不同八仙花品种对遮阴和强光处理的生理响应与评价
2021-01-22张宪权刘群录
潘 月, 张宪权, 叶 康,3, 刘群录, 秦 俊,3
(1.上海交通大学农业与生物学院,上海200240;2.上海辰山植物园,上海201602;3.上海城市树木生态应用工程技术研究中心,上海200020)
八仙花[Hydrangea macrophylla(Thunb.) Ser.]为虎耳草科(Saxifragaceae)绣球属(HydrangeaLinn.)离瓣组[Sect.Petalanthae (Maxim.) Wei]植物,是我国华东地区广布种[1].八仙花品种繁多,花大色艳,具有很高的观赏价值,其开花时间长达6~8 个月,单花期可达1 ~2 个月.八仙花受到花卉市场的普遍关注,常用于地栽、盆栽、鲜切花及干花制作,在园林绿地中也广泛应用[2-4];同时,八仙花对铝具有较强的耐受能力,可作为工业区土壤的修复植物[5].
光照与园林花木的生长发育状况密切相关[6].研究表明:对芍药(Paeonia lactiflora)进行遮阴处理后,光饱和点(light saturation point, LSP)和光补偿点(light compensation point, LCP)随着遮阴程度的提高而下降,不同品种间存在一定的差异[7];对紫珠(Callicarpa bodinieriLevl.)进行遮阴处理后,表观量子效率(apparent quantum yield, AQY)、暗呼吸速率和LCP 提高,最大净光合速率(maximum net photosynthetic rate, Pnmax)和LSP 随着遮阴程度的提高而显著下降[8].叶绿素在光合作用中起着重要作用.熊静等[9]研究表明,随着光照的减少,朱砂根(Aisia crenata)的叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量逐渐增加,而叶绿素a/叶绿素b 则逐渐下降.遮阴处理影响植物的光合作用,从而影响营养物质的合成,随着光照的减少,植物地上部分通常通过增大冠幅、增加叶面积和减少总叶片数等来获取更多光能[9-12].薛克娜等[13]研究在10%、20%、40%和100%光照下显脉金花茶(Camellia euphlebia)的生长状况,发现显脉金花茶在20%光照下生长状况较好,这表明适度遮阴有利于某些植物的生长发育,过度遮阴会抑制植物生长.叶片中许多可溶性蛋白的合成受光的调控,弱光下的可溶性蛋白含量通常比正常光照条件下的低[9].光照同样影响叶片合成碳水化合物的能力,因而影响着可溶性糖的含量.
八仙花的花期在5—8 月份,在此期间阳光强烈,喜半阴环境的八仙花易患日灼病, 造成叶片发黄甚至焦灼, 影响其开花观赏价值.因此, 露地栽培时八仙花往往需要在稀疏树荫下和林荫道旁栽植, 光照过强时则需遮阴处理[14].开展八仙花光适应性研究和耐光照品种的筛选具有重要的理论和实际意义.在八仙花的光适应性研究方面,徐慧等[15]研究表明,适度遮阴处理能延长八仙花的观赏期,使花朵和花序变大.Kesumawati et al[16]对4 种日本绣球花品种进行不同程度的遮阴处理,发现遮阴处理的植株,其花序会变绿,且光照越弱,花朵的绿色面积越大.蔡建国等[17]研究表明,在遮阴环境下八仙花通过降低LSP 和LCP、提高AQY 等途径来增强对弱光的利用能力.可见,光照会在不同程度上影响八仙花的生长发育特征.在现有有关八仙花的报道中,研究的品种少,测定的指标覆盖面窄,而作为现今受欢迎的花卉,在园林中的运用越来越广泛,其花期在5—8 月份的强烈光照下,筛选耐光照能力强的品种对其在园林中的推广应用具有重要的实际意义.据此,本研究设置全光照、50%光照和25%光照3 个梯度遮阴处理,测定10 个八仙花品种生长发育和光合生理方面的指标,采用主成分分析、隶属函数分析和聚类分析对10 个品种的耐光照能力进行综合评价.
1 材料与方法
1.1 材料
本试验共采用10 个八仙花品种,由杭州画境种业有限公司提供.10 个品种分别为‘珍贵’(H.serrata‘Preziosa’)、‘蓝矮人’(H.macrophylla‘Blauer Zwerg’)、‘康士坦’(H.macrophylla‘Bodensee’)、‘玛蒂尔德’(H.macrophylla‘Mathilde Gütges’)、‘纱织小姐’(H.macrophylla‘Miss Saori’)、‘汉堡’(H.macrophylla‘Hamburg’)、‘无尽夏’(H.macrophylla‘Forever Summer’)、‘花手鞠’(H.macrophylla‘Hanatemari’)、‘完美玛丽斯’(H.macrophylla‘Mariesii Perfecta’)和‘佳澄’(H.macrophylla‘Kasumi’).将10 个品种的1 年生扦插苗栽植于塑料花盆中(口径160 mm、高度175 mm),基质为草炭和珍珠岩(体积比1 ∶4),于上海辰山植物园种质资源圃中进行常规管理,每日2 次定时定量浇水,叶片展开后于晴天9:00—17:00 遮阴,2 个月后,选取长势一致、无病虫害的植株进行后续试验.
1.2 试验设计
遮阴试验在上海辰山植物园种质资源圃中进行,从2019 年5 月中旬展叶后开始,到8 月中旬结束,设置3 个处理:无遮阴网(100%光照)、50%遮阴网(50%光照)和75%遮阴网(25%光照),两种类型的遮阴网为市场常见的遮阴网,遮阴网距离地面1.8 m,模拟自然遮光[7].每个品种每个处理设3 个重复,每个重复3株,处理结束后随机选取植株顶端向下数的第4~5 枚叶片,每个指标重复3 次.
1.3 指标的测定
1.3.1 光响应曲线的测定和光合特征参数的计算 测定时间为2019 年8 月上旬.在连续晴天的条件下,选取充分受光、朝向一致的成熟叶片并进行标记,运用LI-6400 便携式光合仪测定光响应曲线参数.CO2浓度控制在400 μmol·mol-1,叶室温度控制在(33±0.5) ℃,叶室相对湿度控制在(35±5)%,光照强度梯度分别为2 000、1 800、1 500、1 200、1 000、800、600、400、200、150、100、80、60、40、20、0 μmol·m-2·s-1.每片叶测定前先进行20 min 光诱导,诱导光照强度为1 000 μmol·m-2·s-1,然后开始光响应曲线参数自动测量程序[7,18-19],每个品种测定3 株,每株测定3 片叶,每片叶重复测定3 次.
根据测定所得的数据绘制光合—光响应曲线,光响应过程的模拟采用叶子飘等[20-21]的非直角双曲线模型拟合计算出光合特征参数:Pnmax、LCP 和AQY.利用低光条件下的净光合速率(net photosynthetic rate,Pn)和光合有效辐射(photosynthetically active radiation, PAR)进行直线回归分析[22],得到LSP.
1.3.2 形态指标和生物量的测定 于遮阴处理90 d 后测定.测定所有植株的株高;用叶面积仪测定叶面积,所测叶片为自植株顶端向下数的第4 枚成熟叶片,每个处理每个品种随机选取3 株,每株测3 片叶,并将样品于105 ℃烘箱中杀青30 min,80 ℃下烘干4~6 h 至恒重,称取其干重,比叶面积为叶片的单位面积与其干重之比;遮阴处理90 d 后每个品种每个处理选取3 盆植株,分别称取整株叶片和茎干的鲜重,记录总叶数,然后在105 ℃烘箱中杀青30 min 左右,80 ℃下烘干至恒重,称取叶片和茎干部分的干物质量.
1.3.3 生理指标的测定 取新鲜叶片于天平上称取鲜重(W1),然后在105 ℃烘箱中杀青30 min 左右,80 ℃下烘干至恒重,称取干重(W2),重复3 次,取平均值.叶片含水量/%=[(W1-W2)/W1]×100.
处理结束当天采样并用液氮速冻后放入-80 ℃超低温冰箱中保存,每个指标重复3 次,取平均值.避开叶脉剪取0.1 g 叶片于试管中,加入10 mL 95%(质量比)乙醇,避光浸提48 h 直至叶片无颜色,期间摇动2~3 次,取200 μL 提取液至酶标板中,用酶标仪测定其在波长为665 和649 nm 处的光密度(D),重复3 次,取平均值.叶绿素a 含量=(13.95×D665nm-6.8×D649nm)×0.01/0.2;叶绿素b 含量=(24.96×D649nm-7.32×D665nm)×0.01/0.2;总叶绿素含量=(18.16×D649nm+6.63×D665nm)×0.01/0.2.
采用可溶性蛋白含量测定试剂盒(考马斯亮蓝法)测定可溶性蛋白含量,采用可溶性糖含量试剂盒(蒽酮比色法)测定可溶性糖含量.
1.4 数据处理
采用Excel 2013 和SPSS 20.0 软件对试验数据进行方差分析、主成分分析和聚类分析,运用Duncan 法分析差异显著性.采用模糊数学的隶属函数法[23]计算10 个八仙花品种的隶属函数值,来评价10 个品种的光适应性.
隶属函数值(Uij)的计算公式为:当指标与光适应性呈正相关时,Uij=(Xij-Ximin)(Ximax-Ximin);当指标与光适应性呈负相关时,Uij=1-[(Xij-Ximin)(Ximax-Ximin)].式中,Xij为第i个品种第j个指标的测定值,Ximin为j指标中的最小值,Ximax为j指标中的最大值.
各品种综合评价值(D)的计算公式为:
2 结果与分析
2.1 八仙花的光合特性
从表1 可以看出,10 个八仙花品种间的Pnmax存在一定差异, ‘汉堡’和‘完美玛丽斯’的Pnmax明显大于其他品种,‘无尽夏’和‘佳澄’较低.‘花手鞠’的LCP 显著高于其他品种,‘珍贵’和‘无尽夏’次之,‘康士坦’最低;‘花手鞠’的LCP 比‘康士坦’高2.75 倍:表明‘康士坦’在10 个品种中最耐阴.不同品种间的LSP 不同, ‘花手鞠’的LSP 最大,是‘康士坦’的1.51 倍,表明‘花手鞠’对强光的适应性最好,而‘康士坦’对强光的适应性最差.不同品种间AQY 的差异也非常显著,最高值约为最低值的2.97 倍.
2.2 遮阴处理对八仙花光响应曲线参数的影响
不同光照下八仙花的光响应曲线如图1 所示.
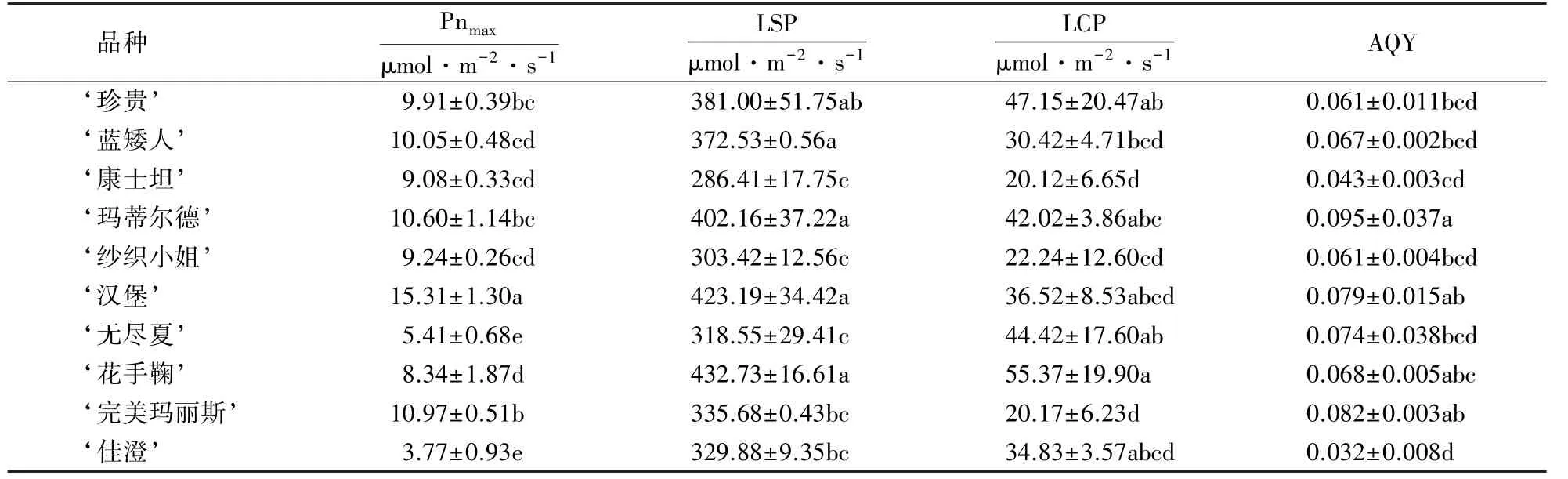
表1 八仙花光响应曲线特征参数1)Table 1 Characteristic parameters of light response curves of H.macrophylla

图1 不同光照下八仙花的光响应曲线Fig.1 Light response curves of H.macrophylla under different light intensities
从图1 可以看出,不同遮阴处理的10 个八仙花品种随着光照的增加,Pn 的变化规律基本相似,即Pn随着PAR 的增大迅速增加,达到一定范围后,Pn 增加减缓,随后趋于平稳.Pn 趋于稳定后为Pnmax,能反映叶片的最大光合能力.
从表2 可以看出:除‘玛蒂尔德’、‘无尽夏’和‘佳澄’外,其他品种的Pnmax均随着光照的减少而下降;除‘玛蒂尔德’外,其他品种的LSP 随着光照的减少逐渐下降;10 个品种的LCP 随着光照的减少全部下降.50%光照下,‘玛蒂尔德’的Pnmax(11.77 μmol·m-2·s-1)和LSP(461.95 μmol·m-2·s-1)最大,表明50%的光照条件更有利于‘玛蒂尔德’的光合作用;50%光照下,‘无尽夏’的Pnmax最大,表明在50%光照下,‘无尽夏’对光能的利用率最大;全光照下,‘佳澄’的Pnmax为3.77 μmol·m-2·s-1,50%和25%光照下的Pnmax分别为5.14、4.86 μmol·m-2·s-1,表明全光照对‘佳澄’的光合特性产生抑制作用.

表2 八仙花光响应曲线参数1)Table 2 Parameters of light response curves of H.macrophylla leaves
2.3 遮阴处理对八仙花生长发育的影响
2.3.1 株高的变化 从图2 可以看出,不同遮阴处理下八仙花株高的变化呈现一定的规律.50%光照下,‘纱织小姐’、‘花手鞠’和‘佳澄’的株高较全光照显著提高,‘珍贵’、‘蓝矮人’、‘汉堡’、‘无尽夏’和‘完美玛丽斯’的株高也大于全光照,但差异不显著;25%光照下,除‘康士坦’和‘玛蒂尔德’外的其他品种的株高均小于50%光照,但差异不显著.‘康士坦’和‘玛蒂尔德’的株高随着光照的减少而下降,‘康士坦’在全光照下的株高(28.79 cm)明显高于50%光照(25.59 cm)和25%光照(25.60 cm),‘玛蒂尔德’3 个处理间的株高差异不显著.不同品种间的株高差异显著.

图2 不同光照下八仙花株高的变化Fig.2 Changes in H.macrophylla plant height under different light intensitiesty
2.3.2 比叶面积的变化 比叶面积能够反映植物适应环境的能力.从图3 可以看出,除‘完美玛丽斯’外,其他品种比叶面积的变化规律一致,均表现为:25%光照>50%光照>全光照.50%光照下,‘蓝矮人’的比叶面积相较于全光照增加幅度最大,增加了101.12%;25%光照下,‘纱织小姐’的比叶面积相较于全光照增加幅度最大,增加了178.31%,‘蓝矮人’的比叶面积相较于全光照增加了170.10%.‘完美玛丽斯’在全光照下的比叶面积为111.64 cm2·g-1,50%光照下的比叶面积(109.26 cm2·g-1)略有下降,但差异不显著,25%光照下的比叶面积显著增加,为203.24 cm2·g-1.
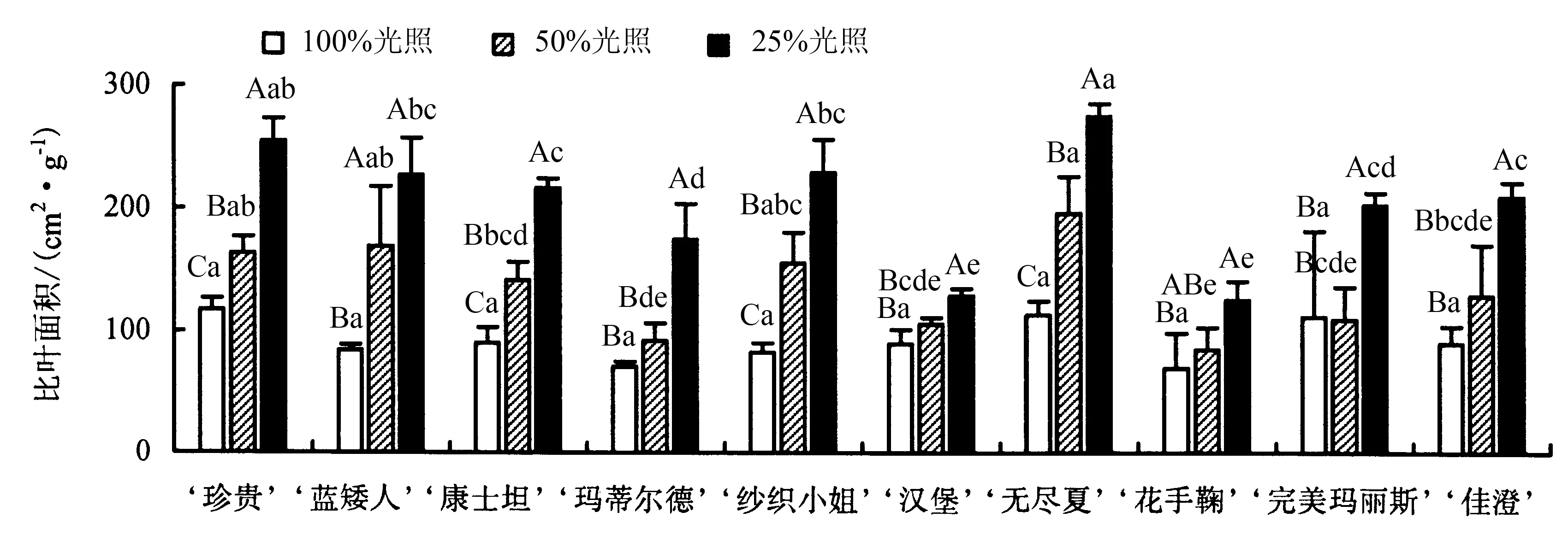
图3 不同光照下八仙花比叶面积的变化Fig.3 Changes in specific leaf area of H.macrophylla under different light intensities
2.3.3 总叶数的变化 从图4 可以看出:50%光照下,‘珍贵’、‘玛蒂尔德’和‘佳澄’的总叶数比全光照下的总叶数多,但差异不显著,其他品种的总叶数有不同程度的减少;25%光照下,‘珍贵’的总叶数仍比全光照多,但差异不显著,其他品种的总叶数比全光照少.

图4 不同光照下八仙花总叶数的变化Fig.4 Changes in total leaf number of H.macrophylla under different light intensities
2.3.4 生物量的变化 表3 显示,10 个八仙花品种叶和茎的鲜重与干重及地上总生物量随着光照的减少呈下降趋势.3 个遮阴处理间,除‘珍贵’和‘佳澄’外,其他品种叶片鲜重的差异均达到显著水平.全光照下,10 个品种的叶鲜重为136.64~234.71 g,‘珍贵’、‘无尽夏’和‘佳澄’的叶鲜重显著低于其他品种;50%光照下,10 个品种的叶鲜重为100.71~187.28 g,品种间的差异不显著;25%光照下,10 个品种的叶鲜重为98.65~159.69 g,品种间的差异亦不显著.随着光照的减少,除‘珍贵’、‘无尽夏’、‘完美玛丽斯’和‘佳澄’外,其他品种的叶干重显著下降,不同品种间的差异显著.除‘康士坦’、‘纱织小姐’和‘汉堡’外,不同遮阴处理对其他品种茎鲜重的影响不显著,品种间的差异显著.不同遮阴处理对茎干重的影响显著, 50%光照下大部分品种的茎干重显著低于全光照下的茎干重,‘珍贵’、‘无尽夏’、‘花手鞠’和‘完美玛丽斯’的茎干重在3 个遮阴处理间的变化不显著,同一光照不同品种间的茎干重差异显著.各品种地上总生物量的变化规律与叶、茎干重的变化规律一致,随着光照的减少呈逐渐下降的趋势,仅‘蓝矮人’和‘无尽夏’的变化规律不同,‘蓝矮人’地上总生物量的变化由高到低依次为全光照、25%光照、50%光照,‘无尽夏’依次为50%光照、全光照、25%光照.

表3 不同光照下八仙花生物量的变化1)Table 3 Changes in H.macrophylla biomass under different light intensities
2.4 遮阴处理对八仙花生理指标的影响
2.4.1 叶片含水量的变化 从图5 可以看出,10 个八仙花品种的叶片含水量随着遮阴程度的增大呈上升趋势,除‘珍贵’、‘花手鞠’和‘完美玛丽斯’外,其他品种3 个处理间的差异均达到显著水平.50%光照下,‘纱织小姐’的叶片含水量相对于全光照上升了8.29%,上升幅度最大,‘完美玛丽斯’的叶片含水量相对于全光照下降了2.01%,但差异不显著.25%光照下,同样是‘纱织小姐’的叶片含水量上升幅度最大,达到12.06%,‘完美玛丽斯’上升幅度最小(4.69%),差异不显著.
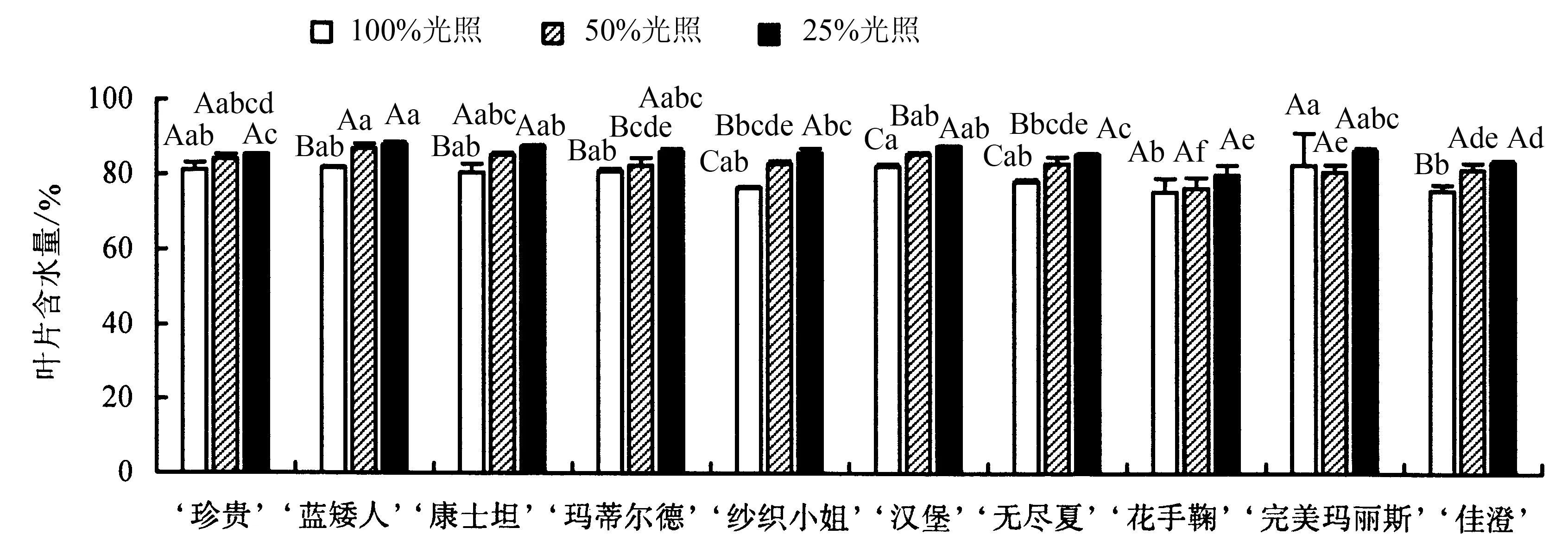
图5 不同光照下八仙花叶片含水量的变化Fig.5 Changes in H.macrophylla leaf water content under different light intensities
2.4.2 叶绿素含量的变化 图6 ~9 显示,10 个八仙花品种的叶绿素a、叶绿素b、总叶绿素的含量及叶绿素a/叶绿素b 随着光照的减少而依次递增.

图6 不同光照下八仙花叶绿素a 含量的变化Fig.6 Changes in chlorophyll a content of H.macrophylla under different light intensities

图7 不同光照下八仙花叶绿素b 含量的变化Fig.7 Changes in chlorophyll b content of H.macrophylla under different light intensities
从图6 可以看出,除‘蓝矮人’、‘玛蒂尔德’和‘完美玛丽斯’外,其他品种的叶绿素a 含量随着光照的减少逐渐增加.25%光照下,‘蓝矮人’和‘玛蒂尔德’的叶绿素a 含量显著高于全光照,但50%光照下的叶绿素a 含量低于全光照.50%光照下,‘完美玛丽斯’的叶绿素a 含量显著高于其他2 个处理.50%光照下,除‘蓝矮人’、‘玛蒂尔德’和‘佳澄’外,其他品种的叶绿素a 含量相较于全光照显著提高.25%光照下,所有品种的叶绿素a 含量与全光照下的差异显著,与50%光照下的叶绿素a 含量相比,除‘珍贵’、‘汉堡’、‘无尽夏’和‘完美玛丽斯’外,其他品种均显著提高.
从图7 可以看出,遮阴处理下各品种叶绿素b 含量的变化趋势与叶绿素a 含量的变化一致.除‘玛蒂尔德’外,其他品种的叶绿素b 含量随着光照的减少而逐渐提高.50%光照下,‘玛蒂尔德’的叶绿素b 含量比全光照下降了10.77%,但差异不显著,25%光照下的叶绿素b 含量比全光照提高了17.63%.50%光照下,除‘蓝矮人’、‘玛蒂尔德’和‘佳澄’外,其他品种的叶绿素b 含量与全光照相比显著提高;25%光照下,所有品种相较于全光照显著提高,相较于50%光照,除‘无尽夏’和‘完美玛丽斯’外的其他品种均显著提高.
从图8 可以看出:10 个品种的叶绿素a/叶绿素b 为1.13~1.68,其中,‘珍贵’的比值最低;在全光照、50%光照和25%光照下,50%光照下的比值最高.不同光照下各品种间的变化趋势基本一致,叶绿素a/叶绿素b 随着光照的减少而提高,除‘玛蒂尔德’和‘完美玛丽斯’外,其他品种的差异均达到显著水平,表明八仙花各品种在遮阴条件下叶绿素b 含量相对于叶绿素a 含量明显提高.

图8 不同光照下八仙花叶绿素a/叶绿素b 的变化Fig.8 Changes in chlorophyll a/chlorophyll b of H.macrophylla under different light intensities
从图9 可以看出:遮阴处理下八仙花各品种总叶绿素含量的变化趋势与叶绿素a 含量、叶绿素b 含量的变化基本一致;除‘玛蒂尔德’外,其他品种的总叶绿素含量随着光照的减少逐渐提高,且差异均达到显著水平.50%光照下,‘玛蒂尔德’的总叶绿素含量比全光照下降了8.41%,但差异不显著,25%光照下的总叶绿素含量比全光照提高了23.70%.50%光照下,除‘蓝矮人’、‘玛蒂尔德’和‘佳澄’外,其他品种的总叶绿素含量相较于全光照显著提高;25%光照下,所有品种的总叶绿素含量相较于全光照显著提高,与50%光照相比,除‘珍贵’、‘无尽夏’和‘完美玛丽斯’外,其他品种的总叶绿素含量均显著提高.
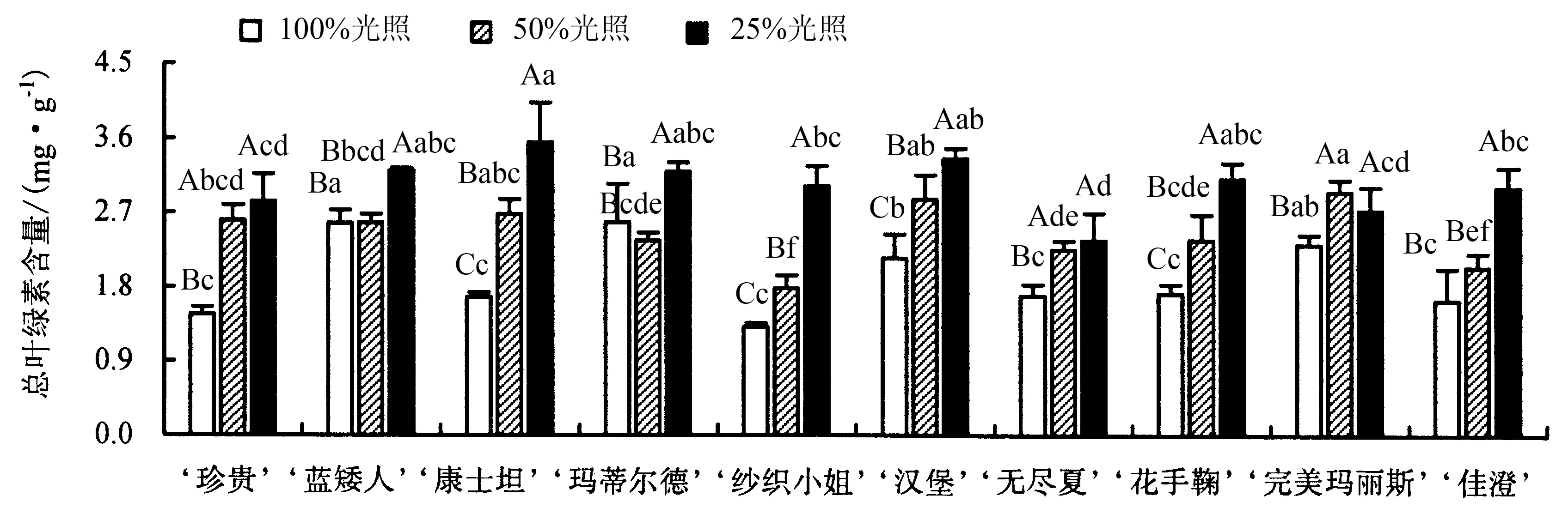
图9 不同光照下八仙花总叶绿素含量的变化Fig.9 Changes in total chlorophyll content of H.macrophylla under different light intensity
2.4.3 可溶性蛋白含量的变化 由图10 可知,全光照下,‘花手鞠’的可溶性蛋白含量最高,为5.53 mg·g-1,‘完美玛丽斯’和‘蓝矮人’次之,分别为4.64、4.62 mg·g-1,‘汉堡’最低,仅为2.03 mg·g-1,其他品种为3.00~4.20 mg·g-1.不同遮阴处理对八仙花各品种可溶性蛋白含量的影响不太一致.50%光照下,‘珍贵’、‘玛蒂尔德’、‘无尽夏’、‘完美玛丽斯’和‘佳澄’的可溶性蛋白含量较全光照均有不同程度的增加,其他品种的含量却下降;25%光照下,除‘玛蒂尔德’、‘花手鞠’、‘完美玛丽斯’和‘佳澄’外,其他品种的可溶性蛋白含量较全光照有不同程度的增加,表明重度遮阴能促进部分品种可溶性蛋白质的合成,提高代谢活动.
2.4.4 可溶性糖含量的变化 由图11 可知,在50%和25%光照下,除50%光照下的‘纱织小姐’外,其他品种的可溶性糖含量相较于全光照均有所下降.50%光照下,‘汉堡’的可溶性糖含量相较于全光照下降幅度最大,达54.53%,其次是‘花手鞠’(下降52.71%)和‘珍贵’(下降52.23%),‘佳澄’的下降幅度最小,仅下降了13.87%,而‘纱织小姐’则上升了42.26%,其他品种下降幅度为17.96%~40.70%,除‘蓝矮人’和‘佳澄’外,其他品种的差异均达到显著水平.25%光照下,除‘蓝矮人’相对于全光照下降了1.84%,但差异不显著,其他品种的可溶性糖含量均显著下降,下降幅度为36.28%~58.52%.
2.5 10 个八仙花品种光适应性的综合评价
2.5.1 光适应性生理生化指标的筛选 10 个八仙花品种在不同光照处理下的形态指标和光合生理指标的变化幅度均不相同,用100%、50%和25%光照下的各项指标来评价各品种光适应性的结果均不相同,因此,数据分析结合研究目的选取具有代表性的10 个八仙花品种在全光照下的测量数据进行光适应性综合评价.
利用SPSS 20.0 软件对八仙花的形态指标和光合生理指标进行标准化处理,对标准化后的指标进行主成分分析.结果(表4)显示,八仙花4 个主成分的累计贡献率达到了88.39%.第1 主成分的贡献率为36.76%,包括叶绿素a 含量、叶绿素b 含量、叶绿素a/叶绿素b、总叶绿素含量和叶鲜重5 个指标,主要反映遮阴处理对叶绿素和生物量的影响;第2 主成分的贡献率为22.54%,包括叶干重、地上总生物量和叶片含水量3 个指标,主要反映遮阴处理对形态指标的影响;第3 主成分的贡献率为18.51%,包括茎鲜重和茎干重2 个指标,主要反映遮阴处理对形态指标的影响;第4 主成分的贡献率为10.58%,包括LSP 和LCP 2个指标,主要反映遮阴处理对光合特性的影响.综上所述,八仙花的叶绿素a 含量、叶绿素b 含量、叶绿素a/叶绿素b、总叶绿素含量、叶鲜重、叶干重、茎鲜重、茎干重、地上总生物量、叶片含水量、LSP 和LCP 等指标的荷载量较大,可作为八仙花光适应性的鉴定指标.根据各指标间相关系数的大小,删减相关系数达到0.85 以上的重复性指标[24],最终选取叶绿素a 含量、叶鲜重、叶干重、茎鲜重、地上总生物量、叶片含水量、LSP 和LCP 等8 个指标计算综合指标值.
2.5.2 光适应性综合隶属函数分析 将光适应性筛选出来的指标值标准化后运用隶属函数法得到10 个八仙花品种的隶属函数值,该值反映了品种耐光照能力的强弱,数值越大,表示耐光照能力越强.对隶属函数值进行排序,耐光照能力(表5)表现为:‘玛蒂尔德’>‘汉堡’>‘完美玛丽斯’>‘花手鞠’>‘蓝矮人’>‘佳澄’>‘无尽夏’>‘珍贵’>‘康士坦’>‘纱织小姐’.
进一步采用组间连接的方法进行系统聚类分析,对隶属函数值进行分类,从而得到所有品种耐光照能力的分类结果.从图12 可以看出,在距离为2.5 处可将10 个八仙花品种耐光照能力的级别分为5 类.第1类仅有‘玛蒂尔德’,耐光照能力最强;第2 类为‘汉堡’、‘完美玛丽斯’和‘花手鞠’,具有较强的耐光照能力;第3 类为‘蓝矮人’和‘佳澄’,耐光照能力一般;第4 类为‘珍贵’和‘无尽夏’,具有较弱的耐光照能力;第5 类为‘康士坦’和‘纱织小姐’,不耐光照.

表5 八仙花耐光照能力的隶属函数值Table 5 The membership function value of strong light resistance of H.macrophylla
3 讨论
3.1 八仙花的光合特性
LSP 和LCP 能够反映植物叶片对强光和弱光的利用能力,阴性植物的LSP 为50~1 000 μmol·m-2·s-1或更低(<20 μmol·m-2·s-1),阳性植物的LCP 为50~100 μmol·m-2·s-1[7].本研究结果表明:八仙花的LSP 为194.91~461.95 μmol·m-2·s-1,LCP 为1.71 ~55.37 μmol·m-2·s-1,更接近阴性植物;LSP 大的品种对强光的适应性强,10 个品种对强光的适应性大小为:‘花手鞠’>‘汉堡’>‘玛蒂尔德’>‘珍贵’>‘蓝矮人’>‘完美玛丽斯’>‘佳澄’>‘无尽夏’>‘纱织小姐’>‘康士坦’,与所有指标计算出的隶属函数值的排序基本一致,表明LSP 对八仙花品种耐光照能力的判断具有一定的可参考性.
AQY 反映植物对弱光的利用能力,在适宜的生长条件下,一般植物的AQY 为0.030 ~0.050[25],八仙花的AQY 为0.032 ~0.103,表明八仙花对弱光的利用能力较强于一般植物.
随着光照的不同,八仙花光响应特性也随之发生变化.全光照下,八仙花的LSP 较高,为286.41 ~432.73 μmol·m-2·s-1,LCP 为20.12 ~55.74 μmol·m-2·s-1,‘花手鞠’对强光的利用能力最大,其次是‘汉堡’,‘康士坦’对强光的利用能力最小.50%光照下,八仙花的LSP为215.20~461.95 μmol·m-2·s-1,LCP 为12.00 ~31.49 μmol·m-2·s-1,除‘玛蒂尔德’外,其他品种的LSP 和LCP 均比全光照低,表明在遮阴环境下,八仙花通过降低LCP 和LSP 来提高对弱光的利用能力;‘玛蒂尔德’、‘无尽夏’和‘佳澄’的Pnmax相较于全光照上升,表明这3 个品种在50%光照下的光合能力强于全光照.25%光照下,八仙花的LSP 为194.91~271.71 μmol·m-2·s-1,LCP 为1.71~19.01 μmol·m-2·s-1,每个品种的光适应范围明显低于全光照,LSP 显著低于全光照,LCP也低于全光照,但部分品种间的差异不显著,所有品种的Pnmax均低于全光照,表明25%光照对八仙花的光合作用产生了抑制作用.

图12 10 个八仙花品种聚类树状图Fig.12 Cluster analysis of 10 cultivars of H.macrophylla
3.2 形态指标、生物量、叶片含水量与八仙花光适应性的关系
遮阴处理使植物的生长环境产生变化,温度下降、空气相对湿度和CO2浓度的提高导致植物形态特征发生变化来适应环境的改变[7].本研究中,随着光照的减少,大部分八仙花品种的株高表现出先提高后下降的变化趋势,表明适度遮阴对八仙花苗高生长量有一定的促进作用,仅‘康士坦’和‘玛蒂尔德’的株高随着光照的减少而下降.唐相红[26]对红厚壳幼苗进行不同程度的遮阴处理后,苗高和地径随着光照的减少出现了先提高后下降的趋势.前人研究表明,在光照较低的环境中,植物常通过增大叶面积,提高叶片含水量来提高光能捕获能力,以弥补光照不足,确保营养生长的正常进行[9,11].本研究结果显示,‘完美玛丽斯’比叶面积的大小表现为:25%光照>全光照>50%光照,其他品种则表现为:25%光照>50%光照>全光照.随着光照的减少,八仙花的叶片含水量逐渐上升,而总叶数、叶和茎的鲜重与干重逐渐下降,表明八仙花通过调整地上部的生物量来适应弱光环境.
3.3 叶绿素与八仙花光适应性的关系
叶绿素含量与植物对光能的利用状况有关.本研究中,随着光照的减少,叶绿素a、叶绿素b 和总叶绿素的含量逐渐增加,与薛欢[27]、吕彬洋等[28]的研究结果一致.在遮阴条件下,以散射光为主,蓝光占比提高[29],叶绿素b 含量显著增加,能够有效吸收蓝光,捕捉更多光能,同时增加叶绿素a 含量,对捕捉的光能进行固定、贮藏[9],提高光能的利用效率.本研究中,叶绿素a/叶绿素b 随着光照的减少而增大,与前人的研究结果[28,30]不同,具体原因有待进一步研究.
3.4 渗透调节物质与八仙花光适应性的关系
光合作用产物主要是糖类,淀粉和可溶性糖是重要的初生代谢产物,植物组织中的可溶性糖和可溶性蛋白都是植物体内重要的渗透调节物质[31].本研究中,不同八仙花品种的可溶性蛋白含量随着光照变化的规律各不相同.除‘玛蒂尔德’、‘花手鞠’、‘完美玛丽斯’和‘佳澄’4 个品种外,其他品种可溶性蛋白含量的大小表现为:25%光照>全光照>50%光照下.造成这种现象的原因可能是,25%光照下,植物体内蛋白质的合成受到抑制,降解加速,使得游离氨基酸含量上升,从而导致可溶性蛋白含量提高[30].可溶性糖含量随着光照的减少而表现出下降的趋势,这与刘玉兰等[32]的研究结果一致,即光照减弱,糖分积累减少.
