FMO3调控犊牛原代肝细胞VLDL-TG的代谢
2021-01-19邓清华宋景旭张玉明刘国文
邓清华,宋景旭,张 焱,刘 婷,张玉明,刘国文,
(1.内蒙古民族大学动物科学技术学院,内蒙古 通辽 028000;2.吉林省松原市前郭县医院,吉林 松原 138000;3.吉林大学动物医学学院,吉林 长春 130062)
我国奶牛脂肪肝的发病率高于30%。脂肪肝影响奶牛的产奶量及乳品质,且常诱发皱胃变位、胎衣不下及生产瘫痪等围产期的其他疾病,给奶牛业带来严重的经济损失[1]。围产期奶牛的能量代谢特点是干物质摄入减少而需求增加所导致的能量负平衡[2]。能量负平衡引发脂肪动员并引起血液中非酯化脂肪酸(Non-eserified fatty acid,NEFA)浓度增加[3]。NEFA 仅有20%能被乳腺组织吸收利用,其余大部分进入肝脏[4-5]。过多的NEFA 在肝脏中代谢生成甘油三酯(Triglyceride,TG),以极低密度脂蛋白(Very low density lipoprotein,VLDL)的形式转运出肝脏,被外周组织利用[6]。NEFA增多不仅加强了促炎因子的活性,还加重胰岛素抵抗,形成恶性循环,最终加剧脂肪肝的发生[7]。由于反刍动物肝脏缺乏足量的脂蛋白脂酶和肝脂酶,通过水解氧化以清除TG的途径受到明显限制,VLDL 的分泌就成为肝脏清除TG 的主要途径。当脂肪动员产生过量的NEFA 被酯化生成TG 而缺乏足够多的VLDL 时,TG 就会积累在肝脏中而引发脂肪肝[8]。VLDL 的组装与分泌主要由载脂蛋白B(Apolipopro⁃tein B,ApoB)100、载脂蛋白E(Apolipoprotein E,ApoE)、微粒体甘油三酯转运蛋白(Microsome tri⁃glyceride transfer protein,MTTP)等蛋白的参与[9]。
含黄素单加氧酶3(Flavin containing monooxy⁃genase 3,FMO3)作为肝脏中的一类微粒体酶,能参与外源性物质的代谢,是动物肝脏中参与非营养物质代谢和转化的重要蛋白[10]。此外,FMO3可通过调节与肝脏脂肪生成和糖异生有关的多个基因来控制血脂的形成[11]。近年来,国外对于FMO3参与调控脂代谢研究较多。在敲除LDL 受体缺陷小鼠肝脏FMO3 基因后,可降低小鼠肝脏和血浆脂质水平、胆汁酸的浓度以及肝脏TG分泌水平;而当PPARα被激活时,可通过其靶基因的反式激活增加FMO3的表达,促进脂肪酸氧化和糖异生[12]。研究表明,FMO3参与肝脏脂质代谢的过程。因奶牛的代谢特征比较特殊,肝脏脂肪代谢的过程存在一定差异,目前尚无关于FMO3 与奶牛脂肪肝和对VLDL 组装和分泌调节的相关文献,所以对于FMO3在奶牛脂肪肝以及调节VLDL组装和分泌方面的研究可为阐明疾病的发病机理及疾病的防治提供参考。
1 材料与方法
1.1 试验动物
1日龄健康荷斯坦犊牛。
1.2 主要试剂
Abcam 牛极低密度脂蛋白ELISA 测定试剂盒;Abcam 牛甘油三酯ELISA 测定试剂盒;天根2×TaqPCR Mastermix;碧云天细胞裂解液(PMSF);TaKaRa DL2000 DNA Marker和逆转录试剂盒;Roche荧光定量试剂。
1.3 试验方法
1.3.1 肝细胞的分离与培养
1日龄的禁食犊牛以无菌方法取出肝脏尾状突。用两步灌流法分离犊牛原代肝细胞[13]。将分离的肝细胞按每孔1×106个细胞放入6 孔细胞培养板,在37 ℃,5%CO2下进行培养。待细胞贴壁后更换培养液,培养72 h后进行后续操作。
1.3.2 腺病毒载体的构建及感染肝细胞
根据GenBank 中牛FMO3基因的编码序列,设计筛选有效的shRNA 并且合成FMO3 基因全序列,构建FMO3基因的低表达和过表达腺病毒并携带绿色荧光蛋白,培养HEK293细胞进行病毒包装、扩增和纯化,所得重组腺病毒测定其滴度,并达到109PFU·mL-1开始收集病毒。将体外培养的肝细胞贴壁培养72 h后用低表达、过表达和空腺病毒载体感染,设空白对照组(Control),低表达FMO3腺病毒载体组(LOW),过表达FMO3 组(OVER)和阴性对照组(NC,腺病毒空载体),每组6 个重复孔,病毒感染复数(MOI)设为100[5],继续培养48 h 后收集细胞及上清液置于-80 ℃。
1.3.3 实时荧光定量PCR
细胞感染48 h 后用Trizol 收集细胞,提取RNA,反转录成cDNA。根据Genbank 中牛的FMO3,ApoB100,MTTP,ApoE,β-actin基因序列,用Primer5软件设计引物,引物序列见表1。用StepOnePlus 荧光定量PCR 检测FMO3、ApoB100、ApoE和MTTP的mRNA相对表达量。

表1 qRT-PCR相关引物
1.3.4 VLDL和TG的收集与检测
VLDL 收集:细胞培养48 h后,将细胞培养液收集到无菌离心管中,1000 g离心20 min,取上清液,用于后续的测定。
TG 提取:细胞培养48 h 后,用细胞刮收集于离心管中,按照碧云天细胞裂解液说明书加入细胞裂解液150 μL,待细胞充分裂解后14000 g离心5 min,取上清液用于检测。
2 结果与分析
2.1 细胞培养及感染腺病毒结果
犊牛原代肝细胞分离培养72 h 后,状态良好,适合感染腺病毒(见图1A)。感染病毒后用荧光共聚焦显微镜观察感染率,研究表明,当MOI=100时,感染效率>80%(见图1B和1C)。
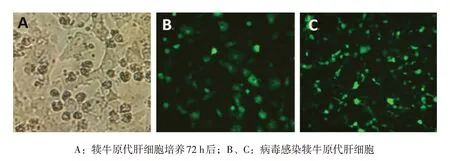
图1 犊牛原代肝细胞和重组腺病毒感染犊牛原代肝细胞荧光共聚焦结果(400×)
2.2 FMO3、ApoB100、MTTP和ApoE基因的mRNA表达量
犊牛原代肝细胞感染低表达、过表达和空载体48 h后用qRT-PCR检测FMO3,ApoB100,ApoE和MTTP基因的mRNA 表达量结果见图3A~D。其中,Control 为空白对照组,LOW 为低表达FMO3腺病毒组,OVER为过表达FMO3腺病毒组,NC为腺病毒空载体组,阴性对照。数值用“平均值±标准误”表示(n=6),不同字母代表差异显著(P<0.01);相同字母代表差异不显著(P>0.05)。

图2 犊牛原代肝细胞感染低表达、过表达和空载体48 h后用qRT-PCR检测FMO3、ApoB100、ApoE和MTTP基因的mRNA表达量
由图2可知,当细胞感染FMO3 低表达腺病毒时,FMO3、ApoB100 和ApoE的mRNA 水平显著降低(P<0.01),MTTP基因的表达量与空白对照组相比差异不显著(P>0.05);当细胞感染FMO3过表达腺病毒时,FMO3、ApoB100、ApoE和MTTP基因的mRNA 水平显著增加(P<0.01),NC 组与Control组相比,各基因mRNA水平差异不显著(P>0.05)。
2.3 细胞上清VLDL和细胞内TG浓度测定结果
根据Abcam 牛VLDL 和TG ELISA 试剂盒检测细胞上清VLDL 和细胞内TG 的浓度。结果表明,FMO3 低表达组(LOW)VLDL 的浓度显著低于空白对照组(P<0.01),高表达组(OVER)显著高于空白对照组(P<0.01),阴性对照组(NC)也显著高于空白对照组(P<0.01,见表2)。低表达组与空白对照组相比细胞内TG 含量明显降低(P<0.01),过表达组显著高于空白对照组(P<0.01),阴性对照组与空白对照组相比无显著差异(P>0.05,见表3)。
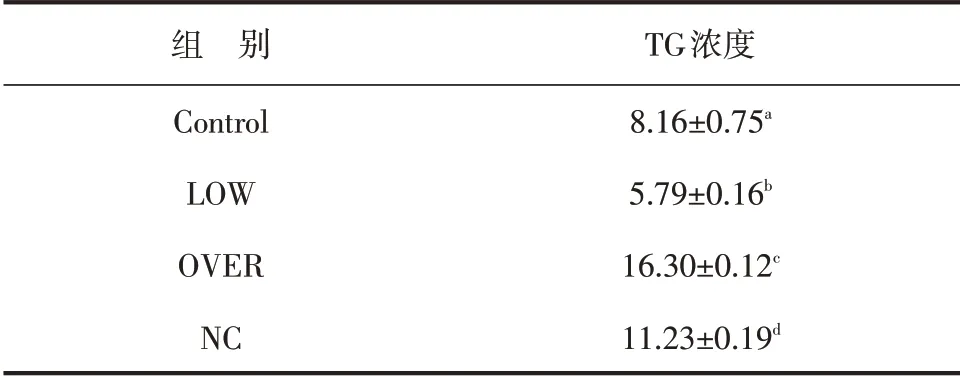
表2 各组VLDL浓度
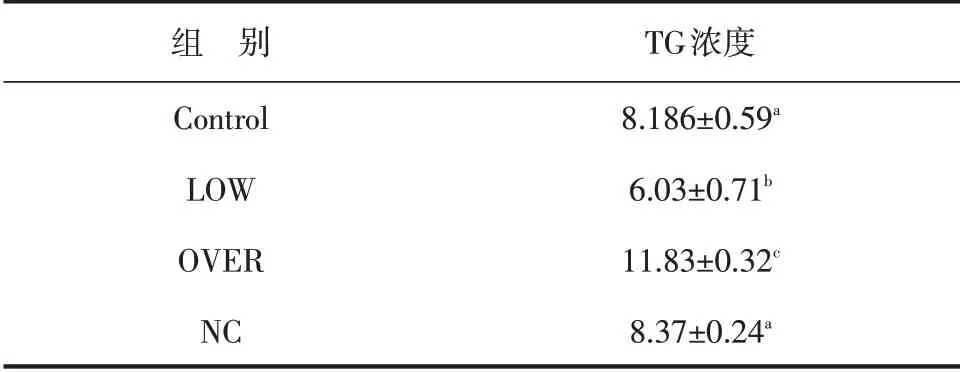
表3 细胞中甘油三酯浓度
3 讨 论
脂肪肝是围产期奶牛常见多发的营养代谢性疾病,是能量负平衡而导致的肝脏脂质沉积,对奶牛养殖业的危害较大[14]。能量代谢主要在肝脏进行,肝脏参与脂质代谢的多个重要环节,包括脂肪酸的摄取与合成,脂质的加工、贮存、氧化分解及输出等[15]。FMO有多个基因型,基因产物多分布于肝组织的内质网中[16]。人肝脏中含有3 种FMO,在成人肝脏至少有两类FMO,其中最主要的是FMO3[17]。FMO3 是近年来发现的与脂代谢相关的调节因子[18]。牛的代谢特征与人和鼠的不同,且奶牛脂肪肝的发病率高于单胃动物,FMO3与牛的脂代谢是否有关系至今尚未见研究。
VLDL 是肝细胞内TG 运出外周组织的唯一因子。VLDL 主要在肝脏合成,其功能是将肝内TG以VLDL 的形式从肝脏转移至外周储存利用[19]。当肝脏内脂肪酸显著升高时,肝脏VLDL输出总量增加,但并不能减弱脂肪沉积。ApoB100、ApoE 是VLDL 的重要组成部分,ApoB100 是VLDL 中的TG结合部分,ApoE 是VLDL 由新生到成熟的重要蛋白[9]。有研究表明,过表达ApoE 能恢复小鼠原代肝细胞中VLDL-TG 的代谢,Patatin 样磷脂酶结构域蛋白7(Patatin-like phospholipase domain contain⁃ing protein 7,PNPLA7)能通过与ApoE的相互作用调节ApoE 的稳定性从而调节肝脏VLDL 的分泌。MTTP 是VLDL 合成过程中重要的调节蛋白,主要功能是将肝细胞中游离的TG 转移到VLDL 上,使TG 随VLDL 的分泌排出[20]。广藿香醇可通过增加ApoB100 的分泌和MTTP 的活性来恢复VLDL 的合成和输出[21-22]。
研究表明,在高脂血症小鼠体内过表达FMO3会增加肝脏和血浆中的脂质[12]。本试验中,FMO3低表达时ApoB100 和ApoE基因的mRNA 表达量显著低于对照组,表明FMO3低表达会减弱ApoB100和ApoE基因的mRNA 表达水平,过表达FMO3 时ApoB100 和ApoE的mRNA 表达量则显著升高。MTTP 在FMO3 低表达时与空白对照没有显著性差异,但是过表达FMO3 时则显著升高。而VLDL和TG 在低表达FMO3 组显著降低,过表达时显著升高。这些结果说明了FMO3低表达时,通过抑制ApoB100和ApoE的表达,降低了VLDL的组装与分泌以及TG 的合成;FMO3 过表达时,通过增加ApoB100、ApoE基因和MTTP的表达增加了VLDL的组装与分泌及TG 的合成。研究表明,FMO3 过度表达时能增加肝细胞中TG 的沉积,在体内可引发脂肪肝;而FMO3 低表达可降低肝细胞TG 的积聚,从而降低体内TG 的含量,减少脂肪肝的发生,本研究为奶牛脂肪肝的发生机理和治疗奠定理论基础。
