利用CRISPR/Cas9技术编辑AFP1基因提高水稻耐逆性
2021-01-22周天顺余东刘玲欧阳宁袁贵龙段美娟袁定阳
周天顺 余东 刘玲 欧阳宁 袁贵龙 段美娟 袁定阳
(1湖南杂交水稻研究中心/杂交水稻国家重点实验室, 长沙 410125; 2湖南大学 研究生院隆平分院, 长沙 410125; 3湖南省农业科学院, 长沙 410125;4湖南农业大学 农学院, 长沙 410126; #共同第一作者; *通信联系人, E-mail: duanmeijuan@163.com; yuandingyang@hhrrc.ac.cn)
非生物胁迫是阻碍水稻生长、发育和繁殖的主要因素之一[1]。研究表明,干旱、高盐、高温等非生物胁迫可能会导致水稻大面积减产,甚至绝收[2]。而通过挖掘逆境相关基因选育耐逆水稻品种是降低逆境胁迫危害最有效的途径之一。
AFP蛋白(ABI5结合蛋白,ABI5 binding protein)可以通过抑制ABI5活性,参与植物生长发育和多种逆境胁迫响应[3]。研究表明,AtTMAC2(AtAFP4)过表达拟南芥对 ABA和盐的敏感性降低,存活率下降,叶中淀粉积累增多,生育期延迟,根长伸长受到抑制[4]。AtAFP2通过调节 SOM 的表达,GA和 ABA的动态平衡,打破种子由高温所引起的二次休眠[5]。AtAFP2还可形成 CO-AFP2-TPR2蛋白复合体负调控拟南芥的开花时间,AtAFP3和AtAFP4同样影响开花时间,但其机制是否相同有待进一步研究[4,6-7]。OsMODD通过MODD-TPR3-HDA702和MODD-OsOTLD1复合体在干旱恢复期促进OsbZIP46蛋白的降解,防止干旱相关基因的过度表达影响水稻生长发育[8-9]。向日葵HaABRC5基因响应 ABA、干旱和盐胁迫[10]。在拟南芥中异源过表达类GgAFP基因可降低拟南芥种子萌发期的ABA敏感性,并改变ABA相关基因的表达,促进种子萌发[11]。
前期研究中,我们发现水稻AFP家族基因AFP1(MODD)被ABA、干旱等多种激素和非生物胁迫处理诱导表达[15]。为进一步鉴定AFP1在水稻非生物胁迫抗性中的作用,快速创制优异等位基因。我们利用CRISPR/Cas9基因编辑技术对AFP1基因进行定向编辑,以期创制耐逆性提高的突变体材料。
1 材料与方法
1.1 材料与载体
以优异籼型恢复系华占为遗传转化受体材料,所有材料种植于湖南杂交水稻研究中心网室,常规水肥管理。sgRNA表达载体pYLgRNA- U3/U6和敲除载体pYLCRISPR/ Cas9-MH9(MT)由华南农业大学刘耀光课题组馈赠。引物(表 1)合成和测序均委托湖南擎科生物技术有限公司完成。
1.2 靶点设计与载体构建
利用CRISPR-GE(http://skl.scau.edu.cn/)设计AFP1(Os03g0214200)敲除靶点。利用IDT Oligo Analyzer 3.1在线软件(https://eu.idtdna.com/calc/analyzer)计算 GC含量。载体构建参见单子叶敲除载体构建方法[16]。将MT-AFP1载体送至武汉伯远生物科技有限公司进行遗传转化。
1.3 T0代阳性株系获得及突变位点检测
采用CTAB法提取T0代植株的基因组DNA,利用Hpt和Cas9(表 1)进行阳性植株检测。利用AFP1-J扩增靶位点序列进行测序。测序结果为双峰的株系,利用TA克隆+测序确定突变位点。
1.4 T1代无T-DNA株系筛选及脱靶位点分析
以T1代植株DNA为模板,HPT-F/R和Cas9-F/R为引物进行PCR扩增。两者均不能扩出目的条带的株系为无 T-DNA株系,用于下一步分析。利用CRISPR-GE预测脱靶位点。每个株系随机选取 20株,对前3个最高脱靶率的位点进行检测。

表1 本研究所用引物Table 1. Primers used in this study.
1.5 T2代纯合突变体农艺性状调查
水稻完全成熟后,在非边际区域随机调查突变体和野生型(每个株系5株)的株高、有效分蘖数、一次枝梗数、穗长、结实率、千粒重和单株产量。
1.6 afp1突变体耐逆性鉴定
对 4叶期幼苗进行胁迫处理。极速干旱处理:将幼苗直接暴露在空气中直至萎蔫表型;高温处理:将幼苗放置在42℃(湿度为70%)人工气候箱处理至叶片坏死。冷处理:将幼苗转移至 4℃的培养箱中处理2~3 d。处理后正常条件下恢复1周,统计存活率。
40粒发芽的种子转移至装有含150 mmol/L甘露醇、200 mmol/L NaCl的1/2 MS固体培养基中,分别进行渗透胁迫和盐胁迫抗性鉴定,培养7~10 d后,统计 10株幼苗的株高、根长和鲜质量。对照处理采用1/2 MS固体培养基。
随着人民水平的提高,越来越多的人饲养宠物。城市中的宠物,由于其生活空间小,周围的犬只少,公犬获得交配的机会十分的少。如果犬只没有被阉割的话,公犬达到中老年以后(尤其是6岁以后),其发生前列腺脓肿的几率会很大。而且在临床上这几年犬的前列腺脓肿也在增加。在诊断方面,由于医生的水平和医院的仪器设备不同,理论上最好要用B超检查,然后腹腔探查确诊,但国内对于该疾病不是特别重视,误诊、漏诊的几率很大;在治疗方面,由于前列腺的特殊结构,其药物治疗效果(尤其在抗菌方面)不是很好。
1.7 ABA敏感性分析
萌发期:去壳种子消毒处理后,置于含3 μmol/L ABA的1/2 MS固体培养基中培养3~5 d后,统计发芽率。对照处理采用1/2 MS固体培养基。
苗期:去壳种子进行消毒处理后,转移至 1/2 MS固体培养基培养2~3 d,将生长状况一致的发芽种子置于含3 μmol/L ABA的1/2 MS固体培养基中培养7~10 d,统计幼苗的株高、根长和鲜质量。对照处理采用1/2 MS固体培养基。
1.8 离体叶片失水率的测定
取4叶期突变体与野生型的全展叶,每个株系取3片,擦干表面水分后置于室温条件,每隔0.5 h测量叶片质量,直至恒重。叶片失水率(%)=(初始叶片鲜质量-某一时间叶片鲜质量)/初始叶片鲜质量×100%。
2 结果与分析
2.1 AFP1敲除靶点设计及载体构建
为提高突变效率,在AFP1第1外显子靠近起始密码子ATG处设计了两个20 bp的敲除靶点T1和 T2,PAM序列分别为 AGG和 CGG,T1位于ATG下游25-47 bp,T2位于ATG下游117-140 bp(图1-A)。MT-AFP1敲除载体如图1-B。
2.2 afp1突变体敲除特征分析
通过农杆菌介导法获得12株转MT-AFP1载体水稻,检测阳性株系发现,所有植株均能扩增出特异性条带,说明12株均为阳性植株(表2)。突变位点分析表明,8个株系的靶点T1发生突变,9个株系的靶点T2发生突变,T1和T2位点的编辑效率分别为66.67%和75.00%。T1和T2靶点突变基因型类型最多的均为双等位突变,分别为75.00%和44.44%;而嵌合突变仅存在于T2中,概率为11.12%。

图1 利用CRISPR/Cas9技术定向突变OsAFP1基因Fig. 1. Directed mutation of OsAFP1 using CRISPR/Cas9 technology.

表2 T0代突变体突变基因型比率Table 2. Mutant genotype ratios of T0 mutations.

图2 突变体特征分析Fig. 2. Analysis of mutation type.
突变株系中仅存在插入和缺失突变,分别占80.0%和20.0%(图2)。突变类型中绝大部分为插入突变,而插入碱基数均为1 bp,其中插入腺嘌呤(A)和胞嘧啶(C)约为96.96%。90%突变体的突变长度少于5 bp(图2)。
2.3 不含外源基因T2代纯合敲除突变体的获得
通过无外源 T-DNA筛选和突变位点鉴定,我们获得了 6个不含外源基因的纯合突变株系KO1~KO6(图3)。蛋白质序列分析表明,除KO2株系为 31个氨基酸的缺失外,其他突变体株系均发生移码突变,造成蛋白质翻译的提前终止。脱靶分析表明,预测脱靶位点的错配碱基数为 2~5个,脱靶概率为0.136~0.353,KO1~KO6纯合突变体中6个潜在脱靶位点均未发生突变。
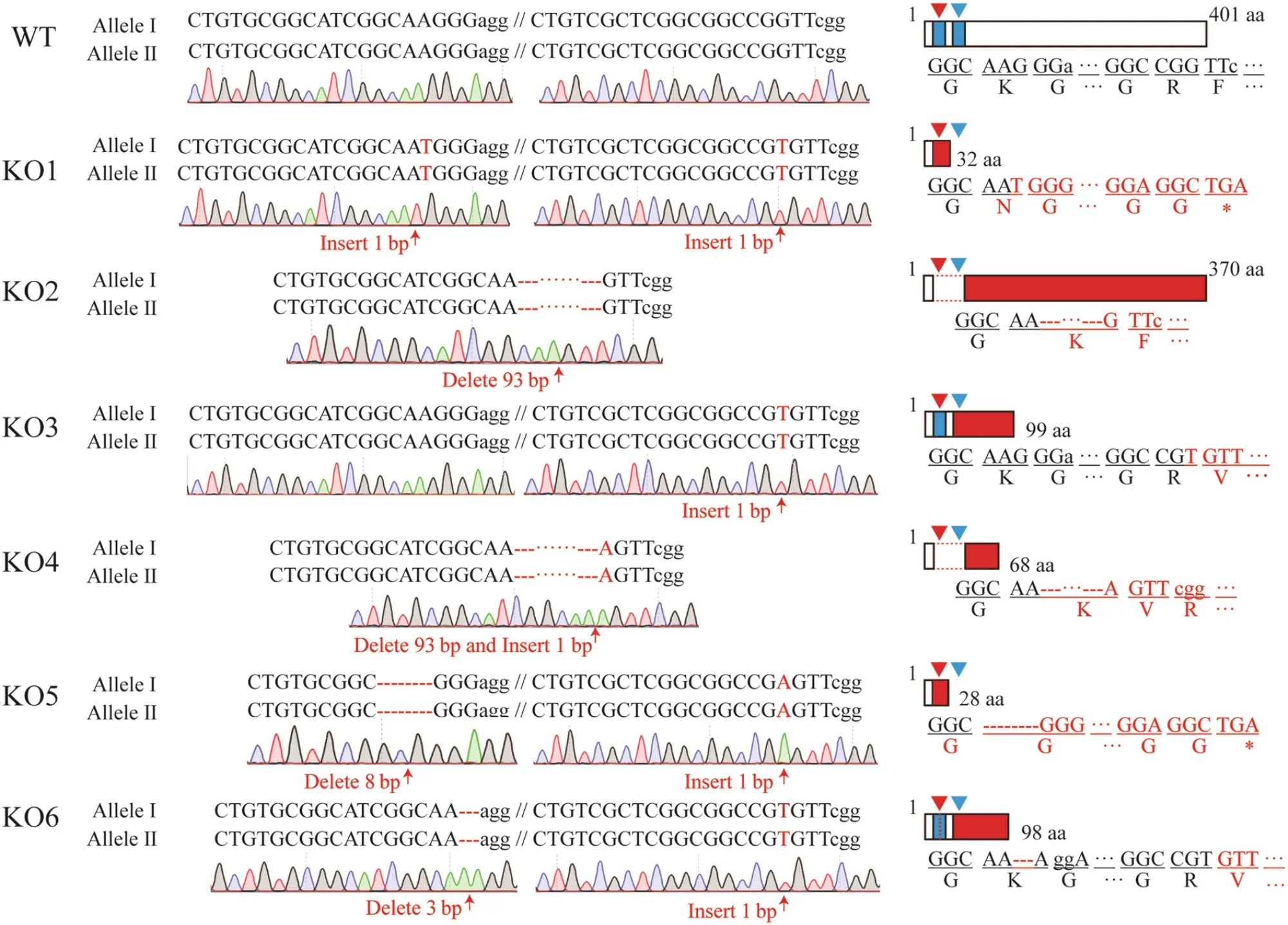
图3 T2代纯合敲除突变体的突变类型Fig. 3. Mutated types of T2 homozygous mutants.
2.4 T2代纯合突变体农艺性状调查
纯合突变体株高有不同程度的降低,生育期提前,有效分蘖增多(图 4)。穗部性状调查表明,突变体穗长显著增加,一次枝梗数和每穗粒数与野生型相当。突变体和野生型的千粒重无显著差异,突变体的结实率下降,但只有 KO3株系达到显著水平。各突变体株系的单株产量呈现不同的趋势,KO1株系的单株产量显著上升,KO2株系的单株产量增加但未达到显著水平,而 KO3株系的单株产量却显著下降。

图4 afp1敲除突变体和野生型的表型比较Fig. 4. Phenotypic comparisons of afp1 knockdown mutants and the wild type.
2.5 afp1突变体ABA敏感性增加
为研究AFP1是否参与ABA信号途径调控,我们进行ABA敏感性测试(图5)。结果表明,3 μmol/L ABA处理后,突变体幼苗的株高、根长和鲜质量均显著低于野生型,种子萌发率也显著降低,正常条件下,野生型和突变体之间却没有明显区别。这些表明敲除AFP1后水稻ABA敏感性升高。
2.6 afp1突变体多种非生物胁迫抗逆性提高
ABA依赖型途径是植物响应干旱的主要信号途径。极速干旱12 h后,afp1突变体和野生型均出现叶片卷曲、植株萎蔫等失水症状,复水10 d后,突变体绿叶面积多,存活率明显高于野生型(图6-A和B),说明afp1突变体耐旱性提高。
大田生产中,高温胁迫往往与干旱胁迫同时出现,严重影响水稻产量。热胁迫处理15 d后,野生型和突变体叶片发白,叶片卷曲,但突变体的绿叶面积和生长状态明显好于野生型,而萎蔫程度低于野生型。以上结果表明,afp1突变体的耐热性提高。
为进一步探讨水稻耐旱和耐热性增强的原因,我们进行了离体叶片水分散失率实验。突变体的水分散失率明显低于野生型,说明突变体的水分散失慢(图6-D)。此外,突变体在甘露醇胁迫下的株高和根长显著长于野生型,根和地上部分的鲜质量显著增高,而正常条件下表型相似,表明afp1突变体的耐渗透胁迫能力增强(图6-C~F)。

图5 野生型和突变体ABA敏感性分析Fig. 5. ABA sensitivity of WT and mutants.
3 讨论
CRISPR/Cas9技术体系中影响编辑效率的因素有很多,如sgRNA设计、sgRNA启动子、Cas蛋白、转化体系等[17]。sgRNA适宜GC含量在30%~80%之间,需具有完整二级结构,且无7个以上连续碱基对[17-18]。靶点1(T1)和靶点 2(T2)的GC含量分别为70%和75%,位于适宜的GC含量范围,最多仅有3个连续相同碱基。编辑效率和GC呈现出一定的正相关关系,T1的编辑效率稍低于T2,可能是由于T1的GC含量低于T2所导致的。插入突变中,90%以上为A或T碱基,具有明显的A/T偏好性[16](图2)。就突变长度而言,突变长度主要集中在5 bp以下的小片段,这与Li等[19]的报道相似(图2)。T1和T2潜在脱靶位点中,均未发现脱靶现象发生。首先,这可能和严格的 sgRNA设计有关[20]。其次,和载体 Cas9蛋白经过优化,识别准确度高、活性好有关[16]。
在同一水肥管理的大田环境下,突变体和野生型的农艺性状表现出一定的差异。afp1突变体和野生型相比的生育期稍微提前,这在拟南芥中也有相似的报道,过表达AtAFP2/3/4均能造成晚花表型,AtAFP2通过CO-AFP2-TPR2复合体调控拟南芥的开花时间[4,6-7]。据报道,AFP1(MODD)可以抑制ABA信号途径,及时减少水稻因抗性提高所带来的能量消耗[8]。afp1突变体的株高有不同程度的下降,这可能是由于敲除AFP1基因后,提高了植物的抗逆性,增加了能量的消耗所导致的(图 4)。虽然突变体穗长显著增加,但是未导致突变体每穗粒数的增加,这可能和一次枝梗数降低相关(图 4)。水稻的产量主要由有效穗数、每穗粒数、结实率和千粒重共同决定。KO1和KO2的单株产量较野生型有所增加,结实率降低,但千粒重和每穗粒数均无明显变化,表明这主要是有效分蘖显著增加所引起的。KO3的单株产量却显著下降,这是由于KO3结实率下降所导致的产量损失大于通过有效分蘖增加的产量。这表明afp1具有育种应用潜力,但需要对突变体材料进行严格筛选,淘汰对产量和其他农艺性状有明显负面影响的株系。

图6 耐旱性和耐渗透性鉴定Fig. 6. Identification of drought and osmotic stress tolerances.
afp1突变体多种非生物胁迫筛选发现,osafp1突变体的耐旱性、耐渗透胁迫能力和耐热性显著提高,但耐盐性和耐寒性没有影响(图6)。拟南芥AFP成员突变体均对ABA超敏感,而osafp1突变体萌发期和苗期ABA敏感性升高,暗示植物中AFP蛋白的功能比较保守。atafp2-4突变体的耐盐性均显著提高,而osafp1突变体在200 mmol/L盐胁迫处理下存活率和生长状态和野生型相似[21-22]。afp1突变体抗逆性增强的原因主要包括两个方面:第一,在干旱胁迫和 200 mmol/L甘露醇处理下,osafp1突变体的根显著长于野生型,这有助于干旱条件下植物的水分吸收,提高自身的水分含量,而另一方面,osafp1突变体的离体叶片失水率显著降低,减少自身的水分散失,从而提高了抗逆性。
