感染高毒力球孢白僵菌的大蜡螟幼虫转录组分析
2021-02-01李敬龙
张 力,李敬龙
齐鲁工业大学(山东省科学院) 生物工程学院,生物基材料与绿色造纸国家重点实验室,济南 250353
大蜡螟(Galleriamellonella)是一昆虫纲,鳞翅目,螟蛾科昆虫,其免疫特点与哺乳动物近似,体内蛋白质含量极其丰富,利用其生长繁殖周期短等特点,常常被广泛应用在病原菌感染模型当中,以此来研究致病的机制和相关基因转录组的分析[1-2]。
球孢白僵菌(Beauveriabassiana)在国内外被用于害虫的生物防治实验与研究,且取得很好的对害虫持续性的消除控制功效,其被广大科学研究者称为最具开发潜力的昆虫病原真菌之一[3-4]。实验室从大蜡螟上分离得到6个样品并进行转录本的对比分析,共检测到12 770个共表达的基因。我们利用转录组测序技术分析了大蜡螟幼虫,在受白僵菌侵染的免疫应答相关基因,使用 TRIzol 法,利用苯酚等物质从感染了 B6 菌株和感染野生菌株的 3 d的大蜡螟幼虫(200 mg)中分离提取出总 RNA,并对其进行定性和定量分析,研究高毒力菌株侵染的大蜡螟幼虫的转录组变化情况,为从分子学水平上应用白僵菌防治生物害虫的研究奠定信息学基础。
1 材料与方法
1.1 菌株和供试昆虫
实验中使用的球孢白僵菌野生菌株B.bassianaARSEF2860,由实验室保存。实验中使用的球孢白僵菌高毒力菌株为实验室通过ARTP和FACS诱变筛选得到。
1.2 白僵菌侵染
将4龄的大蜡螟幼虫分别浸泡在浓度为107个/ml的野生型和高毒力菌种B6的分生孢子悬浮液中,以浸泡在0.02%无菌吐温-80中的大蜡螟幼虫作为对照组进行试验。浸泡10 s后,立即将大蜡螟幼虫拿出,并放入干净的透明塑料盒中,在室温下培养3 d。每组实验使用30头大蜡螟幼虫,处理组和对照组各取3个生物学重复。
1.3 RNA提取、文库制备和转录组测序
使用TRIzol法,利用苯酚等物质从感染了B6菌株和感染野生菌株的3 d的大蜡螟幼虫(200 mg)中分离提取出总RNA,并对其进行定性和定量分析。用磁珠对mRNA进行纯化,分别合成两条cDNA产物。cDNA片段产物纯化验证后,用phi29扩增以制成DNA纳米球并在BGISEQ-500平台上读取150 bp的单端读数。
利用Trimmomatic对原始数据进行过滤,去掉原始数据中带有接头的数据、含N的比例大于5%的数据以及低质量数据,最后获得纯净数据。将纯净数据与参考基因组进行对比。为探究大蜡螟幼虫对高毒力菌株侵染的反应变化,将差异倍数为两倍以上(|FC|≥4),Q≤0.001的基因定义为显著差异表达基因。对差异基因进行GO和KEGG的基因功能和生物通路进行分类分析和富集分析,Q≤0.05视为显著富集。
2 结果与分析
研究对球孢白僵菌高毒力菌株和野生型菌株侵染得到的6个样品(B6和野生型各三组样品)进行转录本的对比分析,在高毒力和野生型球孢白僵菌菌株感染72 h的大蜡螟4龄幼虫中,共检测到12 770个共表达的基因。用Log2 Fold Change(FC)作为筛选标准,得到了|FC|≥4,Q≤0.001的224个上调基因和227个下调基因,并定义为显著差异表达的差异基因。
2.1 显著差异基因的GO(Gene ontology)GO分类
Gene Ontology分类分析根据451个显著差异基因的功能将其分为分子功能(molecular function)、细胞组分(cellular component)和生物过程(biological process)等三大类中的33条GO二级条目中。
注释在生物过程分类中的差异基因分布于13条GO二级条目,其中,注释到代谢过程的差异基因数目是最多的,其次是细胞过程条目。注释到细胞过程的17个差异基因中,有15个是显著下调的,注释到代谢过程的23个差异基因中有14个是显著下调的。注释在细胞组分分类中的差异基因分布于12条GO二级条目,其中,注释到膜和膜的一部分条目的差异基因数目是最多的。注释到这两个条目的63个差异基因中,有42个是显著上调的。注释在分子功能分类中的差异基因分布于8条GO二级条目,其中,注释催化活性条目的差异基因数目是最多的,其次是结合。注释到这两个条目的差异基因分别是52个和45个,都分别有31个是显著下调的,具体分类见表1、表2和表3。

表1 感染高毒力菌株的大蜡螟幼虫转录组中差异表达基因的GO分类(生物过程) 个

表2 感染高毒力菌株的大蜡螟幼虫转录组中差异表达基因的GO分类(细胞组成) 个
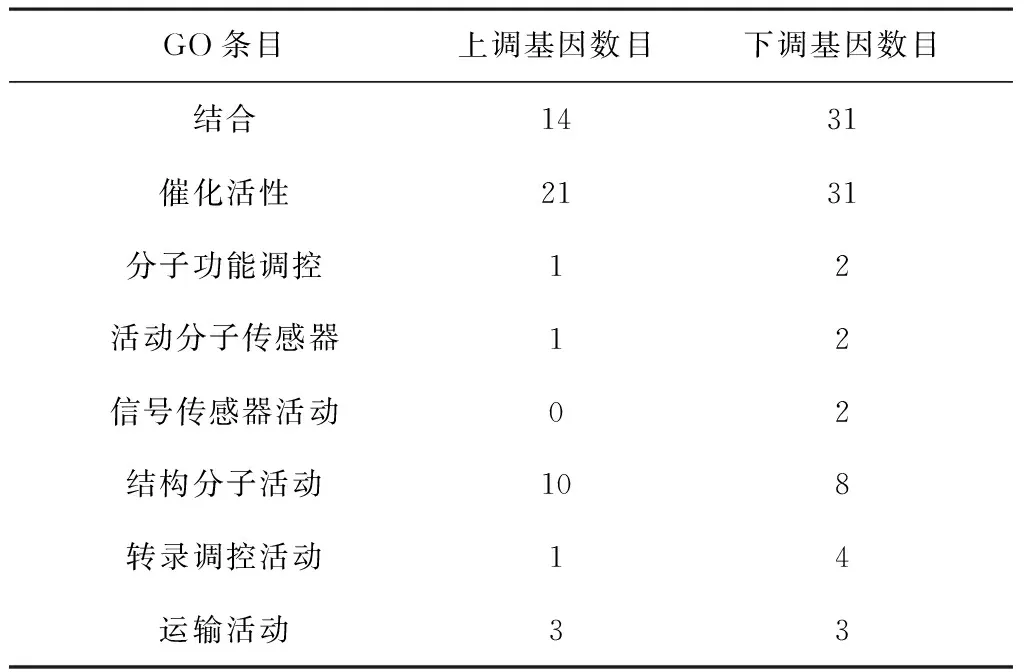
表3 感染高毒力菌株的大蜡螟幼虫转录组中差异表达基因的GO分类(分子功能) 个
2.2 显著差异基因的KEGG(Kyoto Encyclopedia of Genes and Genomes)富集
将显著性差异的基因进行KEGG富集,451个差异基因注释在了231条代谢通路上。其中有18条条目的Q≤0.05,被视为了显著富集。在显著富集的18条途径当中,注释在蛋白质消化和吸收途径,胰液分泌,肾素血管紧张素系统和溶酶体途径中的上调基因是多于下调基因的。其余的代谢途径中下调基因数目多于上调基因。其中,注释在脂肪酸合成途径的13个差异基因中有9个是显著下调的。注释在脂肪酸代谢途径的22个差异基因中有17个是显著下调的。注释到RNA聚合酶途径的16个基因中有11个是显著下调的。注释在AMPK途径的15个基因中有11个是显著下调的见表4。
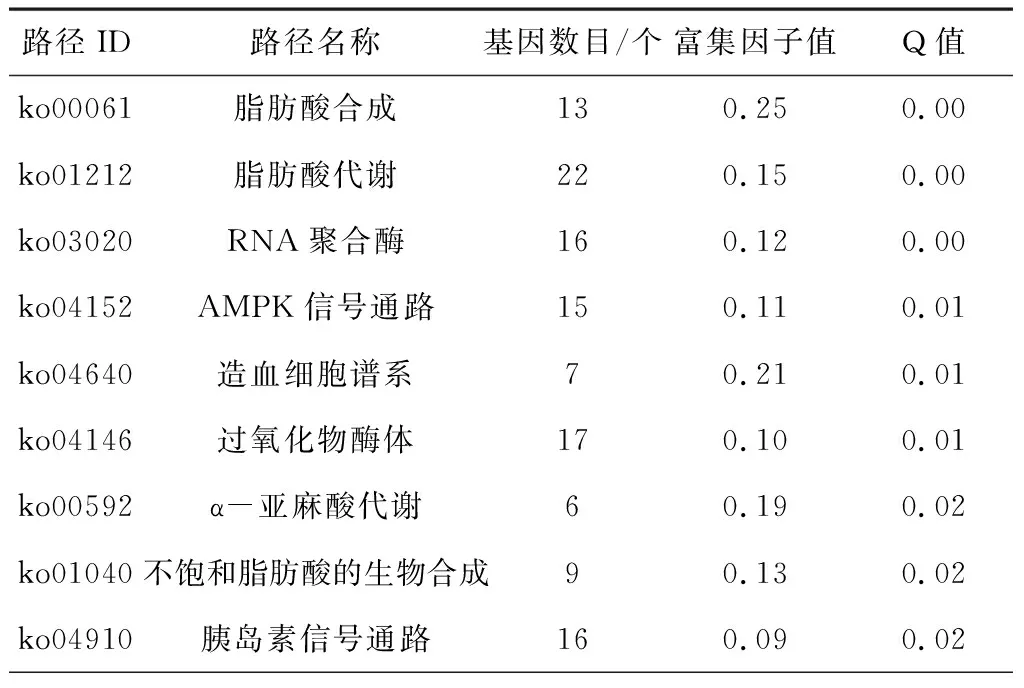
表4 感染高毒力菌株的大蜡螟幼虫转录组中差异表达基因的KEGG富集
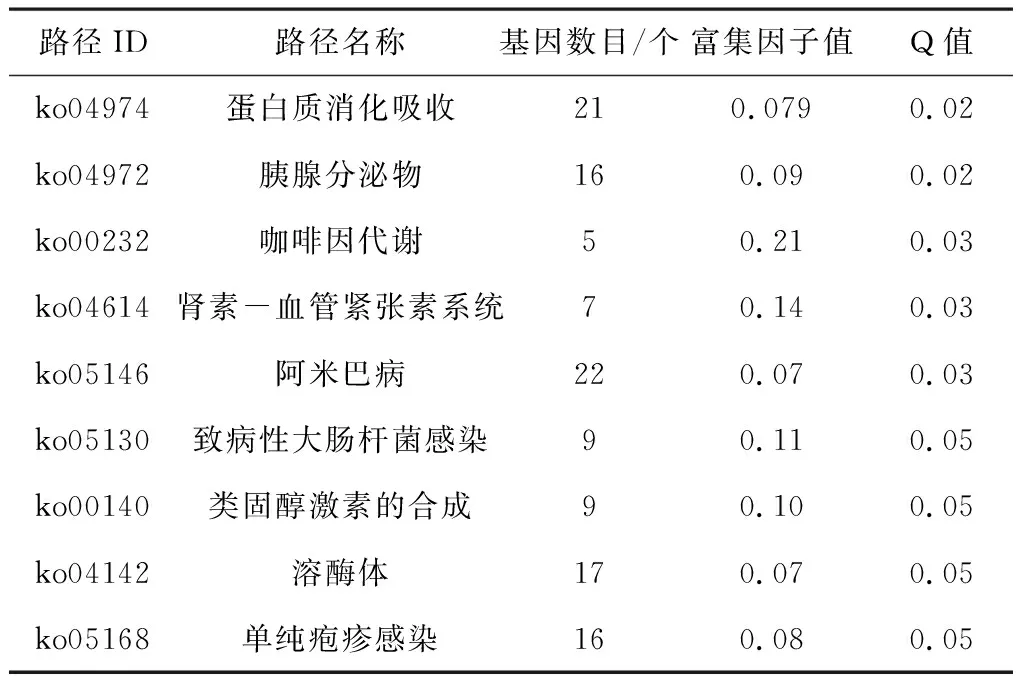
续表
2.3 显著差异基因分析
为了进一步研究高毒力菌株侵染的大蜡螟幼虫的转录组变化情况,我们把所有差异基因按照差异倍数,即Log2 Fold Change进行排序,分别统计了上调和下调程度最高的20个差异基因进行分析。
结果显示,前20个上调基因的FC范围在11.29~8.02。其中,2个与麦芽糖酶表达相关的基因,2个与胰蛋白酶表达相关的基因和3个与氨肽酶表达相关的基因是显著上调的。这与KEGG富集分析蛋白质消化吸收途径的上调是一致的。前20个下调基因的FC范围在-12.55~-7.47。值得注意的是,其中有4个与脂肪酸合成相关的酶和一个与脂肪酶相关的酶是显著下调的。这与富集分析中脂肪酸合成和代谢途径的下调结果是一致的,见表5和表6。
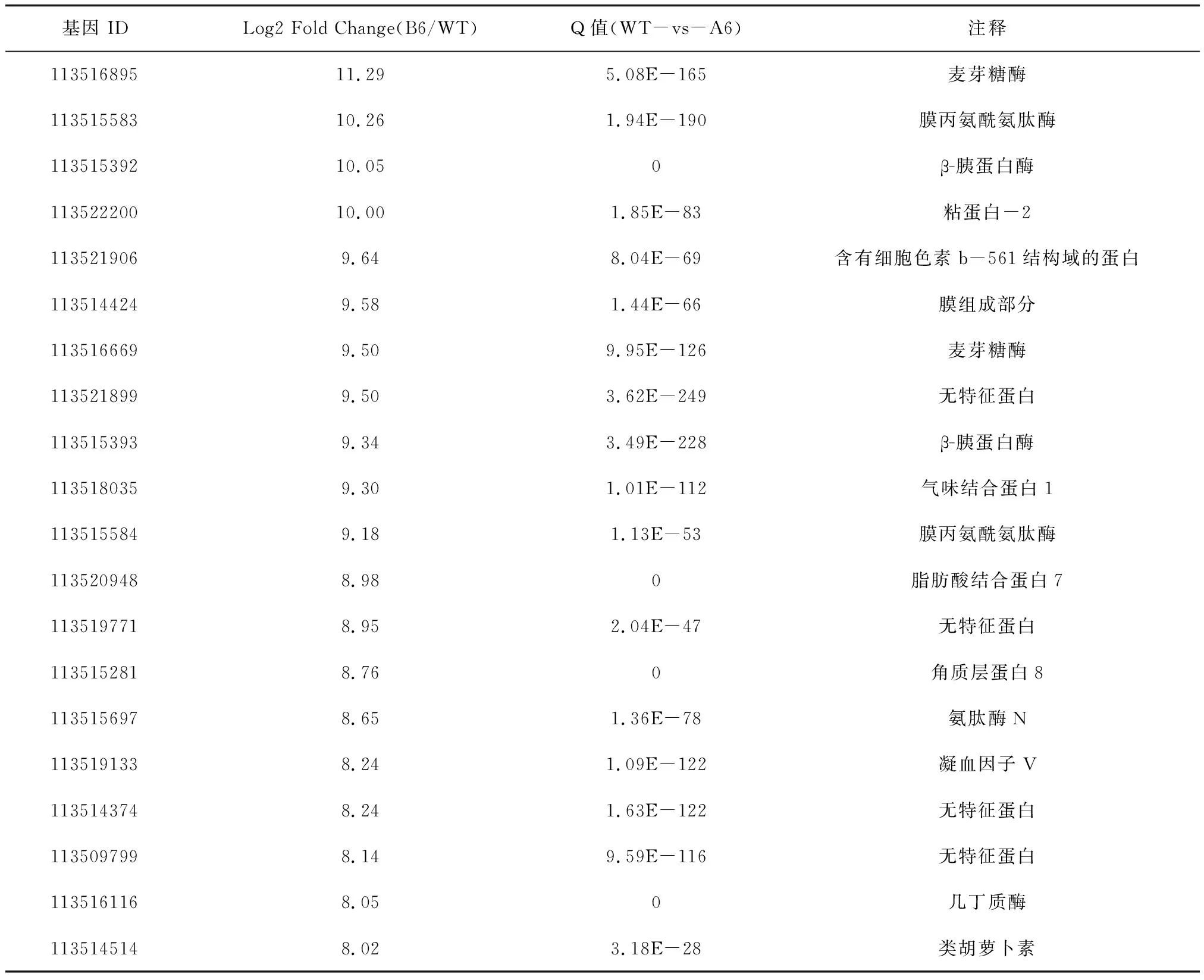
表5 B6对WT感染的大蜡螟DEGs的前20个上调基因
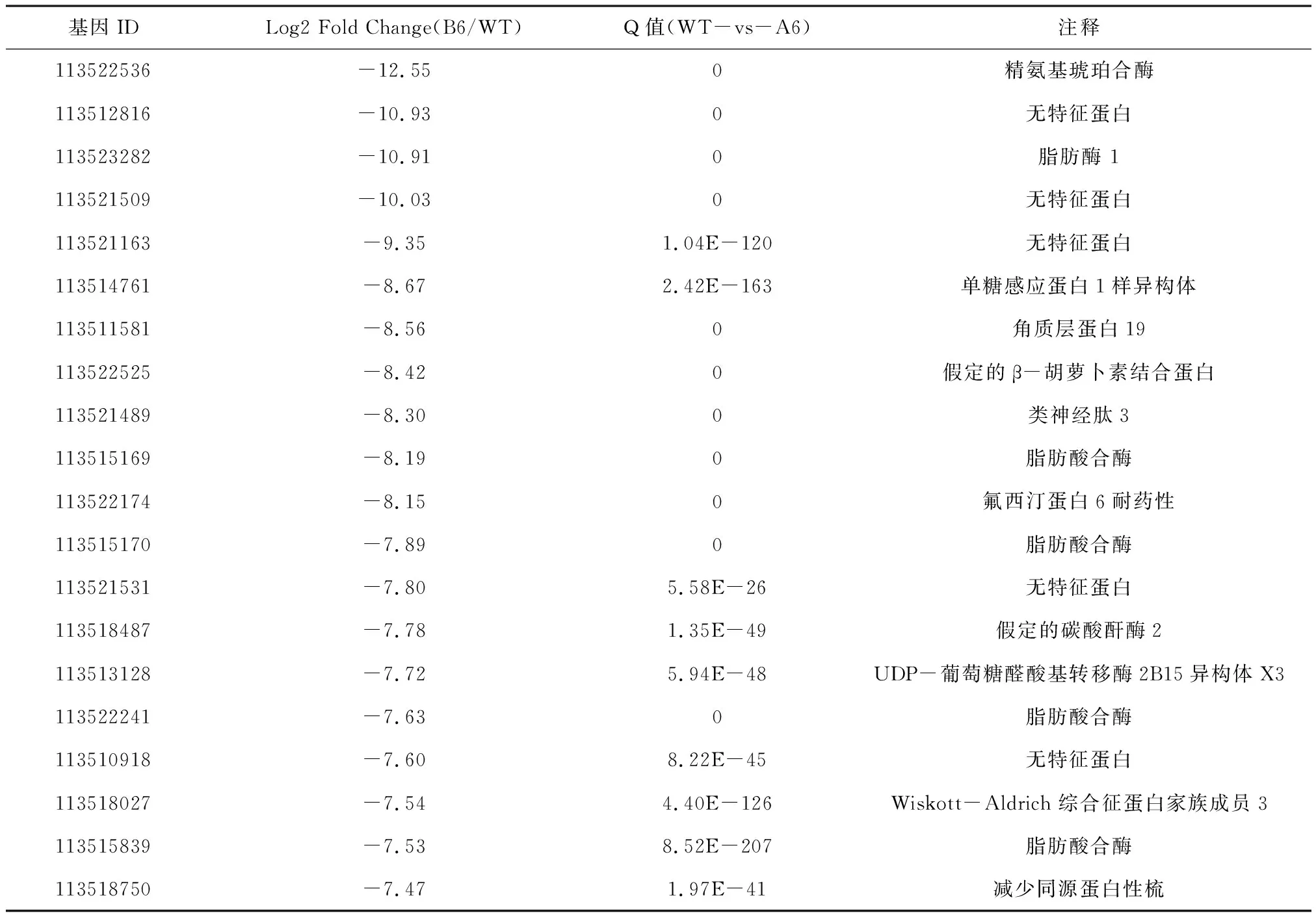
表6 B6对WT感染的大蜡螟DEGs的前20个下调基因列表
3 讨 论
当球孢白僵菌侵染寄主时,首先会将分生孢子附着在昆虫角质层上并萌发菌丝体,在此过程中分泌蛋白酶和几丁质酶等会破坏角质层[5]。菌丝体穿透角质层到达宿主血腔后,会破坏血细胞的细胞壁结构,分泌次生代谢物以逃避宿主免疫[6]。分生孢子转换成芽生孢子以便快速增殖并消耗宿主营养[7-8]。此外,球孢白僵菌还会产生白僵素、卵孢霉素有毒代谢物加速昆虫的死亡[9-10]。
在昆虫病原性真菌的研究中,对宿主进行转录组分析可以对宿主的病原体的相互作用进行整体性研究[11-12]。本研究对球孢白僵菌高毒力菌株B6和wt感染3 d的大蜡螟4龄幼虫转录组进行了G0分类注释和KEGG富集分析。分析表明,在B6感染3 d后的大蜡螟幼虫中检测到622个上调基因和869个下调基因。用Log2 Fold Change(FC)作为筛选标准,得到了|FC|≥4,Q≤0.001的224个上调基因和 227个下调基因。
转录组分析结果表明,高毒力菌株侵染3 d后,大蜡螟体内脂肪酸合成,AMPK信号通路,糖原异生等代谢过程以及催化活性,RNA聚合等活性受到了高度抑制。其中,脂肪酸合成途径中,14个编码脂肪酸合成酶(FAS)的酰基载体蛋白的基因被下调,从而抑制了脂肪酸的合成。值得注意的是,下调的14个编码FAS酰基载体蛋白的基因与AMPK信号通路高度相关。AMPK信号通路是生物体中保持和调节细胞能量动态平衡的重要信号通路[13-14]。
除此之外,大蜡螟幼虫中的蛋白质的消化和吸收进程,肾素血管紧张素系统以及免疫反应被高度激活。在蛋白质的消化吸收途径中,21个分别编码胰蛋白酶、羧肽酶和氨肽酶N的基因被显著上调。在大蜡螟免疫反应相关的基因中,与吞噬体,溶酶体和生物刺激反应相关的12个基因被上调。除此之外,9个与抗菌肽合成有关的基因也被显著上调。使用印楝素处理的Bactroceradorsalis幼虫在组织蛋白酶高度表达时,幼虫的生长和发育受到了显著抑制[15]。家蚕和血吸虫通过上调C-溶菌酶的表达来抵抗病毒和细菌的侵袭[16-17]。
研究对侵染高毒力菌株的大蜡螟幼虫转录组进行分析,从转录组层面对宿主和高毒力病原体的相互作用进行了进一步研究。为筛选高毒力病原体提供了潜在靶点。
