沟渠N2O排放研究进展
2021-01-29张丰粟王小国
张丰粟,王小国①
(1.中国科学院山地灾害与环境研究所,四川 成都 610041;2.中国科学院大学,北京 100049;3.中国科学院山地表生过程与生态调控重点实验室,四川 成都 610041)
全球气候变暖是当今人类面临的严峻环境挑战,大气中温室气体浓度的不断增加是导致气候变暖的主要成因[1]。N2O是当前最受关注的温室气体之一 , 其百年尺度的全球增温潜势(global warming potential,GWP)约为 CO2的296倍,自1750年以来对全球总温室效应的贡献率达6%[2]。N2O产生途径主要有硝化作用和反硝化作用两个过程,它们所产生的N2O约占全球大气中N2O总量的90%[3]。N2O在对流层中的存留时间可长达120 a,在平流层中经催化作用分解成一氧化氮后与臭氧分子反应,是破坏臭氧层、导致臭氧层空洞的重要因素[4]。鉴于大气中N2O具有较高GWP、较长存留时间以及较强的臭氧层破坏效应,关于N2O排放的研究始终受到广泛关注。
沟渠是介于陆地生态系统和水生生态系统之间的过渡净化带,常处于淹水状态和非淹水状态的交替过程,导致其硝化过程和反硝化过程活跃[5]。沟渠作为一种独特的生态系统,有着很高的氧化还原能力,这使得它在具有高水质净化作用的同时,兼有较高的温室气体排放通量;同时,作为连接农业排水、村镇居民生活废水与河流湖泊的重要通道,沟渠生态系统受人为干预影响较大,常常含有高浓度碳、氮物质,是生物地球化学作用非常活跃的场所[6-9]。根据进入沟渠中水体类型的不同,主要可以将其分为农田沟渠与生活污水沟渠。农田沟渠水源主要来自农田的淋溶水以及地表径流,主要特点为泥沙含量大,并且携带大量可溶性碳、氮等营养物质[10]。此类沟渠一般具有较大的表面-体积比,其内部沉积物及植被的存在有利于微生物反硝化脱氮的发生[9]。生活污水沟渠作为村镇生活废水与河流连接的通道,水体主要来源为生活废水,具有较高的碳、氮负荷,并且含有很多高分子有机污染物,N2O排放通量也相对较高。目前,国内外学者对湿地中N2O的产生、排放通量及影响因素等做了大量研究,但对于各类沟渠N2O的排放研究较少,该文综述了沟渠N2O产生、排放及其影响因素,以期为生态沟渠的环境友好管理提供理论依据。
1 沟渠N2O排放研究
1.1 沟渠N2O排放通量的测定方法
目前,国际上已有多种方法用于测定水生生态系统N2O排放通量,主要分为野外观测测定和模型估算法。密闭箱法[11-12]和开放式动态箱法[13]是目前野外原位水-气界面通常采用的两种观测方法。密闭箱法操作过程较便捷,技术要求不高,是浅水域N2O排放通量研究最常用的方法。该方法具体操作为在水体表面放置顶部密封、底部中通的箱体,按一定时间间隔定量收集箱内气体,然后利用气体分析仪监测待测气体浓度,最后根据气体浓度随时间变化速率计算覆盖水体N2O排放通量;但该方法可能会受到人为与水面波动的干扰,产生误差,且无法实现大范围、全时段连续观测。开放式动态箱法通过水泵与尼龙管相连,使得空气进入动态箱体,然后用痕量气体分析仪对采集的样品浓度进行分析,其计算公式为F=f(Co-Ci)/A。其中,F为N2O气体排放通量,f为气体流速,Co和Ci分别为尼龙管进口和出口气体浓度,A为水面交换面积。此方法测量精度较高,连续性好,但技术要求较高。此外,可调谐二极管激光吸收光谱[14](tunable diode laser absorption spectroscopy,TDLAS)技术和微气象学观测方法中的涡度相关法[15-16]是用于对水域温室气体扩散通量进行大面积连续测量最有效、精确度较高的两种方法,但其对环境条件和技术要求均较高。目前国际上己采用上述两种方法对面积较大的水库和湖泊进行观测研究,但国内还较鲜见。
模型估算法又称梯度法[17-18],首先测量表层水和大气中N2O浓度,并计算两者差值,运用Fick定律估算N2O排放通量F,计算公式为F=kx(Cwater-Cair)。其中,kx为气体交换系数,Cwater为水体表层溶解的N2O浓度,Cair为大气中对应N2O浓度。此方法气体交换系数kx值的确定是解决该方法模拟精度的关键,但其值时常受到表层水体絮流混合作用及环境因素影响而变得难以确定,因此模型估算结果存在较大的不确定性[19]。
1.2 国内外各类沟渠N2O排放通量研究
沟渠类型多样,广泛分布于世界各地,水和污染物通过地表径流、壤中流等进入沟渠[20]。沟渠主要承担对污水的传输与净化功能,在各种净化方式(如底泥吸附、植被吸收等)中,尤以微生物反硝化脱氮对氮素的去除最彻底[21]。在高氮、低碳和低氧条件下反硝化细菌脱氮作用显著,生成的N2O通过水-气界面排放,是水体排放N2O最主要的方式[22]。BEDARD-HAUGHN等[23]研究表明小型湿地超过75%的N2O排放是通过反硝化脱氮作用产生的,而反硝化速率受温度、水体溶解氧和pH等很多因素影响,不同研究区反硝化速率差异较大。植物传输主要指沉积层产生的N2O通过分子扩散或对流方式进入水生植物输导组织,并向上输送进入大气,是植被丰富的水域区N2O排放的重要方式。
表1[24-38]显示,研究以农业沟渠居多,对生活污水沟渠与天然沟渠的研究较少,且不同纬度地区N2O排放呈明显空间差异。已报道的沟渠N2O排放通量范围为-9.2~56 580 μg·m-2·h-1,表明N2O排放通量变化在空间、时间上的异质性较大。已报道的最大值在日本一条农业沟渠测得,其测定时间为农田施肥时期,沟渠中汇集的淋溶水具有较高的碳、氮负荷,为硝化-反硝化过程提供了丰富的底物;最小值在美国Neuse River河流沟渠测得,该河流水质优良,且在测定时温度较低,反硝化微生物活性较低。DE CLEIN[39]研究表明,温度是影响反硝化速率最关键的因子,当温度达到30~35 ℃时,反硝化细菌活性最高,反硝化速率最强,并且较高的温度可以促进矿化和硝化作用,为反硝化过程提供更多的硝酸盐。由表1可知,N2O排放通量高峰大都出现在温度最高的夏季。冯香荣等[30]对亚热带生活沟渠进行周期为1 a的N2O排放通量观测,排放极大值出现在6月,通量为70.35 μg·m-2·h-1,极小值为12月测得的-2.27 μg·m-2·h-1,结果表明N2O排放存在显著的时间变异性;这主要是由于季节气候的变化通过影响土壤理化性质、植被生长和微生物活性进而影响沟渠中N2O排放。田琳琳等[34]在川中丘陵区农田沟渠的不同位置设置多个采样点位,发现沟渠中污染物浓度与植株的有无对N2O排放都会产生巨大影响,这表明同一条沟渠可能因为不同区段环境因子差异而出现N2O排放差异。一般来说,生活污水沟渠年N2O排放通量高于农田沟渠,主要是因为生活污水沟渠常年处于淹水状态,并且水体具有较高的碳、氮负荷,厌氧反硝化作用显著。但是农田沟渠N2O排放通量的季节性变化更加明显,这主要是由于受农田施肥及季节性降雨的影响,沟渠内水体养分含量随之变化。另外,在纬度相差不大的地区,董宏伟[26]和李飞跃等[35]关于农业沟渠N2O排放通量的研究结果相差数十倍,这是由于两地沟渠形态和底泥基质养分含量不同所致。此外,沟渠内植被也可以显著影响N2O排放。植物不仅可以自身排放N2O至大气中,还可以明显降低水中氧气溶解度从而提高反硝化速率,促进N2O排放[40-41]。
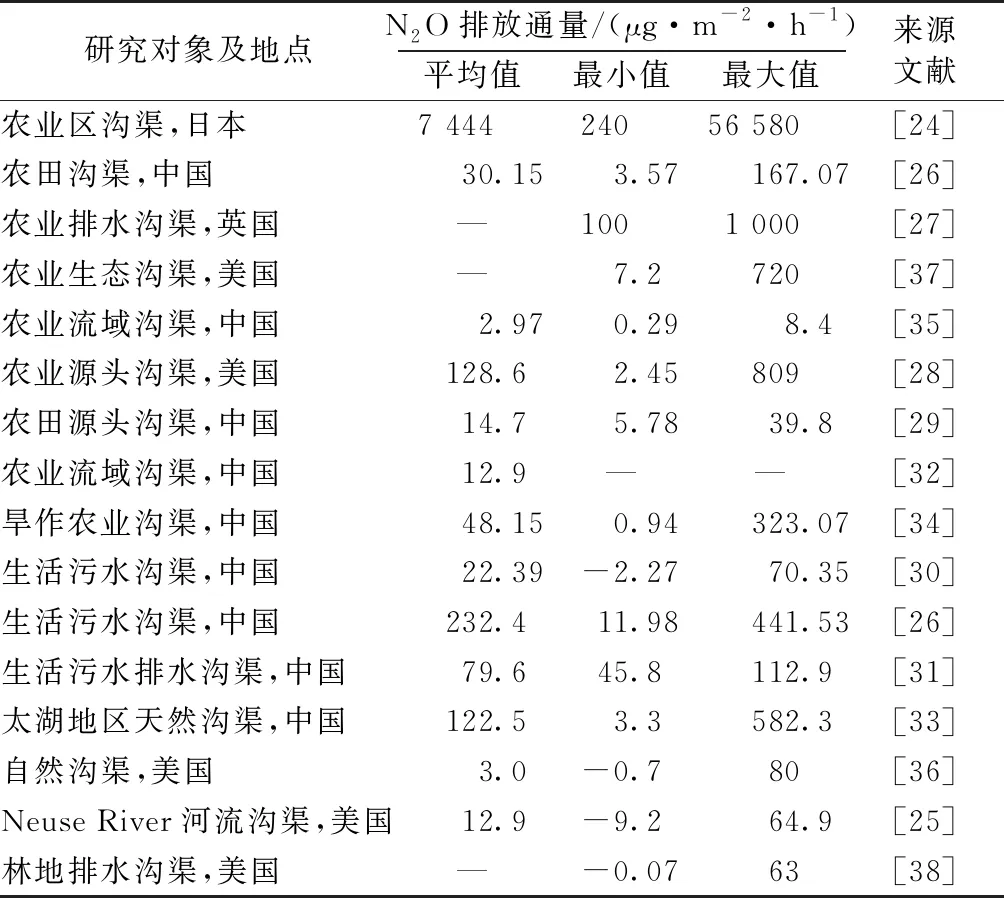
表1 不同地区沟渠 N2O 排放通量比较
2 沟渠N2O排放的主要影响因素
2.1 水体可溶性有机碳(DOC)含量
可溶性有机碳(dissolved organic carbon,DOC)是陆地和水生生态系统中重要和非常活跃的化学组分,也是很多环境污染物迁移的载体[42]。DOC具有一定的溶解性,有易流动、易分解、易氧化和生物活性高等特性[43],是异养反硝化微生物重要的能量与物质来源,也是进行反硝化作用的物质基础(电子供体),其含量直接影响微生物的新陈代谢和活性,进而间接影响反硝化速率和 N2O 的产生[44]。农田淋溶水与生活污水均含有大量杂质及DOC等营养物质,此类水体进入沟渠后会促进N2O排放。如HARRISON等[45]对亚热带地区农业沟渠的研究发现,N2O排放与沟渠中水体DOC含量密切相关,水中有机碳含量可用于较好地预测N2O排放(r2=0.52)。JURADO等[46]将高DOC含量的水源注入水稻田后,发现DOC的加入大大提高了水稻田N2O排放速率,这可能是由于DOC的加入可促进反硝化作用,进而促进N2O排放。JAHANGIR等[47]研究也发现,农业区地下水中 N2O浓度与DOC含量呈正相关,这表明反硝化作用更容易在富含有机质的地下水厌氧微环境中发生,水中DOC含量的增加可促进N2O的产生。
2.2 水体溶解氧(DO)含量
水体中DO主要影响硝化、反硝化和厌氧氨氧化等 N2O 产生过程,也影响电子受体 O2和 NO3--N间的竞争关系及酶活性等,进而最终影响N2O的产生及排放量[48-49]。研究表明,N2O还原酶(Nos)对DO含量最敏感,少量 DO(如 22.3 μmol·L-1)即可抑制 N2O的还原,因此,当DO含量较低时反硝化可产生较高的N2O[50]。SILVENNOINEN等[51]发现,在Bothnian Bay的河流水-沉积物界面中随O2含量增加其反硝化作用受到限制,从而使N2O排放量最高值出现在ρ(O2)低于0.2 mg·L-1的河流环境中。TOYODA等[52]研究认为,在高NH4+-N和低DO含量水体中,反硝化作用有助于N2O的产生。LIM等[53]的研究表明,由于微生物的反硝化作用是一个严格的厌氧过程,当水体中ρ(DO)超过0.2 mg·L-1时,反硝化作用难以发生。对加拿大安大略省内多条沟渠的研究发现,沟渠中高N2O产生量与较低DO含量紧密相关,在低DO含量区,反硝化作用是N2O产生的主要来源[54]。YU等[31]发现,上海城市河网排水沟渠中DO含量是影响其N2O排放的主要因子,表明沟渠中反硝化脱氮程度依赖于DO含量变化。ROSAMOND等[55]研究发现与NO3--N含量相比,水体DO含量对N2O排放影响更大,DO含量与N2O排放通量呈显著负相关,并提出在未来研究中应将水体DO含量纳入N2O排放预测模型中。
2.3 底泥有机碳、氮含量
由于沟渠水体往往携带大量泥沙,进入沟渠后泥沙会沉淀,并且作为沟渠基质为N2O产生提供条件。由于绝大多数反硝化细菌是化能异养型,土壤中有机质可以为此类细菌提供电子受体和养分,是调控土壤生物反硝化速率和作用强度的重要因素[56]。GROFFMAN等[38]研究发现底泥有机质含量高的沟渠反硝化速率较高。MEGONIGAL等[44]研究发现,由于土壤有机碳可以直接加速有氧异养呼吸,产生厌氧环境,所以在有机碳含量较高的土壤中形成厌氧小区的现象十分明显,这更有利于厌氧反硝化作用的发生。土壤中有机碳对反硝化过程的主要影响可能是推动无氧环境的形成。由于硝化-反硝化进程主要取决于土壤氧气含量的高低,而氧气的消耗速率主要受有机碳可利用性的控制,土壤中较高的有机碳含量可以直接加速有氧异养呼吸或者间接导致厌氧还原剂的产生,继而与氧气发生反应,产生土壤厌氧环境[57]。底泥中有机氮是N2O产生的源,底泥土壤碳氮比(C/N)则是影响N2O产生的重要因素。一般认为,微生物的最适C/N比值为25~30,若C/N比值过高,微生物活性较弱,有机质分解缓慢,N2O释放则受到抑制,反之则可以在一定程度上促进N2O释放[58]。黄耀等[59]研究发现C/N比值在7~12范围时,N2O排放量随着C/N比值的增加而减少。
2.4 温度
土壤温度通过影响微生物酶活性、生物学过程中酶活性和化学反应速率影响硝化-反硝化速率,进而影响N2O排放通量。ZIMMERMAN等[60]发现,温度是影响沉积物中反硝化速率的最主要因素,温度变化可解释52%的反硝化速率变异。RYDEN[61]的研究表明,在相同沟渠基质含水量和硝酸根含量条件下,沟渠基质温度从5 ℃增加到10 ℃,土壤反硝化速率会从0.02 kg·hm-2·d-1增加到0.11 kg·hm-2·d-1。宋长春等[62]对三江平原沼泽湿地的研究发现,冬季三江平原湿地是N2O汇, 随着土壤温度的升高而逐渐成为N2O源。黄树辉等[63]研究表明,在土壤湿度适宜的条件下,一定温度范围内,N2O排放通量(F)对土壤温度(t)的依赖性可用指数函数F=Aeat(A和a均为系数,e为自然对数的底数)来描述。徐亚同[64]对温度与反硝化关系的实验表明,温度为10~30 ℃时反硝化速率平行上升,当超过30 ℃时反硝化速率又开始下降。
水温是沟渠中水体的重要理化参数,也是可影响气体分子N2O、O2等的扩散速度及在水体中的溶解度进而影响N2O交换和传输速率的物理化学参数[65]。有研究表明,在一定范围内水体温度与N2O排放量呈正相关。MCMAHON等[66]对英国自然沟渠的研究表明N2O排放速率季节性变化明显,在温度较高的月份升高明显。JENNI等[67]对10~30 ℃温度范围条件下污水沟渠的研究表明,N2O的产生和排放随温度的升高呈指数增长。此外,国内也发现类似规律,比如:太湖地区河流湖泊、沟渠、地下水中N2O排放及其容存浓度与温度呈正相关[68]。对川中丘陵地区生活污水沟渠与农田沟渠的研究显示,N2O排放季节性差异明显,排放通量以夏季为最高,冬季最低[26]。
2.5 水体pH
水体pH对有机质分解与微生物活动的影响会对N2O净排放产生间接影响,但是,不同学者的研究结论存在较大差异。由于反硝化微生物原生质pH接近中性,所以大部分反硝化微生物适宜在中性条件下生长。黄耀等[59]研究表明,pH值在5.6~6.8范围时,N2O排放与pH呈正相关。PENG等[69]发现,通过调节pH来控制NH4+-N浓度可实现NO2--N的累积,而当pH 高于7.0时可限制NO2-转化为HNO2并确保游离氨选择性地抑制亚硝酸盐氧化细菌,从而有利于短程硝化的稳定。徐亚同[64]认为反硝化作用最适宜pH在7.5左右,当偏离这一适宜值时,反硝化速率会逐渐下降,当pH小于6.5或大于9.0时,反硝化速率下降明显。HARRISON等[45]对亚热带地区农业区沟渠水体中 N2O 排放的研究发现,农田周边排水沟渠中N2O排放与pH呈负相关,而养猪场附近排水沟渠中N2O排放与pH呈正相关。目前,关于pH与N2O排放量之间关系的研究结论不尽相同,这可能是由于碳源不同或其他因素(如温度、含水量)差异对土壤微生物综合作用所致,因此,此问题有待进一步研究。
2.6 干湿交替及水-气界面动力
由于季节性降雨、灌溉和居民用水的时空性差异,沟渠往往会经历干湿交替,从而对N2O排放通量产生影响。研究认为,反硝化作用会随着水位的增高而变强,当土壤孔隙水(waterfilled pore space,WFPS)>60%时,反硝化作用占据主导地位,而当WFPS>80%时,N2O扩散受到抑制,并且N2O可能作为电子受体被还原为N2[70]。当WFPS为45%~75%时,硝化细菌和反硝化细菌都可能成为N2O的主要制造者[71]。MENTZER等[72]研究发现含水量增加会降低水解酶活性,并指出干湿交替会改变氮循环过程,沟渠排干会刺激硝化作用,而积水增加则会刺激反硝化过程,进而影响N2O排放。DETTMANN[73]研究发现,河口地区沟渠中水体参与反硝化的氮素(N)比例随着淡水滞留时间的增加而升高,从而增加N2O排放通量。邓焕广等[74]对不同频率干湿交替土壤的模拟实验结果显示,较长时间间隔(7 d以上)的干湿交替或持续干旱后土壤反硝化速率会显著降低。VON ARNOLD等[75]研究表明沟渠排水后由于矿化作用导致沟渠土壤N2O排放量增加,没有排水的沟渠N2O年排放量比排水土壤低。在淹水期间,水-气界面N2O排放与沟渠中水动力息息相关[33],SILVENNOINEN等[76]对波罗的海 Temmesjoki河的研究表明较快的水流速度与较大的风速有利于水-气界面N2O交换。水流速度通过影响藻类生长环境及水体富营养化而影响光照在水体中的通透性,进而影响水生生物在水体碳氮循环中的活性[77]。风速则会通过影响水-气界面N2O分压平衡而影响水体N2O释放。一方面,风对于水体的切应力会导致水体表面破碎,增加水-气界面面积,促进水体N2O释放;另一方面,风速变化会影响N2O在水体中的饱和度,并引起N2O在源汇角色中转变[78-79]。
2.7 外源氮输入
在农田施肥之后,持续高氮负荷的淋溶水排入沟渠,外源氮输入会作为重要因素影响生态沟渠N2O 排放。TOWNSEND-SMALL等[37]对加利福尼亚一处农业沟渠的研究发现,施肥后沟渠N2O排放通量由施肥前7.2 μg·m-2·h-1上升至720 μg·m-2·h-1,增量幅度达近百倍;这是由于当高氮负荷的农田淋溶水进入沟渠后,NO2--N和NO3--N为反硝化作用的电子受体,NH4+-N和 NO3--N又分别为硝化和反硝化作用的底物,高氮负荷的沟渠水体增强了硝化和反硝化作用,使得N2O排放通量显著提高。PETERSON等[10]利用15N进行的原位实验研究发现,河溪中NO3--N浓度的增加可促进其反硝化作用。田琳琳等[80]研究发现,沟渠中NO3--N浓度的升高可促进水中反硝化作用,进而增加其 N2O排放。
外源氮输入将改变沟渠基质的碳氮循环和碳氮比,从而对N2O排放造成影响。在由于沟渠淹水导致的厌氧环境中,有机氮输入能够增加土壤耦合硝化的反硝化速率,促进N2O排放[81]。LINDAU等[82]发现,与输入NH4+相比,输入NO3-会刺激更多的 N2O排放,并认为这是由于严格的硝化作用限制所致,只有很少一部分增加的 NH4+被脱氮。此外,氮输入还可以通过影响植被而间接影响N2O排放。一般来说,氮输入会刺激植物生长,特别是根部生物量的累积,这有助于微生物通过植物根部获得更多碳源,为反硝化作用提供能量,从而促进N2O排放,并且过剩的氮输入还会使植物直接排放的N2O通量增加[83]。BEAULIEU等[84]研究表明,氮输入后N2O排放峰值只会持续几天,然后会逐渐下降;这可能是由于高氮负荷初期硝酸盐被迅速吸收转化,而后植被吸收了土壤中的有效氮,减少了硝化反硝化作用的反应底物,而剩余的氮又不足以改变土壤硝化-反硝化速率。在外源氮利用上,SILVAN等[85]在加入高浓度氮后的研究表明,15%氮被微生物利用,15%氮以气体形式直接排出,70%氮被植物吸收。然而,也有科学家的研究结论与上述结论相反。KETTUNEN等[86]研究发现氮输入增加了湿地生物量和枯落物,微生物在分解枯落物的初期需利用大量氮元素,这就会产生对氮的争夺,使土壤中可利用氮减少,导致N2O排放量降低。
2.8 沟渠植被
水生植被广泛分布于各类沟渠,是水生态系统的重要组成部分,并发挥着多种生态功能,同时也是C、N循环过程的调节者[87]。沟渠中植被对N2O排放的影响主要有以下几个方面:(1)植物会吸收沟渠基质中的营养物质,并且影响底泥的氧化还原及理化性质[88];(2)植物可以通过通气组织传递N2O[89];(3)植物根部分泌物能够给微生物提供碳源,并且提高反硝化细菌活性[90];(4)植物本身的生长代谢也能产生N2O[91]。董宏伟[26]对川中丘陵区生态沟渠有植株组与无植株组N2O排放通量的对比研究发现有植株组N2O排放通量远大于无植株组,这说明生态沟渠中植株在净化水体的同时也会产生大量N2O。此外,植被类型也可以影响生态系统中N2O的排放差异,如对黄河河口地区沟渠的研究发现,植被种类、覆盖度和生物量共同影响该沟渠生态系统N2O 排放的空间差异[92]。在不同植被类型的河岸带也发现反硝化速率会因植被类型不同而出现明显差异[93]。VERAART等[40]利用15N示踪技术研究了大型挺水植物、浮游植物和沉水植物对沟渠反硝化作用的影响,发现大型挺水植物对反硝化速率的影响最明显,浮游植物由于能明显降低水体氧气浓度而具有最高的反硝化速率。在一定条件下,植物与反硝化细菌对氮源的竞争会影响沟渠中反硝化作用,从而对N2O排放产生影响[94]。如SILVAN等[85]研究发现在白羊毛胡子草湿地中,植被丰度越大,反硝化活性越低,这可能是由于植物与反硝化微生物对土壤养分的竞争所致。PETERSEN等[95]研究表明水生大型植物区N2O排放量比无植株区高6倍。HASEGAWA等[24]对淹水稻田的研究发现有植株区N2O排放比无植株区约高37%。
3 结论与展望
目前,沟渠N2O排放研究主要集中在农田沟渠,对于生活污水沟渠的研究较少。在已有的研究中沟渠N2O排放表现出显著的空间、时间变异性,并且其排放受多个因素的影响,在污水沟渠的生态化建设过程与研究中,提出以下几点展望:
(1)应加强对沟渠N2O排放机制的研究,明确沟渠硝化与反硝化作用的容量,以及硝化和反硝化作用在N2O排放中所占的比例,为沟渠模型的构建提供更好的科学依据。
(2)国内外学者对于沟渠生态系统N2O排放的研究主要集中在建立各因子对N2O排放的影响,有关综合各因子之间的关系及各因子如何共同对N2O排放产生影响方面的研究尚缺乏。
(3)加强不同类型植被-土壤系统对N2O排放的影响研究,以筛选环境友好型植株,并综合考虑沟渠的水体净化功能、温室气体排放量及生态价值。
