褐的性状,花瓣数除了皮乃孜以外的40个品种都为5片花瓣,皮乃孜的花瓣数大部分为5片,但是也存在6和7片的花,这与此品种自身花器官特性和演化过程中基因突变有关,供试41个栽培杏品种表型性状相似性较高,其存在一定的遗传共性。
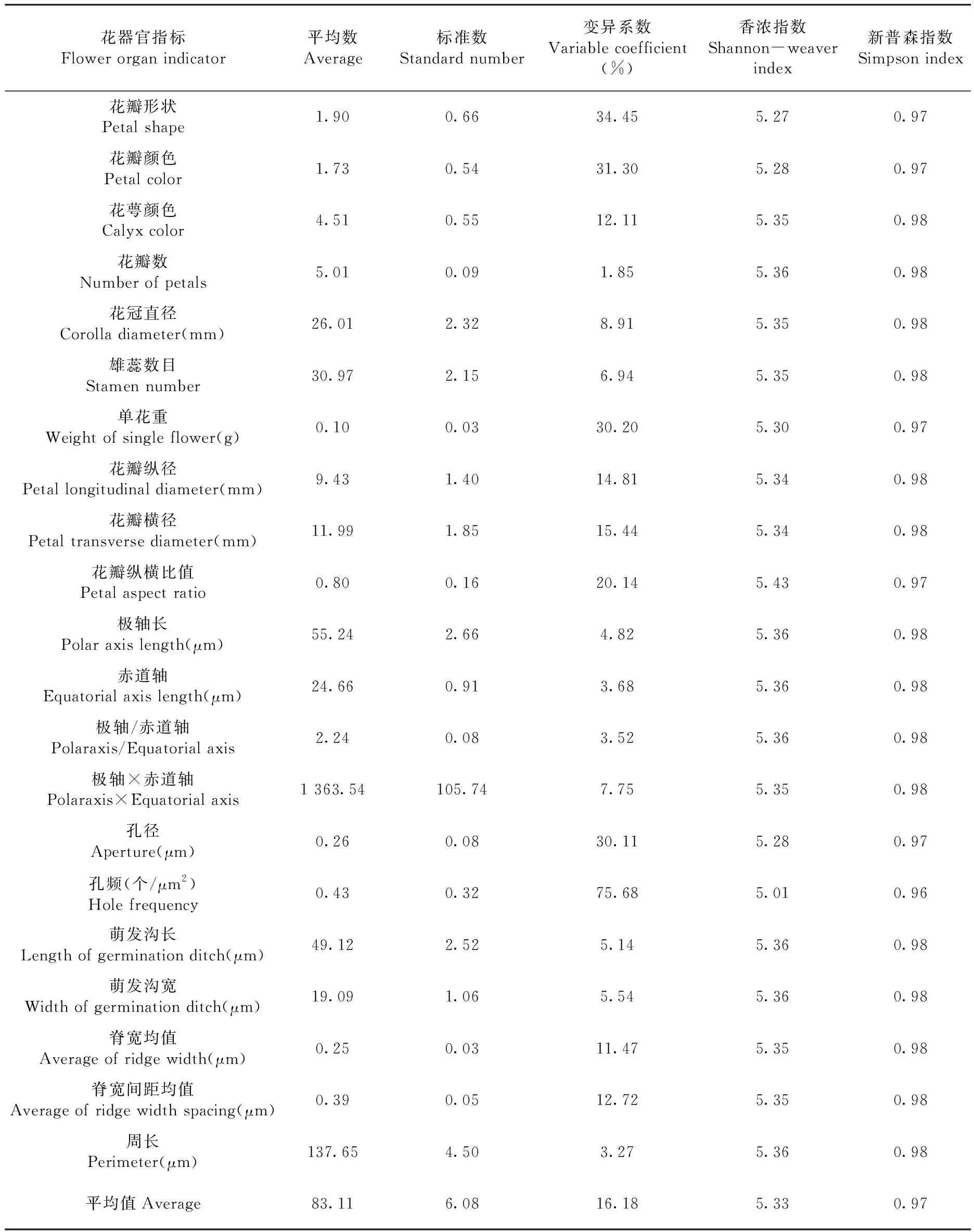
表5 41个新疆杏品种变异系数、Shannon-weaver指数和Simpson指数Table 5 Variable coefficient, Shannon-weaver index and Simpson index of different populations of 41 apricot varieties in Xinjiang
果树中花粉形态(花粉粒的形状、大小、表面纹饰等)与品种差异、品种自身遗传特性等因素息息相关,这就决定了它是判别物种之间亲缘关系的重要依据廖明康等[19]研究发现,不同种和品种花粉的大小、形状、外表纹饰排列及孔频等特征存在明显差异,花粉外壁纹饰种间虽有相似排列,但纹饰粗细、孔频多少各具特色;王玉柱等[20]研究发现花粉粒大小在杏品种间差异较大,不同杏品种间花粉的大小存在一定的差异。基于前人花粉微观特征对杏的亲缘关系研究,如:花粉形态相似性越大的单株亲缘关系越近,且这种分析亲缘关系的方法虽可应用于种或种质类型的水平上,却不适用于品种间[21,22],也有研究[23]持相反观点,认为品种间的亲缘关系分析也可以使用花粉表观形态聚类的方法。试验不同的新疆杏品种间花粉的微观特征花各指标存在一定差异,主要差异体现在极赤比值、孔频、孔径等方面。
花器官特征聚类分析中的分组与前人对新疆杏品种所属亚群的划分不完全吻合[14],如:黄其力干属于库车亚群,洛浦洪待克和脆佳娜丽都属于和田亚群,这3个品种聚为一组,和田亚群和库车亚群品种的花器官性状较为相似,亲缘关系较为亲近;而在另一组中,粗黑叶杏、阿克阿依、麻木达西杏、乔尔胖都属于喀什亚群,佳娜丽、古木杏属于和田亚群,毛拉肖和冬杏属于库车亚群,这8个品种聚类为1组说明喀什亚群、和田亚群、库车亚群3个亚群之间栽培杏品种存在较大相似性,这受到杏种质遗传变异、地域间基因交流和花器官测量过程中人为主观性因素影响,但供试的41个新疆杏品种总体亲缘关系较近。
在聚类过程中喀什亚群和和田亚群间划分界限不明显,如有部分和田亚群的品种被划分到了包含喀什亚群品种较多的1组中,古木杏和脆佳娜丽这2个品种;中熟佳娜丽、麻木达西杏、托乎提、细黑叶杏这4个喀什亚群的品种被划分到和包含和田亚群品种较多的1组,造成这种现象可能是由于引种和花粉传播逐渐在3大地理亚群繁殖,所以部分性状被同化,得出这一最终结论的原因应该由地理条件、环境气候、遗传背景差异或者人为引种等诸多因素造成。
通过花粉微观形态聚类将供试的41个品种划分为3类,大部分品种以地域为依据进行划分,而花器官表型聚类将供试的41个品种划分为4类,其中粗黑叶杏、阿克阿依、古木杏、冬杏、麻木达西杏、毛拉肖、乔尔胖、佳娜丽这8个品种在花器官聚类中单独划分为一大类,而在花粉微观聚类中按照其所属亚群被划分3个组中,由于考虑外界因素的影响,认为杏花器官表型性状相对花粉微观形态易受外界自然因素干扰,其数据在测量过程中也受人为主观因素影响较大,花粉微观形态特征比杏表型花器官性状数据更保守。
3.2 新疆栽培杏品种遗传多样性
植物表型性状既具有变异性,又有其稳定性,同时受植物自身遗传物质和生态环境两方面的影响[24]。刘娟[11]通过对叶片、果实、果核、种仁方面的形态特征对新疆栽培杏表型性状多样性进行了研究,得出造成表型差异的主要因素是单果质量、鲜核质量、鲜仁质量、果实形状、叶片长度果形指数、风味、硬度和可溶性固形物等指标,遗传多样性较丰富。研究中21个表型性状的遗传变异系数(16.18%)、Shannon-weaver指数(5.33)和Simpson指数(0.97)的平均值得出新疆栽培杏品种间表型遗传多样性丰富。变异系数幅度为1.85%~75.68%,孔频(75.68%)、花瓣形状(34.45%)、花瓣颜色(31.30%)、单花重(30.20%)、孔径(30.11%)这4个表性指标受自身的遗传基因和生长期的气候,如光照和水分等影响,导致变异系数较大。刘娟等[14]研究新疆杏表型多样性时也发现其表型性状受生长环境的影响较大。
4 结 论
41个新疆栽培杏品种总体亲缘关系较近,且遗传多样性丰富,主要体现在花瓣颜色、花瓣形状、单花重、孔径、孔频这5个方面。亲缘关系近的品种有贝新纳尔、英吉沙杏、赛买提、胡安娜、白色买提5个品种,在花器官聚类中克孜西米西与其余品种亲缘关系较远,在花粉聚类中,克孜西米西,叶城黑叶杏、粗黑叶杏、阿克达拉孜、晚熟杏这5个品种与其余品种的亲缘关系较远,克孜佳娜丽、克尔基胡安娜、卡拉胡安娜这3个品种在花器官表型和花粉微观2个聚类图中都紧密聚类为1组;也有在2个聚类图中同一品种分组不同的情况,如洛浦1号、洛浦洪待克这2个和田亚群的品种在花粉聚类中被紧密划分为在同一小组,但是在花器官聚类中不在同一小组。21个表型指标中孔频的变异最大,其变异最丰富,花瓣数变异最小,其遗传稳定性较好。花瓣纵横径比值、单花重、花瓣颜色、花瓣形状、孔径和孔频是进行表型多样性观测时的重要指标。
