石漠化治理区不同优势树种根际土壤酶活性与土壤理化性质和微生物数量的关系1)
2021-01-17郑武扬王艳霞王月江冯刚刚
郑武扬 王艳霞 王月江 冯刚刚
(西南林业大学,昆明,650224)
根际是指植物根系周围、受根系影响的数毫米内的区域[1],是研究植物、土壤、微生物之间相互联系的重要生态领域[2]。许多学者在土壤修复[3]、植被恢复[4]、农作物耕作[5-6]相关领域进行了植物根际研究。根际是土壤和植物进行物质交换的重要区域,微生物和酶活性在根际土壤中十分活跃。根际微生物作为重要的物质转化调节器,其生物活性、种类特征能反映土壤的肥力和植物的营养状况[7]。土壤中的一切生物化学反应,实际上都是在酶的参与下进行的[8],土壤酶活性可以反映土壤质量状况。在土壤中,酶活性与土壤理化性质和微生物数量联系紧密,许多学者热衷于研究他们之间的关系[9-11]。简单的相关系数无法完全表明一个变量对另一个变量的直接作用,而通径分析能够对各变量之间的相互关系进行详细分析,将相关分析和通径分析相结合能够更详细地表达两个变量之间的具体关系。我国西南喀斯特地区是世界上面积最大的喀斯特地区,而石漠化是喀斯特地区土地退化的极端形式[12]。石漠化地区土壤性质的研究一直是一个热点[13-15],通过人工造林提高植被覆盖率和土壤质量能够有效改善石漠化[16-17]。在建水县小关村石漠化治理区,人工造林是其主要治理措施。优势植物对土壤酶活性、理化性质、微生物数量具有较大影响。本研究通过对区域内不同优势树种的根际土壤酶活性和理化性质、微生物数量进行研究,进而探讨比较不同优势树种根际土壤环境的差异以及不同指标间的内在联系,能够为全面评价该治理区土壤质量以及植被恢复提供依据。
1 研究区概况
研究区位于云南省建水县小关村喀斯特石漠化综合治理区(23°23′~23°44′N,102°44′~102°48′E),该治理区样地类型为云南松(Pinusyunnanensis)-油杉(Keteleeriafortunei)混交林和马尾松(Pinusmassoniana)-干香柏(Cupressusduclouxiana)混交林,郁闭度为25%,树龄20 a。海拔1 480~1 780 m,坡度较陡,地势南高北低,属亚热带季风气候区,夏季高温多雨,冬季温和少雨。年均降水量为784 mm,年均气温为19.8 ℃,年均日照时间2 322 h。受成土母质的影响,治理区主要土壤类型为红壤和石灰土,样地其他情况见表1。
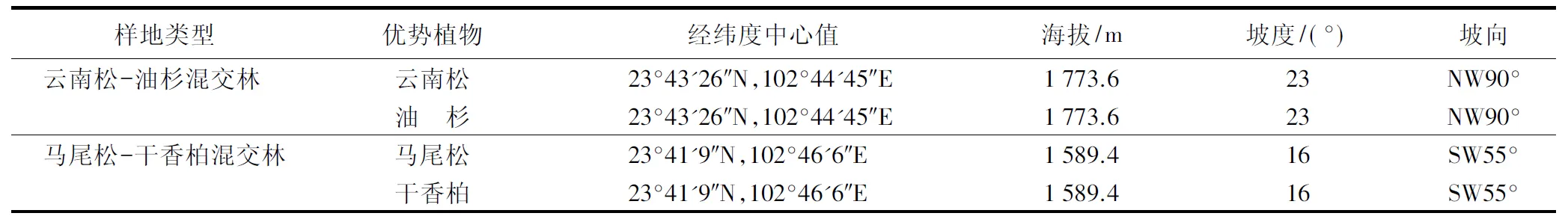
表1 喀斯特石漠化综合治理区样地基本情况
2 研究方法
2.1 采样方法
2018年10月下旬,在治理区内分别采集云南松-油杉混交林中的油杉、云南松,马尾松-干香柏混交林中的马尾松、干香柏4种优势树种的根际土壤。在2个林区各选择1块30 m×30 m的调查样地。采用抖落法[18]采集根际土壤。在每块样地内按“S”型采样法取土,每种优势植物设置3个样点,共采集12个土样,在0~30 cm深度土层中挖取具有完整根系的土体,先轻轻抖落大块不含根系的土壤,再用软刷将根表面附着的土壤(根表面上3 mm以内)刷下,分别装入自封袋中得到根际土壤。采集的土壤样品一部分4 ℃保存,用作土壤微生物试验;一部分经风干、研磨后供土壤酶活性和理化性质测定。
2.2 测定方法
土壤微生物数量的测定采用稀释平板法,细菌使用牛肉膏蛋白胨培养基;真菌使用孟加拉红培养基;放线菌使用改良高氏一号培养基。过氧化氢酶活性采用高锰酸钾滴定法测定,结果以1 g风干土壤滴定所需0.002 mol·L-1KMnO4的体积(以mL计)表示;脲酶采用苯酚钠比色法测定,酶活性测定结果以3 h后100 g土产生的铵态氮的质量(以mg计)表示;蔗糖酶采用滴定法测定,酶活性测定结果以单位土质量的0.1 mol·L-1硫代硫酸钠的体积(以mL计)表示[19-20]。土壤有机质采用重铬酸钾容量法-外加热法测定[21];土壤pH采用电位法测定;土壤全氮采用高氯酸-硫酸消化-蒸馏定氮法测定;土壤碱解氮采用碱解扩散吸收法测定;土壤全磷采用高氯酸-硫酸消化-钼锑抗比色法测定;土壤速效磷采用碳酸氢纳浸提-钼锑抗比色法测定;土壤全钾采用氢氧化钠熔融-火焰光度法测定;土壤速效钾采用乙酸铵浸提-火焰光度法[22]测定。
2.3 数据分析
采用Excel 2010对数据进行整理,SPSS 23.0对试验数据进行LSD比较、相关性分析和通径分析。
3 结果与分析
3.1 不同优势树种根际土壤pH和养分差异
由表2可见,4个树种根际土壤的理化性质各指标中除速效磷质量分数外均存在显著差异(P<0.01)。马尾松和干香柏根际土壤呈中性,云南松和油杉根际土壤呈酸性。马尾松和干香柏根际土壤有机质、全氮、碱解氮、全磷、全钾、速效钾质量分数大于云南松和油杉,而速效磷质量分数则小于云南松和油杉,说明马尾松-干香柏混交林根际土壤理化性质指标总体高于云南松油杉混交林。

表2 喀斯特石漠化综合治理区不同树种根际土壤理化性质
3.2 不同优势树种根际土壤微生物数量差异
由表3可知,4个树种根际土壤微生物数量中除真菌数量外均存在显著性差异(P<0.01),其中,油杉根际土壤微生物数量最多。4个树种均以细菌数量最多,其次是放线菌和真菌。云南松和油杉根际土壤微生物数量显著高于马尾松和干香柏,说明云南松-油杉混交林根际土壤微生物数量高于马尾松-干香柏混交林。

表3 喀斯特石漠化综合治理区不同树种根际土壤微生物量
3.3 不同优势树种根际土壤酶活性差异
由表4可知,4种优势树种根际土壤酶活性均存在显著性差异(P<0.05)。干香柏和马尾松的过氧化氢酶、脲酶活性显著高于油杉,而蔗糖酶活性则显著低于云南松,这说明马尾松-干香柏混交林过氧化氢酶活性和脲酶活性高于云南松-油杉混交林,蔗糖酶活性则低于云南松-油杉混交林。
3.4 根际土壤酶活性与土壤养分、微生物数量的相关性
由表5可知,石漠化治理区4种优势树种根际土壤过氧化氢酶活性与土壤pH、全氮、全磷、全钾呈极显著正相关关系,与细菌数量、放线菌数量呈极显著负相关关系;土壤脲酶活性与土壤pH、全钾呈极显著正相关关系,与土壤全氮呈显著正相关关系,与土壤细菌数量、放线菌数量呈极显著负相关关系;土壤蔗糖酶活性与土壤全氮、碱解氮、速效钾呈显著负相关关系。土壤酶活性与其他土壤理化性质指标和微生物数量相关性较低。其中土壤速效磷和真菌数量与3种酶活性均无显著相关关系。

表4 喀斯特石漠化综合治理区不同树种根际土壤酶活性
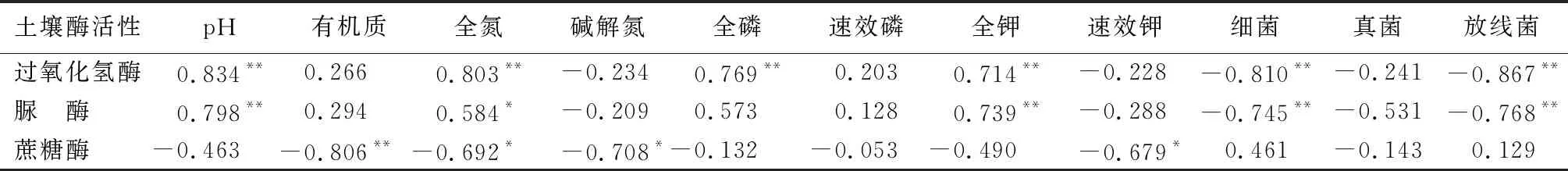
表5 喀斯特石漠化综合治理区4种优势树种根际土壤酶活性与理化性质和微生物相关系数
3.5 根际土壤酶活性与土壤养分、微生物数量的通径分析
对土壤酶活性与土壤养分、微生物数量进行多元回归分析,得到标准化回归方程:
Y1=-0.184X1-0.179X2+0.411X3-0.139X4+0.015X5+
0.481X6-0.241X7-0.208X8+0.324X9+0.085X10-0.597X11。
Y2=0.798X1-0.246X2-0.160X3-0.200X4+0.060X5+
0.269X6-0.188X7-0.147X8+0.208X9-0.319X10-0.213X11。
Y3=-0.079X1-0.806X2-0.458X3-0.293X4-0.258X5-
0.296X6+0.093X7-0.600X8+0.106X9-0.296X10+0.044X11。
式中:Y1为过氧化氢酶活性;Y2为脲酶活性;Y3为蔗糖酶活性;X1为pH;X2为有机质质量分数;X3为全氮质量分数;X4为碱解氮质量分数;X5为全磷质量分数;X6为速效磷质量分数;X7为全钾质量分数;X8为速效钾质量分数;X9为细菌数量;X10为真菌数量;X11为放线菌数量。
由表6可知,土壤理化性质和微生物数量对土壤过氧化氢酶活性直接影响系数大小顺序为土壤全钾、放线菌数量、pH、碱解氮、速效钾、全氮、有机质、细菌数量、全磷。根据各土壤因子对过氧化氢酶活性的通径系数可知,土壤全钾、放线菌数量、pH通过自身的直接作用以及与其他因子的间接作用在很大程度上影响过氧化氢酶活性,土壤全钾、放线菌数量、pH之间的间接作用也影响着过氧化氢酶活性。其他因子的影响系数较小,因此,土壤全钾、放线菌数量、pH是影响过氧化氢酶活性的重要因子。
土壤理化性质和微生物数量对脲酶活性直接影响系数由大到小依次为土壤pH、放线菌数量、全氮、碱解氮、全钾、细菌数量、全磷、速效钾、有机质。根据各土壤因子对脲酶活性的通径系数可知,土壤pH、放线菌数量、全氮通过自身的直接作用以及与其他因子的间接作用在很大程度上影响着脲酶活性,土壤pH、放线菌数量、全氮之间的间接作用对脲酶活性有很大影响,此外,土壤全钾、细菌数量与pH之间的间接关系对脲酶活性也有较大影响,而其他因子对脲酶活性影响较小,因此,土壤pH、放线菌数量、全氮是影响脲酶活性的重要因子。
土壤理化性质和微生物数量对蔗糖酶活性直接影响系数由大到小依次为土壤pH、全钾、全氮、速效钾、放线菌数量、细菌数量、有机质、碱解氮、全磷。根据各土壤因子对蔗糖酶活性的通径系数可知,土壤pH、全钾、全氮通过自身的直接作用以及与其他因子的间接作用在很大程度上影响蔗糖酶活性,土壤pH、全钾、全氮之间的间接作用对脲酶活性有很大影响,其他因子对蔗糖酶活性的影响系数较小,说明土壤pH、全钾、全氮是影响蔗糖酶活性的重要因子。

表6 喀斯特石漠化综合治理区4种优势树种土壤酶活性与土壤理化性质、微生物数量间通径系数
4 结论与讨论
在4种优势树种中马尾松和干香柏根际土壤呈中性,云南松和油杉根际土壤呈酸性。与云南松-油杉混交林相比,马尾松-干香柏混交林根际土壤有机质、全氮、碱解氮、全磷、全钾、速效钾质量分数更高。微生物在根际土壤中十分活跃,其活动和代谢作用促进或抑制根系分泌物的释放[23]。4个树种根际土壤微生物数量均是细菌数量最多,其次为放线菌和真菌,这与许多学者[23-25]的研究结果一致。云南松-油杉混交林根际土壤微生物数量显著高于马尾松-干香柏混交林。马尾松和干香柏根际土壤蔗糖酶活性低于云南松和油杉,但过氧化氢酶和脲酶活性高于云南松和油杉。马尾松和干香柏根际土壤全氮质量分数高于云南松和油杉,主要由于马尾松和干香柏根际土壤脲酶活性较高,因为脲酶主要负责催化水解有机物生成氨和CO2,是氮素的主要来源[26]。
针叶林土壤酶活性与理化性质、微生物数量相关性显著,这与舒树淼等[27]在基于结构方程的滇东石漠化地区土壤理化性质与酶活性研究、孙英杰等[28]在广西十万大山南麓不同植被类型土壤酶活性、鱼海霞等[29]在玉龙雪山土壤酶活性特征及其与土壤养分的关系研究结果一致。微生物分泌物是土壤酶的主要来源之一,这与范周周[30]在岩溶与非岩溶区不同林分根际土壤微生物及酶活性研究结果一致。土壤酶主要来源于植物根系分泌物、微生物分泌物、土壤动物排泄物,不同的植被类型、管理方式、土壤环境都会对土壤酶活性产生影响,进而影响到治理区生态系统,因此要对不同种植模式、不同优势植物土壤的酶活性、理化性质和微生物进行更深入地研究。
土壤全钾、放线菌数量、pH的直接作用是影响过氧化氢酶活性的重要因素。过氧化氢酶能够分解土壤中的过氧化氢,土壤微生物数量的多少与土壤理化性质都会影响这个过程。放线菌在土壤中对物质的转化会起到一定作用[26],其分泌物是过氧化氢酶的重要来源。而土壤全钾作为植物根系的重要营养物质,会影响其分泌物释放,进而影响过氧化氢酶的产生。pH会影响土壤酶促反应的酸碱环境,影响其反应速率。
土壤脲酶活性主要受pH、全钾、全氮的直接作用以及他们之间的间接作用影响。土壤pH通过自身的直接作用以及与全氮、全钾、微生物数量之间的间接作用对脲酶活性产生了极大的影响。脲酶作为一种蛋白质,酸碱度的改变会影响其活性。脲酶可以分解尿素,而尿素是主要由氮组成的有机化合物,因此土壤全氮质量分数对脲酶活性也有较大影响,全氮、全钾还可以影响植物根系分泌过程,而植物根系分泌物和微生物分泌物是土壤酶的主要来源,也可以促进酶促反应。
蔗糖酶活性主要受土壤pH、全钾、全氮的直接作用,以及全钾、细菌数量与土壤pH间接作用的影响。土壤蔗糖酶能够分解转化土壤中的蔗糖,与土壤中的动植物和微生物关系密切,土壤pH、全钾、全氮是土壤动植物生长活动的重要指标和营养物质,因此,它们必然会影响蔗糖酶的活性。土壤细菌分泌物是蔗糖酶的重要来源,土壤酸碱度对细菌生存环境具有重要影响,所以,细菌数量与土壤pH的间接作用也会影响蔗糖酶活性。
建水县小关村石漠化治理区不同树种根际土壤酶活性与理化性质关系和微生物数量之间相关关系显著。土壤全钾、放线菌数量、pH是影响过氧化氢酶活性的重要因子;土壤pH、放线菌数量、全氮是影响脲酶活性的重要因子;土壤pH、全钾、全氮是影响蔗糖酶活性的重要因子。总体来看,pH对3种土壤酶活性都具有重要影响。在石漠化治理区,土壤酶活性受土壤理化性质影响较大,受微生物数量的影响相对较小,且土壤养分含量较少,则凸显了土壤pH对土壤酶活性的影响。
