CCN3对小鼠胚胎成纤维细胞增殖的影响
2021-01-28陈世钰刘俊平石宇彤吴敏敏徐梦歧张凤梅
陈世钰,苏 欣,刘俊平,石宇彤,吴敏敏,徐梦歧,张凤梅,唐 敏
重庆医科大学检验医学院//临床检验诊断学教育部重点实验室,重庆 400016
骨折是最常见的骨损伤情况之一,数据显示有5-10%的骨折为难治性骨损伤,其主要是由于多种情况导致机体自身骨修复能力下降,这类患者可通过骨组织工程进行治疗[1]。骨组织工程通过补充骨生长因子、移植骨替代品以及干细胞,提高骨缺损部位的骨再生能力,达到骨修复目的[1-2]。其中,间充质干细胞(MSCs)由于具有良好的自我增殖和多向分化能力,是骨组织工程重要种子细胞,但外源性间充质干细胞由于来源有限、增殖衰老和异质性等的问题[3-4],临床应用受限,目前骨组织工程主要采用无MSCs的治疗[1,5],也就是单独或联合使用骨支架和成骨生长因子(BMP2等)进行治疗。然而,大量回顾性研究发现,无MSCs的骨组织工程的临床治疗效果不如预期,同时还因为剂量问题易出现异位成骨、强免疫反应等副作用[1-2,4]。因此,我们思考能否通过促进机体骨损伤部分、即自身的间充质干细胞数量增加,解决骨修复所需种子细胞来源问题,再结合促成骨因子、骨支架,促进难治性骨损伤的愈合呢?为此,我们在前期间充质干细胞成骨分化机制研究基础上[6],探讨细胞因子对MSCs增殖、凋亡的影响和机制,期望能发现能刺激骨损伤部位自身间充质干细胞数量增多的细胞因子,以促进骨组织工程在临床中的应用。
CCN3(NOV/CCN3)是由Ccn基因编码的一种细胞外分泌蛋白,在人体多个组织中均有表达,参与调节细胞粘附、有丝分裂以及肿瘤细胞发生发展等诸多生物过程[7-9]。近年来研究发现CCN3的表达与骨形成和发育密切相关[9-11],且CCN3可作为Notch分泌型配体激活Notch信号通路[12]。课题组前期研究结果发现CCN3参与调控Notch对BMPs诱导的间充质干细胞成骨分化过程[13],然而目前尚未见到关于CCN3对MSCs增殖、凋亡影响的相关报导。
小鼠胚胎成纤维细胞(MEFs)已被证实在体内外均具有MSCs的特性,其作为MSCs研究模型被越来越多的运用在MSCs相关研究中[14-16]。综上,本研究拟以MEFs作为研究对象,利用重组腺病毒过表达及干扰CCN3表达,探讨CCN3对MEFs增殖和凋亡的影响及其潜在作用机制。本研究结果将丰富调控MSCs生长发育机制的基础研究结果,且可为解决骨组织工程中种子细胞来源问题提供新思路。
1 材料和方法
1.1 材料
1.1.1 腺病毒、细胞株 小鼠胚胎成纤维细胞MEFs,人肾上皮细胞系HEK 293细胞由重庆医科大学临床检验诊断学重点实验室保存。过表达绿色荧光蛋白腺病毒(Ad-GFP)、过表达红色荧光蛋白腺病毒(Ad-RFP)、过表达CCN3腺病毒(Ad-CCN3)、干扰CCN3腺病毒(AdsiCCN3)均由芝加哥大学何通川教授馈赠。
1.1.2 主要试剂 DMEM高糖培养基购自重庆赛米克公司;胎牛血清(FBS,太仓依科赛);Polybrene以及茜素红S(Sigma);碱性磷酸酶染色试剂盒、Western blot相关试剂及IP细胞裂解液(碧云天);RNA提取试剂盒(奕杉);RT-PCR试剂盒和SYBR GreenⅠ(TaKaRa);CCN3 ELISA试剂盒以及兔抗鼠ERK1+2、p-ERK1+2抗体(Abcam);PVDF膜以及化学发光HRP底物(Millipore);鼠抗鼠β-actin单克隆抗体(钟鼎);兔抗鼠PCNA、Bax、Bcl-2以及乙酰化H3K9抗体(CST);兔抗鼠Cyclin E、Cyclin B1抗体(万类生物科技);牛血清粉BSA(索莱宝);胰蛋白酶以及羊抗兔、羊抗鼠二抗(中杉金桥);磷酸盐粉(博士德);引物分别于北京擎科公司以及上海生工生物工程公司设计及购买。
1.2 方法
1.2.1 细胞培养 小鼠间充质干细胞MEFs、人肾上皮细胞系HEK 293细胞使用含10%胎牛血清、1%青霉素/链霉素(P/S)的DMEM完全培养基在37℃、5% CO2饱和湿度条件的细胞培养箱中培养。当细胞贴壁融合至70%~80%(对数生长期)时,用0.25%胰蛋白酶消化继续传代。
1.2.2 腺病毒处理细胞 取对数生长期MEFs铺板(24孔板,5×105/孔)检测病毒滴度。观察感染率约为30%的对应孔病毒滴度并在荧光显微镜下拍照。确定病毒滴度后,培养MEFs至30%密度左右,加入Polybrene穿孔素(4µL/500µL培养基),再按所测病毒滴度处理细胞,加病毒6~8 h后更换培养基为无血清无抗生素的DMEM培养基。绿色荧光于处理后24 h在荧光显微镜下观察荧光,红色荧光于处理后36小时在荧光显微镜下观察荧光并拍照。
1.2.3 流式细胞术检测细胞周期及细胞凋亡 利用Ad-GFP、Ad-RFP、Ad-CCN3、Ad-siCCN3腺病毒感染细胞,2d后进行细胞周期及细胞凋亡检测。
细胞周期检测:吸去培养基,胰蛋白酶消化细胞,冰PBS洗涤细胞3次,随后离心沉淀细胞,弃上清,用70%预冷的乙醇重悬,4℃过夜,最后经流式细胞仪,采用碘化丙啶(PI)DNA染色法检测细胞周期情况。
细胞凋亡检测:吸去培养基,胰蛋白酶消化细胞,冰PBS洗涤细胞3次,离心沉淀细胞,弃上清,PBS重悬,最后经流式细胞仪,综合采用Annexin-ⅤAPC-A/DAPI PD450-A双染法以及AnnexinV-FITC单染法检测细胞凋亡情况。
1.2.4 ELISA实验 采用Ad-GFP、Ad-RFP、Ad-CCN3、Ad-siCCN3腺病毒感染细胞48 h后收集细胞培养上清液,2000g离心10 min后取上清液按照试剂盒说明书进行ELISA实验。具体步骤如下:将所有材料和试剂放至室温后稀释样本至试剂盒检测的线性范围内,孔板内加入50µL样本或标准品,随后加入50µL抗体混合物密封,400 r/min摇板室温孵育1 h,350 µL1×洗液/孔,洗涤3次后,加入100µLTMB底物,400 r/min的摇板机避光室温孵育10 min后,每孔加入100µL终止液,振荡混匀1min,酶标仪测定在450nm吸光度。
1.2.5 RNA提取、逆转录和qRT-PCR检测细胞mRNA表达 腺病毒感染细胞后,于培养第3天时利用RNA快提取试剂盒提取RNA,并测定RNA浓度。使用逆转录试剂盒将提取的RNA逆转录为cDNA,之后进行RTPCR实验,并采用SPSS 17.0进行统计学分析。P<0.05表示差异有统计学意义。RT-PCR使用的引物序列见表1。
1.2.6 总蛋白提取和Western blot检测蛋白表达水平总蛋白提取:腺病毒感染细胞后,于细胞培养至第3天时提取蛋白质。弃去培养基,预冷PBS洗3次,向培养板中加1mL预冷PBS,刮下细胞,吸至1.5 mL EP管中,5000 r/min,4 ℃,5min,弃上清,加入细胞蛋白裂解液100µL/管,混匀,每溶10 min震荡1次,共3次,12 000 r/min,4℃,20 min,取上清90µL至新的EP管中,留取10µL测蛋白浓度备用,90µL中加入22.5µL 5×蛋白上样缓冲液,混匀,封口胶封口,沸水浴5~10min,-20℃保存。
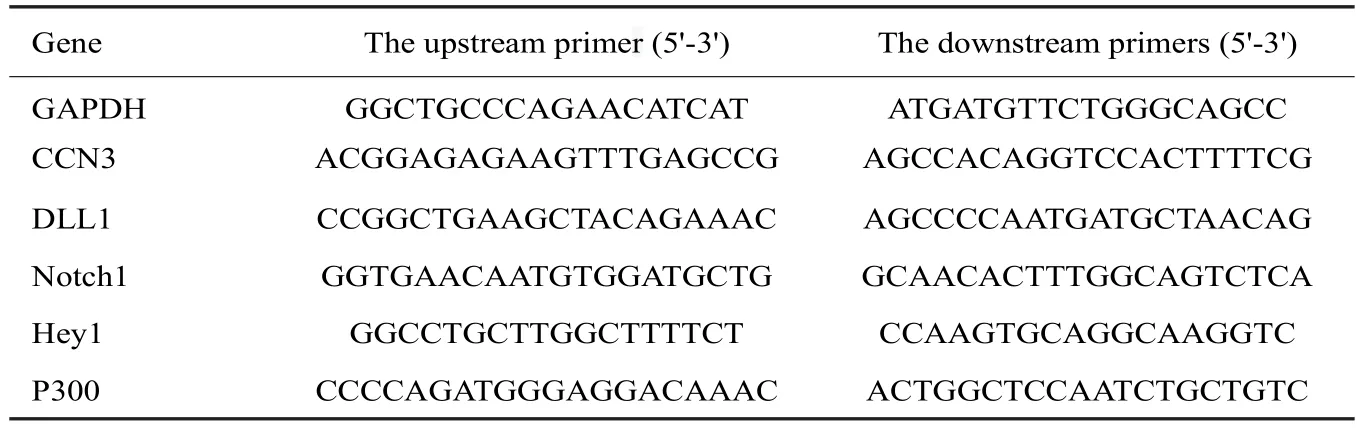
表1 实时定量PCR引物表Tab.1 Primer sequences for real-time quantitative PCR
Western blot实验:根据蛋白相对分子质量配胶,加入适量蛋白进行电泳,恒压浓缩(90 V)和分离(120 V)蛋白,恒流(210 mA)将蛋白转移至PVDF膜上,取出PVDF膜,BSA或5%脱脂牛奶37℃封闭2 h,然后加入相对应的一抗(1∶1000稀释)于4℃放置12~16 h。在震荡仪上用TBST洗膜10 min/次,一共3次。然后加入相对应的山羊抗兔IgG或山羊抗鼠IgG二抗(1∶5000稀释)于37℃孵育1 h,TBST洗膜10 min/次,最后于化学发光成像仪成像。显色结果用Image Lab软件分析。
1.3 统计学分析
各实验均独立重复至少3次,采用GraphPad Prism 5软件进行数据分析,数据以均数±标准差表示,使用单因素方差分析进行多组间比较,t检验进行组间两两比较,检验水准α=0.05。
2 结果
2.1 腺病毒效果鉴定
荧光显微镜下观察,结果显示Ad-GFP、Ad-CCN3加入约24 h后,可在MEF细胞中观察到明显的绿色荧光(图1A左),且RT-qPCR结果表明,与Ad-GFP对照组相比,Ad-CCN3处理组细胞内CCN3表达量明显增高(图1B,P<0.05);Ad-RFP、Ad-siCCN3加入约36 h后,MEF细胞中可以观察到明显的红色荧光(图1A右),RT-qPCR结果显示,与Ad-RFP对照组相比,Ad-siCCN3组CCN3表达量降低(图1C,P<0.001)。

图1 腺病毒感染间充质干细胞情况Fig.1 Mouse embryonic fibroblasts(MEFs)transfected with different adenovirus vectors for CCN3 over-expression or knockdown.A:MEFs with CCN3 over-expression and knockdown under fluorescence microscope.B:CCN3 mRNA expression level in MEFs over-expressing CCN3 detected by RT-qPCR(*P<0.05).C:CCN3 mRNA expression level in MEFs with CCN3 knockdown detected by RT-qPCR(***P<0.001).
2.2 CCN3促进间充质干细胞增殖
与Ad-GFP对照组相比,Ad-CCN3组S期细胞百分比明显增加,增殖指数PI指数明显增加(图2A、B,P<0.01);Ad-siCCN3组,细胞周期无明显变化(图2C、D,P>0.05)。腺病毒感染36 h后提取总蛋白检测经典增殖标志蛋白PCNA、G1/S期周期蛋白标志物Cyclin E和G2/M期标志蛋白Cyclin B1表达,结果显示与Ad-GFP对照组相比,Ad-CCN3组PCNA表达明显增加,Cyclin E表达显著增加,而Cyclin B1表达减少(图2F,P<0.05),Ad-siCCN3组则无明显变化(图2E、F)。
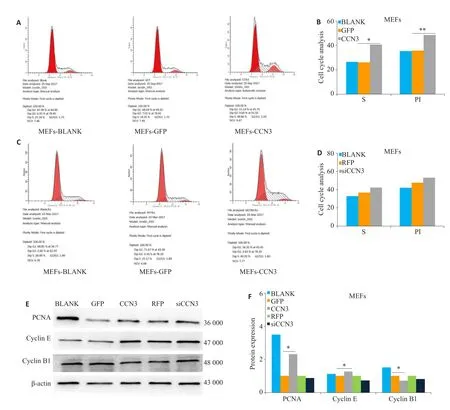
图2 CCN3对细胞增殖的影响Fig.2 Effect of CCN3 over-expression or knockdown on proliferation of MEFs.A,B:Effect of CCN3 overexpression on cell cycle detected by flow cytometry.C,D:Effect of CCN3 knockdown on cell cycle detected by flow cytometry.E,F:Expression levels of PCNA,cyclin E and cyclin B1 in MEFs with CCN3 over-expression or knockdown detected by Western blotting.*P<0.05,**P<0.01.
2.3 CCN3抑制间充质干细胞凋亡
Ad-CCN3组与Ad-GFP对照组相比,细胞凋亡百分比显著降低,其中晚期凋亡细胞百分比降低更为明显(图3A、C,P<0.05);Ad-siCCN3组与Ad-RFP对照组相比,细胞凋亡百分比增加(图3B、D,P<0.05)。随后腺病毒处理36 h后,提取细胞总蛋白检测凋亡抑制蛋白Bcl-2、凋亡促进蛋白Bax表达变化情况。与对照组相比,Ad-CCN3组Bax蛋白表达减少(图3E、F,P<0.05),Bcl-2表达显著增加(图3E、F,P<0.01),Add-siCCN3组Bax表达无明显变化(图3E、F,P>0.05),Bcl-2蛋白表达减少(图3E、F,P<0.01)。
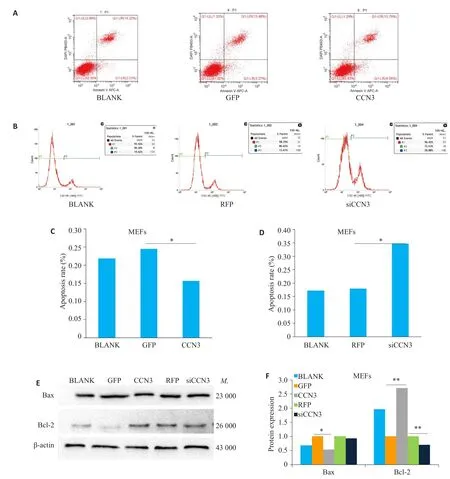
图3 CCN3对细胞凋亡的影响Fig.3 Effect of CCN3 over-expression or knockdown on MEF apoptosis.A,C:The effect of overexpression of CCN3 on apoptosis was detected by flow cytometry(*P<0.05).B,D:The effect of Ad-siCCN3 on apoptosis was detected by flow cytometry(*P<0.05).E,F:The expression levels of pro-apoptotic protein Bax and anti-apoptotic protein Bcl-2 after transfection by CCN3 and siCCN3 viruses detected by Western blotting(*P<0.05,**P<0.01).
2.4 不同处理下CCN3分泌情况
ELISA结果显示:MEFs细胞为低密度时(30%)CCN3浓度在为0.27 ng/mL左右,中等密度(60%)CCN3浓度为0.97~1.05 ng/mL左右,高密度(90%)CCN3浓度为16.04~27.71 ng/mL左右(图4A)。随后ELISA试剂盒检测腺病毒感染后CCN3表达情况结果显示:以高细胞密度组CCN3表达为对照,Ad-CCN3组CCN3浓度增加至849.62~1629.48 ng/mL之间,而siCCN3组CCN3的浓度在9.09~22.5 ng/mL之间,略低于正常水平(图4B)。

图4 不同情况下CCN3表达水平Fig.4 CCN3 secretion level in MEFs cultured under different conditions or with CCN3 over-expression or knockdown.A:CCN3 secretion levels by MEFs cultured at different cell densities(30%,60%,and 90%)detected by ELISA.B:CCN3 secretion levels in MEFs cells with CCN3 over-expression or knockdown detected by ELISA.
2.5 CCN3抑制经典Notch信号通路
与Ad-GFP对照组相比,Ad-CCN3处理组Notch1 mRNA表达增加(图5A,P<0.001),Hey1 mRNA表达减少(图5B,P<0.01),p300 mRNA表达减少(图5C,P<0.05),组蛋白H3K9表达减少(图5D、E,P<0.01);Ad-siCCN3组结果则刚好相反,与Ad-RFP对照组相比,Notch1 mRNA表达减少(图5A,P<0.05),Hey1和p300 mRNA表达明显增加(图5B、C,P<0.05;P<0.01),组蛋白H3K9表达增加(图5D、E,P<0.05)。此外RTPCR结果显示,过表达CCN3能抑制DLL1 mRNA表达(图5F,P<0.01),干扰CCN3可促进DLL1mRNA表达(图5F,P<0.05),而过表达DLL1抑制CCN3 mRNA表达(图5G,P<0.05)。
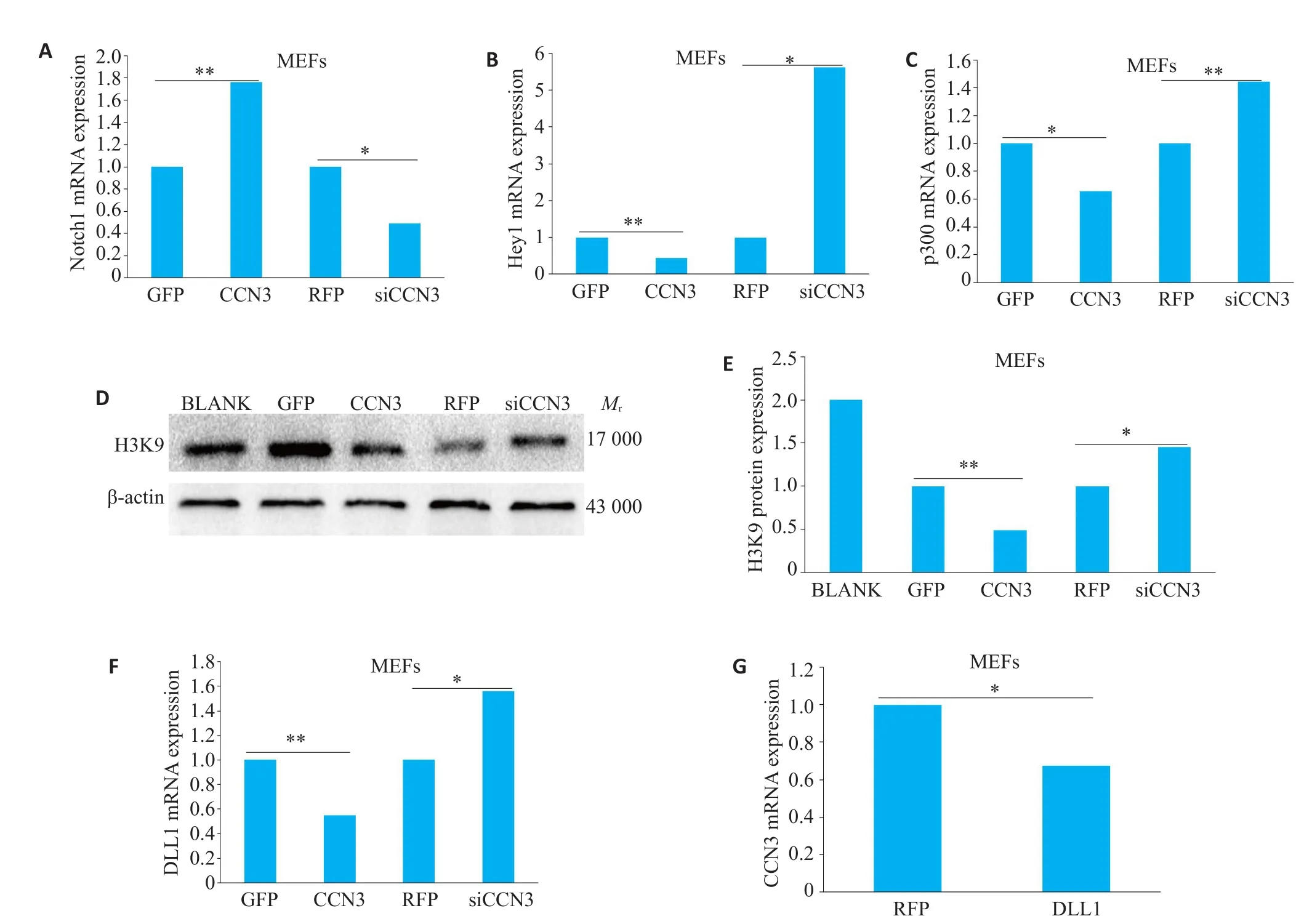
图5 CCN3抑制经典Notch信号通路Fig.5 CCN3 inhibits the classical Notch signaling pathway.A:The influence of Ad-CCN3 and Ad-siCCN3 on the mRNA expression of Notch1 was detected by RT-qPCR(*P<0.05,***P<0.001).B:RT-qPCR was used to detect the effects of overexpression of CCN3 and siCCN3 on Hey1 mRNA expression(*P<0.05,**P<0.01).C:RT-qPCR was used to detect the effects of overexpression of CCN3 and siCCN3 on p300 mRNA expression(*P<0.05,**P<0.01).D,E:The protein expression levels of acetylated histone H3K9 after transfection of CCN3 and siCCN3 viruses determined by Western blot(*P<0.05,**P<0.01).F:RT-qPCR was used to detect the effects of overexpression of CCN3 and siCCN3 on DLL1 mRNA expression(*P<0.05,**P<0.01).G:RT-qPCR was used to detect the effects of overexpression of DLL1 on CCN3 mRNA expression(*P<0.05).
2.6 CCN3激活ERK/MAPK信号通路
与Ad-GFP对照组相比,Ad-CCN3处理组ERK1+2和p-ERK1+2蛋白表达增加(图6,P<0.001,P<0.01);与Ad-RFP对照组相比,Ad-siCCN3组ERK1+2蛋白表达量无明显变化(图6,P>0.05),p-ERK1+2蛋白表达减少(图6,P<0.05)。

图6 CCN3激活ERK/MAPK信号通路Fig.6 CCN3 over-expression activates the ERK/MAPK signaling pathway in MEFs.A,B:Protein expression levels of ERK1+2 and p-ERK1+2in MEFs with CCN3 over-expression or knockdown determined by Western blotting.*P<0.05,**P<0.01,***P<0.001.
3 讨论
近年来骨组织工程作为难治性骨损伤的重要治疗手段,越来越受到临床重视。其通过各种方式促进骨损伤部位的间充质干细胞成骨分化,实现骨修复[17],因此,保障骨损伤部位间充质干细胞的数量对骨修复能力至关重要,然而外源性间充质干细胞由于存在来源有限、异质性以及增殖衰老发育机制不清楚等问题限制了其使用[18-19],因此,拟在前期对间充质干细胞成骨分化的研究基础上,进一步探讨间充质干细胞增殖的调控机制,探索能够促进机体自身间充质干细胞数量增多的方法,为改进骨组织工程治疗策略、提高骨组织工程的临床治疗效果提供思路和依据。
肾母细胞瘤过表达因子CCN3近年来被发现在肿瘤发生发展以及骨再生过程中发挥重要调控作用[20-22]。前期研究发现,DLL1/Notch信号通路能明显促进BMPs调控的间充质干细胞成骨分化[23],而CCN3却明显抑制间充质干细胞成骨分化[13]。因为增殖和分化是细胞功能两个不同的方向,因此,我们考虑CCN3在这一过程中,是否是作为间充质干细胞增殖因子存在。查阅文献,发现有研究结果表明CCN3可促进牙髓干细胞DPSCs以及内皮细胞的增殖[21,24],而也有学者的研究结果所得结论恰好与之相反[25-27],这间接提示CCN3调控细胞增殖机制复杂,值得进一步研究。
本研究利用重组CCN3腺病毒感染细胞,探究CCN3对细胞增殖的影响及潜在机制。结果显示:上调MEFs中CCN3表达,能够显著促进细胞增殖,这与CCN3对Balb/c 3T3成纤维细胞作用中的发现一致[28],而这一过程中,G1/S期标志周期蛋白Cyclin E的表达增加,G2/M期Cyclin B1的表达则是下调的,提示CCN3可能是通过影响细胞G1/S期调控细胞增殖。然而与预期结果不一致的是干扰CCN3表达后对细胞增殖无明显影响。考虑到CCN3是一种分泌型蛋白,其发挥调控作用受到其浓度的影响,且已有研究表明不同浓度CCN3对成骨分化和细胞增殖作用不同[10,28-29],我们进一步检测不同处理情况下CCN3的表达量,ELISA结果显示,MEFs中CCN3的基础表达量约为21.87ng/mL,腺病毒处理后可显著上调细胞CCN3分泌,约为基础表达量的60倍,然而Ad-siCCN3感染后仅在细胞基础表达量上轻微下调CCN3表达,因此我们猜测Ad-siCCN3组结果与预期不符的原因可能由于是腺病毒下调CCN3表达量不足以影响细胞增殖,这一猜想尚需要进一步设计实验验证。研究中,我们还发现过表达CCN3能够抑制MEFs细胞凋亡,提示CCN3同时还通过抑制细胞凋亡的途径增加细胞数量。
那么CCN3是通过什么途径调控MEFs增殖的呢?因为CCN3已经被证实可与Notch1结合,是Notch信号通路的非经典配体之一[12]。于是我们首先检测了CCN3对Notch信号通路的激活,RT-PCR结果显示,过表达CCN3能促进Notch1 mRNA表达,而siCCN3则抑制其表达,证实CCN3的确可激活Notch1。然而进一步研究发现,过表达CCN3明显抑制了Notch经典通路中靶基因Hey1和p300 mRNA表达,Western blot结果也显示组蛋白乙酰化水平明显下降;干扰CCN3后结果刚好相反,提示过表达CCN3虽然激活Notch1,但在功能上却抑制了经典的Notch信号通路。前期研究已经发现Notch信号通路可激活MAPK[30-31],且MAPK通路与细胞增殖密切相关[32-33],因此我们进一步检测了MAPK通路下游蛋白变化情况,Western Blot结果显示,过表达CCN3能够促进ERK1+2以及p-ERK1+2蛋白表达,这提示CCN3可能通过上调MAPK信号通路调控细胞增殖和凋亡。
综上所述,本研究在前期研究基础上,对Notch分泌型配体CCN3在MEFs增殖和凋亡中的作用和机制进行了初步探讨,研究结果显示CCN3可能与Notch1结合,通过抑制DLL1/Notch信号通路,激活MAPK信号通路,促进细胞增殖,抑制凋亡,从而有效增加细胞数量。本结果提示CCN3有望作为MSCs的有效促增殖因子应用于骨组织工程中。但在骨组织工程中,如何合理使用CCN3,与骨形成蛋白BMPs、骨支架等协同作用,促进骨损伤部位骨再生,尚需进一步研究。
