荒漠河岸胡杨林生态系统能量分配及蒸散发
2021-01-16马小红
马小红,冯 起
1 太原师范学院汾河流域科学发展研究中心, 晋中 030619
2 中国科学院西北生态环境资源研究院内陆河流域生态水文重点实验室, 兰州 730000
全球干旱地区覆盖了地球表面的41%,养育着全球约三分之一的人口,包括极端干旱、干旱、半干旱和半湿润地区[1],水过程是这些地区最为关键的生态过程[2]。深入了解旱地生态系统的水热交换过程是有效地、可持续地进行水资源调控和管理的前提[3]。与周围的荒漠环境相比,内陆河岸生态系统的生物多样性最为丰富、生产力最高、动态变化最显著[4],是维护地区生态安全、保障地区生态环境的重要屏障。黑河是我国的第二大内陆河,在20世纪90年代末期,水资源的过度开发和水管理政策的缺失使得黑河下游曾面临着严重的生态环境问题[5],如尾闾湖消失、绿洲面积萎缩、沙漠化严重等等。因此,为了恢复黑河下游地区恶化的生态环境,黑河流域管理局依据2001年国务院批复的《黑河流域近期治理规划》开始实施生态输水工程[6]。生态输水工程对黑河下游地区胡杨的水热交换过程产生了重要的影响,但这一部分的研究还远远不够。因此,有必要对其进行深入研究,从而为流域水资源的合理开发、利用和管理提供一定的帮助。
在干旱地区,蒸散发一般代表了整个生态系统的水汽耗散,将生态系统的水文状态和生物过程紧密地联系在一起[2]。而生态系统中潜热和感热分配决定着水循环过程、边界层厚度和结构、以及环境因子的变化[7]。涡度相关技术是直接测定下垫面与大气间水汽、二氧化碳及能量通量交换的微气象技术[8]。利用涡度相关技术,从生态系统尺度进行长期、连续的定点观测,有助于更加深入地了解旱地生态系统蒸散发和能量通量的变异性。目前,全球大约建有240多个通量观测塔[9- 14],其中绝大多数用于观测北方森林、温带森林和热带森林生态系统与大气之间的水热交换过程,而用于观测干旱地区森林生态系统与大气之间水热交换过程的观测塔较少。Cleverly等研究了有无洪水淹没条件下,美国南部格兰德河中游河岸林(柽柳)的蒸散量[15];Yuan等通过研究塔里木河下游荒漠河岸林(胡杨和柽柳)的蒸散规律及关键控制过程得出:地下水是地表蒸散的主要水分来源,是控制荒漠河岸林水循环的关键因素[16];Yu等通过研究黑河下游荒漠河岸林的蒸散发得出:当地下水埋深小于3 m,蒸散发与地下水埋深之间呈线性相关关系[17]。以上研究虽然很好地分析了荒漠河岸林蒸散发的变化规律,但是有关荒漠河岸林的水热交换过程及影响机理的研究仍然比较欠缺,有必要进一步深入研究。本文基于水文、气象和涡度通量观测数据进行以下研究:(1)分析能量平衡各分量的季节变化趋势及年际差异;(2)确定胡杨林的能量分配特征,量化潜热和感热通量占有效能的比例;(3)确定胡杨林的蒸散量;(4)分析生态输水过程对能量分配和蒸散发的影响。
1 研究区概况
研究区位于黑河下游的额济纳绿洲,天然绿洲面积约为3428 km2,占额济纳旗总面积的5.7%[18]。通过分析1957—2016年额济纳旗气象观测站的数据可知:该区域气候极端干旱,平均年降水量为37.5 mm,多年平均潜在蒸发量为2240.5 mm,干旱指数小于0.05;多年平均空气温度为8.9 ℃,其中1月和7月的多年平均空气温度为- 11.5 ℃和27.0 ℃;年降水量的季节变化趋势和年际波动明显,约75%的降水集中在6—10月,1957—2016年的年降水量在7—103 mm之间波动。
涡度通量观测仪器位于七道桥胡杨林国家自然保护区内,地理坐标为101°10′ E,41°59′ N,海拔高度为921 m(图1)[17]。该区域的植被群落结构简单,基本由单一的胡杨组成,胡杨占地表的75%,植株密度为350株/hm2,树高为(10.1±1.7) m,胸径为(22.9±4.8) cm,树龄约30—60 a。林下灌木和草本极为稀疏,灌木树种为柽柳,草本层主要为苦豆子。额济纳地区的地带性土壤为灰棕漠土和石膏棕漠土,而在不同的植被类型下土壤的性质和结构差异很大,从而形成了不同的非地带性土壤。该研究样地的土壤,以沙壤土和沙土为主,土壤结构变化不明显,自地表往下依次为沙壤土(0—60 cm),沙土(60—120 cm),沙土和黏土混合层(120—160 cm)。该区域的降水极为稀少,胡杨林的生长主要依赖于地下水,而地下水埋深的高低直接受生态输水过程的影响。生态输水过程包括春季和秋季的漫水灌溉。
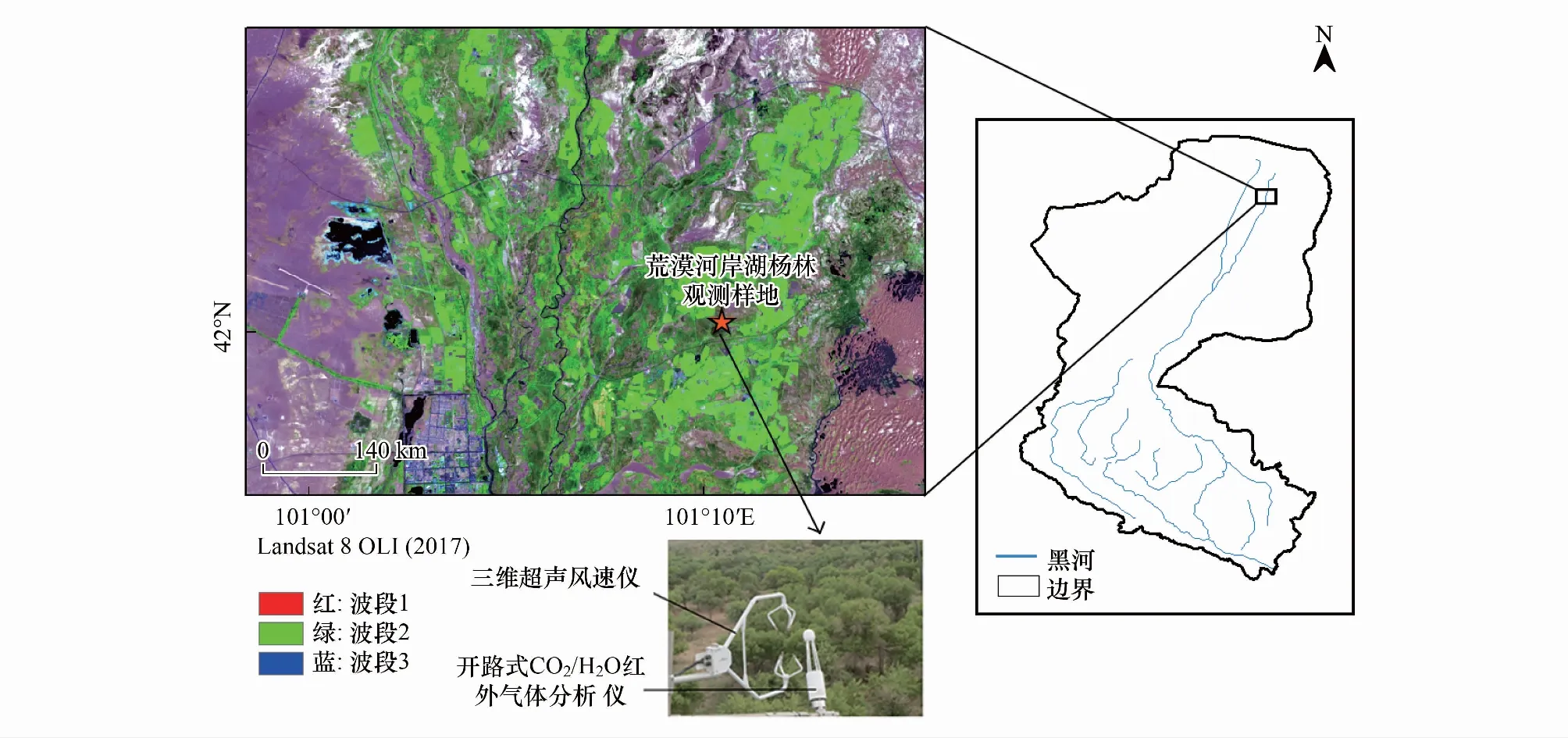
图1 研究区的地理位置(胡杨林观测样地)Fig.1 The location of the study (Populus Euphratica Forest Stand)
2 数据来源及方法
2.1 涡度通量及环境因子监测
涡度相关系统所在区域的下垫面地势平坦、地表植被均一性好、受扰动小,胡杨林在各个方向上可延伸几千米。地表粗糙度和零平面位移分别为1.38 m和7.9 m,地表坡度为1/5000。主要的通量贡献源区为盛行风(西北东南风)方向上,距离观测塔109 m至1520 m的区域;其中距离观测塔228 m处的通量贡献最大。
涡度系统包括用于测量风速的三维超声风速仪(CSAT3, Campbell Scientific, Inc., Logan, UT, USA)和用于测量水汽和CO2浓度的开路式CO2/H2O红外气体分析仪(LI- 7500, LI-COR Inc. Lincoln, NE, USA),安装高度为20 m。每年的5月或6月,对红外气体分析仪进行校准。净辐射(Rn,W/m2)、空气温度(Ta,℃)、相对湿度(RH,%)和涡度通量交换数据,均由CR3000数据采集器(Campbell Scientific, Logan, UT, USA)采集。其中Rn由四分量净辐射仪(CNR4, Kipp & Zonen, Delft, NL)测得,Ta和RH由相对温湿度传感器(HMP45C, Campbell, USA)直接测得。水汽压亏缺(VPD,hpa)是根据温度和相对湿度计算而来的。土壤热通量(G,W/m2)由两个被埋藏于地下5 cm处、间隔0.5 m的土壤热通量板(HFP,Hukse Flux Thermal Sensors B.V., Delft, The Netherlands)测得。此外,在距离涡度通量塔30 m处,有一个约6 m深的地下水埋深观测井,里面安装有一个自记式压力传感器(HOBO-U20)用于测量并计算地下水埋深(GWD, m),测量频率为半小时。土壤温度、湿度和盐分传感器(SMEC300)安装在距离地面10 cm,30 cm,50 cm和80 cm土层深处,其中每个土层安装6个传感器,通过测量土壤温度(Ts,℃)、土壤含水量(θ,%)和电导率(EC, mS/cm),来反映观测样地的土壤温湿度和盐分状况。
2.2 通量数据处理及缺失数据插补
涡度通量数据处理的基本过程包括:(1)利用Loggernet软件对原始10Hz的通量数据进行格式转换及数据分割;(2)用Eddypro软件进行基本的数据处理,包括异常值剔除、倾斜校正、时间滞后校正、频率响应校正、超声虚温校正以及密度效应校正(WPL)校正;(3)剔除夜间湍流发展不充分时数据(摩擦风速小于0.16 m/s)。对于缺失的通量数据,若缺失数据不超过2小时则用线性内插方法插值,否则用查表法根据空气温度和光照等主要环境因子对缺失数据进行插补[8,19]。在此基础上,再利用平均日变化法(移动窗口为15 d)进行插值,得到完整时间序列的能量及水汽通量数据。
2.3 胡杨林生态系统的能量闭合方程
根据热力学第一定律,涡度通量观测中的能量平衡闭合是指涡度系统所观测的潜热通量(LE)与感热通量(H)的和应该等于有效能[20]:
LE+H=Rn-G-S-Q
(1)
式中,Rn是净辐射,G是土壤热通量,S是冠层内储存热量的变化量,Q是所有额外能量的源和汇。在实际应用过程中,Q由于太小通常被忽略掉了。由于荒漠河岸林的树木分布比较稀疏且冠层盖度较低,S所占净辐射的比例很小,在能量平衡分析过程中可以被忽略[21]。本研究发现S占净辐射的比例不足0.4%。因此,在能量平衡分析过程中忽略了S项,荒漠河岸胡杨林生态系统的能量平衡方程简化为:
LE+H=Rn-G
(2)
式中,Rn是净辐射,G是土壤热通量,LE是潜热通量,H是感热通量。潜热通量是近地层大气与下垫面之间水分的热交换,主要包括植物蒸腾和土壤蒸发所消耗的热量;感热通量是温度变化所引起的下垫面与大气之间的热交换,即温度升高或降低所需要吸收或者释放的热量。
能量闭合度不仅能在一定程度上反映胡杨林生态系统的水热交换过程,还是反映涡度通量数据可靠性的重要指标[22]。通常有两种方法用于评价生态系统的能量闭合程度:一种是线性回归方法,对湍流能量通量(LE+H)和有效能(Rn-G)进行线性拟合,线性拟合的斜率用来说明能量闭合的程度;另一种是通过计算能量闭合率(EBR)来说明能量闭合情况。本文利用线性回归方法分析了荒漠河岸胡杨林生态系统的能量闭合程度。
2.4 胡杨林蒸散发的计算
涡度相关技术能够直接测得生态系统尺度的潜热通量,若要得到半小时生态系统蒸散发(ET,mm/30min)数据,则需要根据下列公式进行计算:
(3)
式中,LE是潜热通量(MJ m-230min-1),L为水的汽化潜热(2.45 KJ/g),ρw为水密度(1 g/cm3)。
2.5 波文比的计算
波文比(β)为某一界面上感热通量和潜热通量的比值,即:
(4)
式中,H是感热通量,LE是潜热通量。
3 结果与分析
3.1 环境因子的季节及年际变化特征
黑河流域的生态输水工程直接影响着荒漠河岸胡杨林生态系统的水文状况。由图2可以看出,地下水埋深(GWD)和土壤含水量(θ)的变化直接受灌水事件的影响。灌水后,GWD的值迅速减小,甚至地表会出现积水(图中GWD的值在0 cm以上表明地表有积水);随后GWD的值逐渐增加,在下一次灌水时,GWD的值又迅速减小。灌水的量直接决定了GWD的整体水平,2014—2016年GWD的平均值分别为1.28 m、1.79 m和1.16 m。通过对比2015年和2016年秋季灌水前后的地下水埋深来看,2016年的灌水量更大,相应地30 cm 和50 cm深处的土壤含水量也更大。2015年秋季灌水前后的地下水埋深分别为2.27 m和1.14 m,而2016年灌水前后的地下水埋深分别为2.27 m和0.00 m。土壤含水量的季节变化趋势明显,秋季灌水后θ值迅速增大,随后在蒸发和入渗作用下逐渐降低;春季随着Ta和Ts的回升,θ值逐渐增加,春灌后θ值迅速增大。
空气温度(Ta)的季节变化趋势明显(图2),自3月开始快速增加,5月中旬增加趋势变缓,7月至8月增加到最大且波动明显,9月之后快速降低。土壤温度(Ts)的季节变化趋势和空气温度的基本一致,只是变化幅度不如空气温度的剧烈。一般而言,10月至次年2月,空气温度低于土壤温度;3—9月,空气温度高于土壤温度。
水汽压亏缺的季节变化趋势明显,年际之间无明显差异(图2)。11月至2月,水汽压亏缺的值很低且基本没有变化;自3月开始水汽压亏缺的值快速增加;5—9月水汽压亏缺的值较高且波动明显;9月之后水汽压亏缺的值快速降低。2014—2016年生长季(5—10月)水汽压亏缺的平均值分别为19 hpa、22 hpa和21 hpa,水汽压亏缺的年际差异不明显。
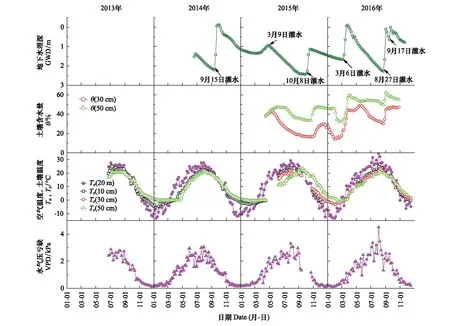
图2 地下水埋深(GWD)、土壤含水量(θ)、空气温度(Ta)、土壤温度(Ts)及水汽压亏缺(VPD)的季节和年际变化特征Fig.2 The seasonal and yearly variations of groundwater depth (GWD), soil moisture content (θ), air temperature (Ta) and vapor pressure deficit (VPD)
3.2 能量分配特征
3.2.1能量闭合程度
本文通过线性回归方法对荒漠河岸胡杨林生态系统的湍流能量通量(LE+H)和有效能(Rn-G)进行线性拟合,如图3所示。2013—2016年湍流能量通量和有效能之间的拟合斜率分别为0.63,0.79,0.90和0.81,平均为0.78。已有研究表明湍流能量通量和有效能之间的线性回归斜率在0.55至0.99之间波动[16,20,23,24],因此本研究结果是合理的、可接受的。
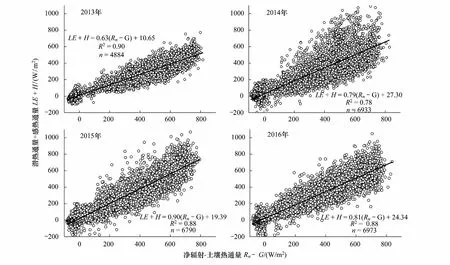
图3 2013—2016年半小时尺度上胡杨林生态系统的能量闭合程度Fig.3 Linear regression for the half-hourly energy balance closure for 2013—2016
3.2.2能量平衡各分量的季节及年际差异
图4给出了2013—2016年净辐射、潜热通量、感热通量和土壤热通量的季节变化趋势。(1)净辐射的季节变化趋势明显(图4)。2014—2016年,净辐射分别在5.5—23.1 MJ m-2d-1、4.7—22.0 MJ m-2d-1和3.4—23.2 MJ m-2d-1之间波动。2014—2016年,5—10月净辐射的日均值分别为18.5、19.8、20.2、19.0、17.0和12.1 MJ m-2d-1,其中6月和7月的净辐射最大。(2)潜热通量的季节变化趋势明显,年际波动较大。3月开始,土壤进入消融阶段,土壤中大量的固态水转化为液态水,土壤蒸发增大,LE随着Rn和Ta的增加而增加。同时,春灌使得土壤中可供蒸发的总水量进一步增大,LE增加到一定程度后保持不变。5月胡杨进入萌芽期,LE随着植物蒸腾的增大而逐渐增大;6—8月,LE增加到最大,且波动明显。9月,虽然植物蒸腾开始下降,但受秋季灌水的影响地表蒸发较大,LE的值仍然较高,并未随Rn和Ta的下降而降低。10月,LE随着土壤蒸发和植物蒸腾的下降而快速降低;12月至次年的2月,LE的值波动不大,小于1 MJ m-2d-1。潜热通量的季节和年际波动主要与土壤中可供蒸发的总水量和胡杨的物候阶段有关。春季和秋季的灌水事件直接影响着土壤中可供蒸发的总水量,从而间接地改变了潜热通量的季节变化过程。(3)感热通量的季节变化趋势与潜热通量的季节变化趋势大致相反。2014—2016年,1—12月感热通量的日均值分别为2.88、4.37、5.77、6.95、5.95、1.96、0.37、1.32、1.06、3.43、2.77和2.55 MJ m-2d-1。(4)相比于潜热通量和感热通量,土壤热通量的值很小。土壤热通量的季节变化趋势明显:春季,土壤热通量由负值转为正值;秋季,土壤热通量由正值转化为负值。
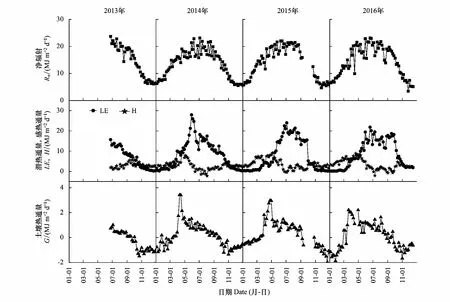
图4 净辐射(Rn)、潜热通量(LE)、感热通量(H)和土壤热通量(G)的季节变化特征及年际差异Fig.4 The seasonal and yearly variations of net radiation (Rn), latent heat flux (LE), sensible heat flux (H), and soil heat flux (G)
3.2.3能量分配特征
表1给出了2014—2016年不同月份的潜热通量、感热通量和土壤热通量占净辐射的百分比,以及不同月份的波文比。由表1可以看出,生长季,荒漠河岸胡杨林生态系统的能量消耗以潜热通量为主,潜热通量占净辐射的84.28%;其次是感热通量,感热通量占净辐射的15.87%;而土壤热通量仅占净辐射的2.1%。非生长季,能量消耗以感热通量为主(H/Rn=39.97%),其次是潜热通量(LE/Rn=18.17%)。波文比(β)的季节变化趋势明显:12月至次年2月,能量消耗以感热通量为主,β值平均为5.85;3—4月,能量消耗仍然以感热通量为主,β值平均为1.29;5月,能量消耗以潜热通量为主,β值为0.45;6—8月,能量消耗以潜热通量为主,β值平均为0.07;9—10月,受灌水的影响能量消耗仍然以潜热通量为主,β值分别为0.07和0.55;11月,能量消耗以感热通量为主,β值为1.53。

表1 2014—2016年不同月份的潜热通量(LE)、感热通量(H)和土壤热通量(G)占净辐射(Rn)的比值及波文比(β)Table 1 The monthly average of the ratio of latent heat flux (LE), sensible heat flux (H) and soil heat flux (G) to net radiation flux (Rn), and bowen ratio (β) in 2014—2016
3.3 蒸散发的季节变化特征及年际差异
蒸散发(ET)是内陆河流域水量平衡的重要组成部分,在水分短缺的干旱和半干旱地区,对蒸散发的估计在生态保护和水资源配置过程中起着非常重要的作用。荒漠河岸林的蒸散发主要受地下水埋深的影响,地下水是地表蒸散的主要水分来源[16,17]。黑河流域的生态输水工程影响着该区域的地下水埋深及土壤含水量。为了更加清楚地分析蒸散发的季节变化过程及其对生态输水的响应,这里选择气候条件较为一致、输水量差异较大的2015和2016年进行比较分析,如图5所示。
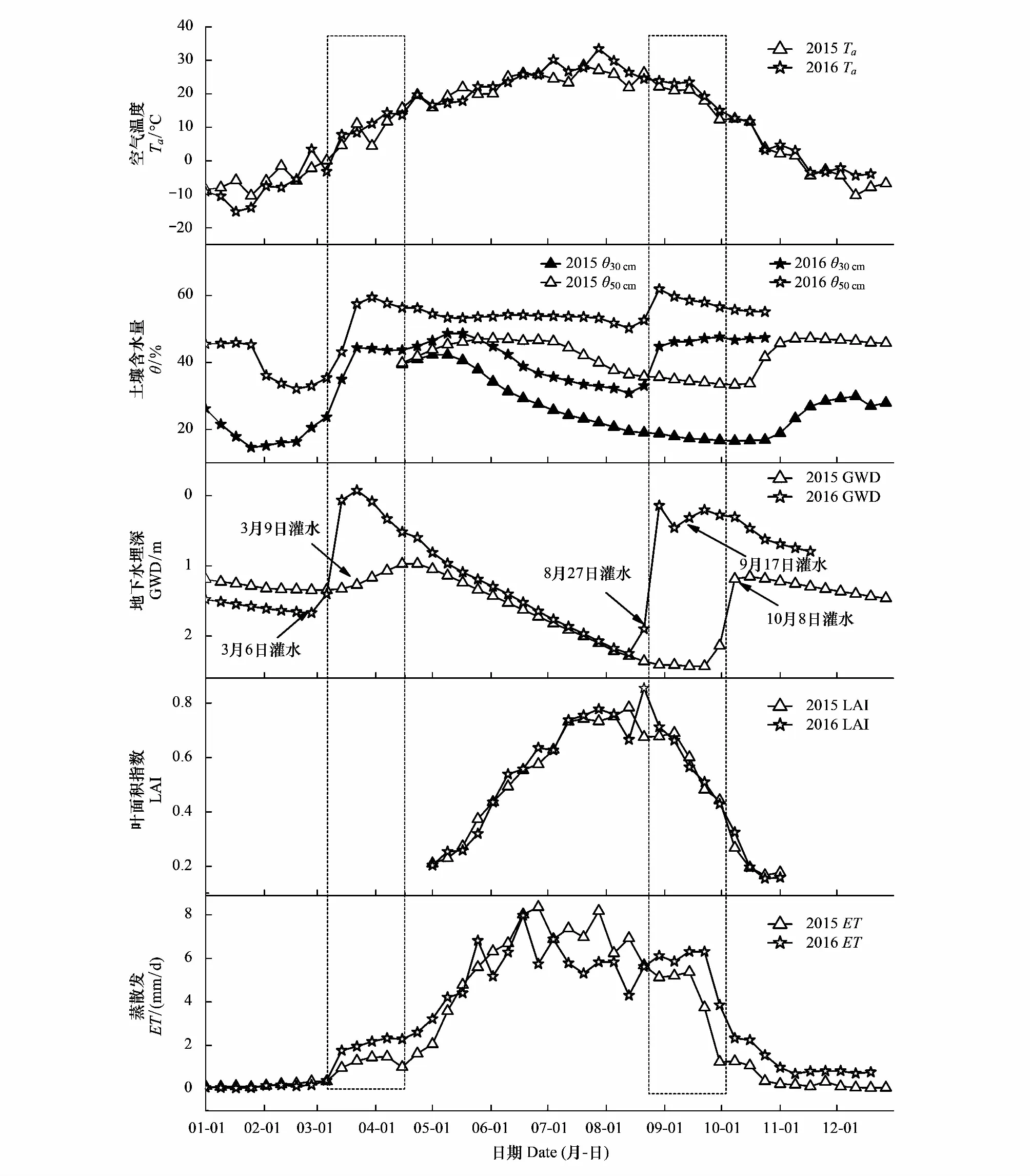
图5 2015—2016年空气温度(Ta)、土壤含水量(θ)、叶面积指数(LAI)及蒸散发(ET)的季节变化特征Fig.5 The seasonal variations of air temperature (Ta), soil moisture content (θ), leaf area index (LAI), and evapotranspiration (ET) for 2015—2016
由图5可知,荒漠河岸胡杨林蒸散发的季节变化趋势明显。11月至次年2月,ET的值很小且变化不大。3月初,ET随着θ和Ta的升高而增大。3月中旬至4月,受灌水的影响,ET维持在较高值,ET与θ的变化趋势基本一致,不再随Ta的升高而增加(红框)。自5月开始,胡杨进入生长阶段,ET随着LAI的增大而增大;6—8月ET达到最大,且波动明显。9月,受灌水的影响,ET的值较高,并未随LAI和Ta的降低而减小(红框)。春季和秋季胡杨林的蒸散发及年际差异主要受灌水的影响。灌水的时间改变了ET的季节变化趋势,使得春季和秋季的ET值较高,并未随Ta和LAI的变化而变化;灌水的量决定了春季和秋季蒸散量的年际差异,土壤含水量越大,地下水埋深越浅,蒸散量越大。2014—2016年ET的日平均值分别为3.07 mm/d、2.80 mm/d和3.11 mm/d,地下水埋深分别为1.28、1.79和1.16 m。此外,蒸散发的季节变化过程还受胡杨物候阶段的影响,2014—2016年,萌芽期(5月)、旺盛生长期(6—9月)和叶片衰老期(10月)的ET平均分别为4.03 mm/d、5.67 mm/d和1.61 mm/d。总之,荒漠河岸胡杨林蒸散量的波动主要受生态输水过程(地下水位和土壤含水量)和胡杨物候阶段(LAI)的影响,温度、辐射等气象因子的年际差异不足以引起蒸散量的较大波动。
4 讨论
4.1 能量分配特征
干旱、半干旱生态系统的能量分配通常受降水及其相关因子(浅层土壤水分和干旱事件)的控制[25- 27]。然而,黑河下游荒漠河岸林的生长主要依赖于地下水[28],其能量分配受地下水的控制[21];而生态输水工程影响着该区域的地下水埋深。本研究发现生态输水的时间和量影响着胡杨林生态系统的能量分配。
能量分配的季节变化过程受胡杨物候阶段和生态输水的显著影响。生长季胡杨林生态系统的能量消耗以潜热通量为主,萌芽期(5月)、旺盛生长期(6—9月)和叶片衰老期(10月)的波文比(β)分别为0.45、0.07和0.55。其中旺盛生长期的β值与Yuan等人对塔里木河下游柽柳灌丛的研究结论比较相近(表2),而萌芽期和叶片衰老期的β值远低于Yuan等人的研究结论[21];这主要是因为受黑河下游生态输水的影响,春季和秋季的潜热通量较大。对于极端干旱的荒漠地区而言,水分条件是决定能量分配的关键因素:若生态系统的水分充足,能量消耗则以潜热通量为主,反之则以感热通量为主(表2)[21,25,29-30]。例如Kalthoff 等对阿塔卡马沙漠南部艾尔奇山谷农耕地的研究发现:灌溉或湿季降水后,β值约等于0.7;而无植被覆盖的干季,β值约等于2.5[30]。Brümmer 等对西非热带稀树草原的研究也得出相似的结论,湿季β值小于0.7,而干季β值大于1.5[29]。荒漠生态系统能量分配的差异主要来源于水分条件、植被类型及气候特征[25]。
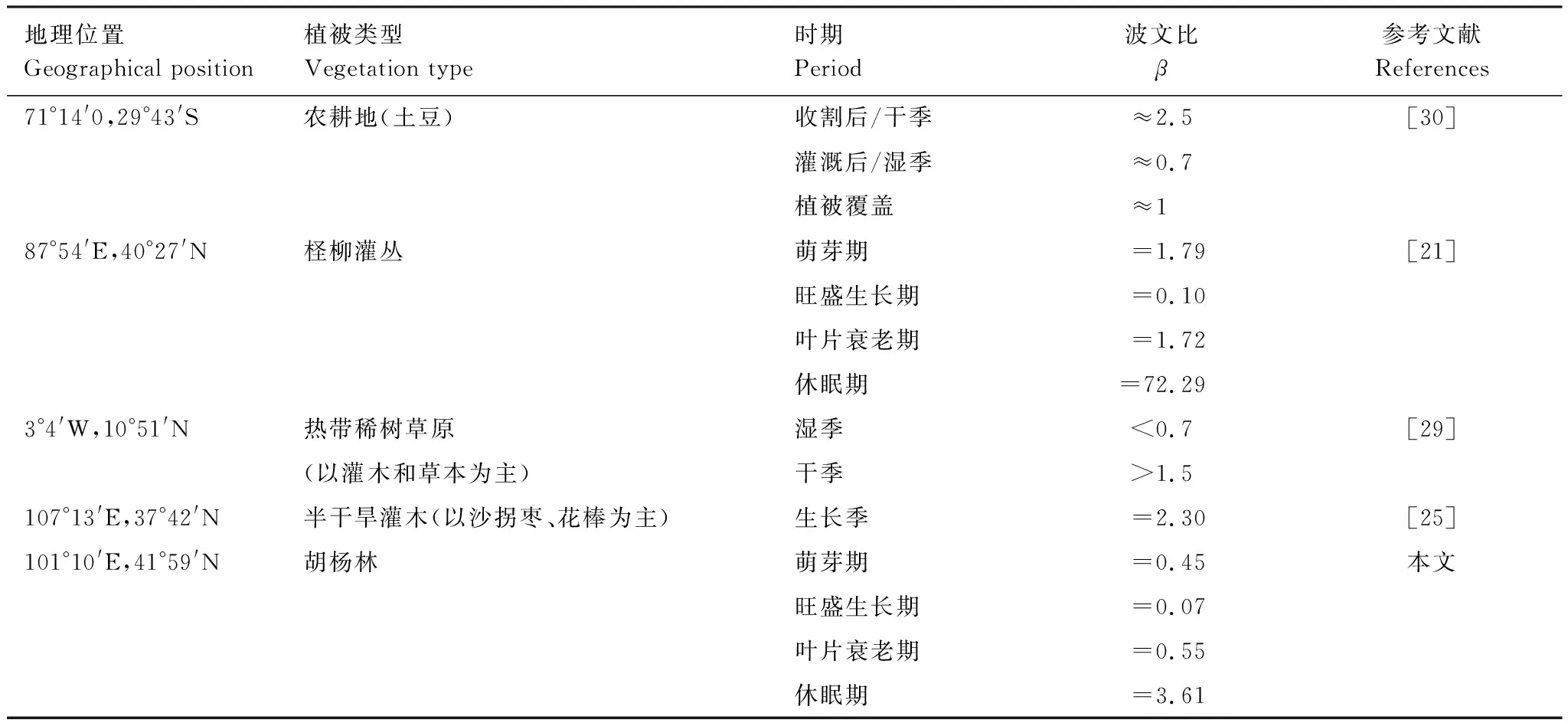
表2 旱地生态系统的能量分配特征Table 2 The characteristics of energy partitioning in dryland ecosystems
4.2 蒸散发
本研究发现春季和秋季胡杨林的蒸散发及年际差异主要受生态输水过程的影响。灌水的时间改变了ET的季节变化趋势,受灌水的影响春季和秋季的ET较高,并未随温度和叶面积指数的变化而变化;而灌水的量决定了土壤中可供蒸发的总水量,是年际之间ET出现差异的主要原因,土壤含水量越高,蒸散量越大(图5)。此外,蒸散发的季节变化过程还受胡杨物候阶段的影响。非生长季胡杨林的蒸散量很小,2015—2016年非生长季的ET分别为83 mm和193 mm;二者的差异较大,这种差异主要源于生态输水量。2015年秋季灌水前后的地下水埋深分别为2.27 m和1.14 m,而2016年秋季灌水前后的地下水埋深分别为2.27 m和0.00 m;可见2016年的灌水量高于2015年的(图2)。同理,2016年春季的灌水量也高于2015年的,这使得2016年春季的蒸散量相对较大,从而进一步增大了2015和2016年非生长季ET的差异。
2015—2016年,胡杨林的年蒸散量分别为1022 mm和1135 mm。本研究得出的ET值与Kochendorfer等人对美国加州中央山谷北部河岸林(杨树)的研究结果接近(ET=1095 mm/a)[23];比Yuan等人得出的塔里木河下游荒漠河岸胡杨林蒸散发的值大[16],这是因为植被的生长状况和生态系统的水分状况差异较大(表3)。受黑河流域生态输水工程的影响,几乎每年都会在春秋两季对下游地区的胡杨林地表进行大面积的漫水灌溉,这是该区域胡杨林蒸散量比其他区域大的主要原因。Cleverly等利用涡度相关技术对美国南部格兰德河中游河岸林的研究表明:生长季,无洪水淹没和有洪水淹没的柽柳生态系统的蒸散量分别为740.0和1220.0 mm[15]。由此可见,漫水灌溉事件对河岸林蒸散量的影响极其重要,它可能从两个方面影响蒸散量,一是通过调节河岸林的水分状况来改变植被的生理过程,二是通过增大地表蒸发来增大整个生态系统的蒸散量。
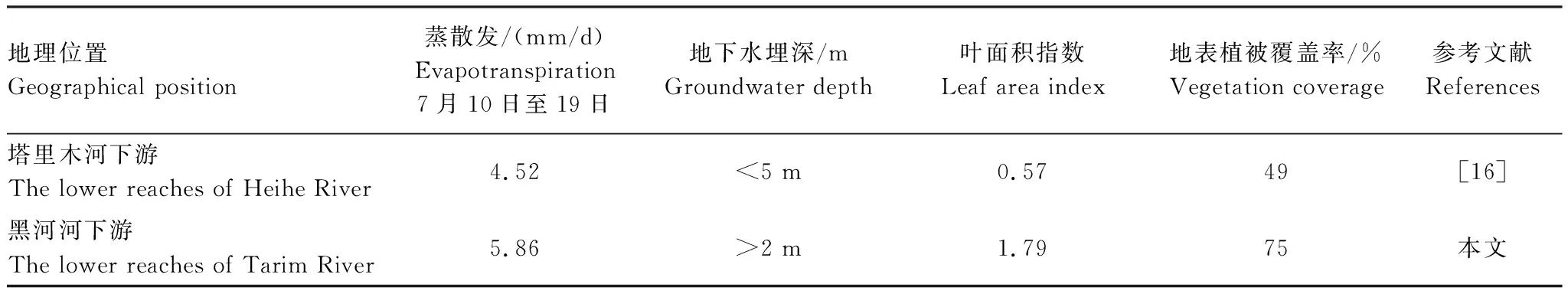
表3 中国两大内陆河流域下游地区荒漠河岸胡杨林的蒸散发Table 3 Evapotranspiration of the desert riparian Populus Euphratica forests distributed in the lower reaches of two major inland rivers in China
5 结论
本文基于涡度相关技术,研究了荒漠河岸胡杨林生态系统的能量分配及蒸散发特征,得出的主要结论有:(1)胡杨林生态系统的能量闭合率为78%。(2)生长季胡杨林生态系统的能量消耗以潜热通量为主,波文比为0.21。能量分配的季节变化过程受胡杨物候阶段和生态输水的显著影响。(3)春季和秋季胡杨林的蒸散发及年际差异主要受生态输水过程的影响。生态输水的时间改变了蒸散发的季节变化趋势,而生态输水的量决定着蒸散量的年际差异。(4)2014—2016年,生态系统的年蒸散量平均为1092 mm,生长季蒸散量平均为932 mm。
