黄土高原退耕还草地C、N、P生态化学计量特征对植物多样性的影响
2021-01-16海旭莹董凌勃汪晓珍李继伟刘玉林李妙宇潘英杰吕文文上官周平
海旭莹, 董凌勃, 汪晓珍, 邓 蕾,,*, 李继伟, 刘玉林, 李妙宇, 潘英杰, 吕文文, 上官周平,
1 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100
2 中国科学院水利部水土保持研究所, 杨凌 712100
在干旱和半干旱生态系统中,自然条件下的草地恢复被认为是控制土壤侵蚀、改善土壤质量、提高植物和生态系统健康的有效途径[1],而植物多样性恢复是植被恢复过程中的直接体现[2],因此,植被恢复过程可以通过植物功能群的组成和植物多样性的变化来反映[3]。碳(C)、氮(N)、磷(P)是生物体细胞结构组成和功能代谢中最重要的元素,也是植物生长的必需元素,其在植物生长和各种生理调节过程中起着重要的作用[4],C、N、P之间互相作用和耦合关系对维持植物良好生长和生态系统可持续发展具有重要意义[5]。生态化学计量比是近年来生态学研究的热点之一[6],C∶N∶P计量比与植物体的生长、物种多样性、群落结构与动态以及生态系统过程的联系紧密[7- 8],借助生态化学计量比特征不仅可以判断植物养分限制状况,还能揭示植物对C、N、P的利用策略[9]。因此,阐明生态系统中植物-土壤C、N、P化学计量特征对植物多样性的影响,对了解退耕还草地恢复过程中生态系统的结构和功能的变化具有重要的意义。
现有研究关于退耕还草地恢复过程中植物多样性的变化规律已有了基本一致的结论,即在恢复前期,随着恢复年限的增加植物多样性增加,在恢复后期,当群落中优势种地位突出时,植物多样性会降低[10],群落中物种多样性最大值可能出现在草地恢复的中后期[11]。植物和土壤化学计量特征是影响植物多样性的因子之一,不同群落中的植物,其生长率与植物生态化学计量特征变化有关[7, 12],且植物N∶P变化通过限制植物的生长[13],进而影响群落中植物多样性[14]。另外,杨祥祥等[15]在对沙地植物和土壤化学计量特征影响物种多样性的研究发现,在沙地植物群落中,沙柳(Artemisiaordosica)群落和油蒿(Salixpsammophila)群落的物种多样性随植物C∶N的增大而减小,而油蒿(S.psammophila)群落的物种多样性随植物N∶P的增大而增大;物种多样性随沙地土壤C∶N比和N∶P比的增加而增大[16]。然而,目前的研究仍存在一些问题:一方面,研究多集中在化学计量特征或物种多样性上,但从生态化学计量学角度对群落植物多样性方面进行探讨的研究相对较少[17];另一方面,研究多关注某一组分(植物、土壤或微生物),从植物和土壤两方面,同时关注C、N、P及其化学计量比与植物多样性的关系的研究也比较缺乏[18- 19]。
黄土高原丘陵沟壑区,生态环境十分脆弱,水土流失造成表层土壤C、N、P的大量流失[20],同时,土壤养分流失导致土地生产力大幅下降,降低了地表植被凋落物和根系养分与土壤之间的输入和周转,这两个过程直接和间接影响土壤的C、N、P化学计量特征,而土壤C、N、P化学计量特征又反过来影响群落中物种组成和植物多样性。但随着退耕还草地恢复年限的变化,导致恢复过程中植物-土壤系统养分循环及其与植物多样性的关系尚不明确,研究不同恢复年限草地植物-土壤C、N、P及其化学计量比对植物多样性的影响,有助于理解植物和土壤养分与植物多样性之间的互馈机制。本研究在黄土高原丘陵沟壑区纸坊沟流域,选取农田弃耕后自然恢复8 a、15 a、25 a和35 a的草地,分析草地恢复中植物-土壤化学计量特征和植物多样性的动态变化,并探讨植物-土壤化学计量特征对植物多样性的影响,以期为黄土高原地区草地恢复与管理提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省延安市安塞区纸坊沟流域(36° 44′—36° 51′ N, 109°15′ —109°18′ E),海拔1144—1307 m。该地区在气候划分上属于温带半干旱气候,年均温9.1 ℃,年均降水量503 mm,年日照时数2300—2400 h,无霜期160 d左右。纸坊沟属于黄土高原典型丘陵沟壑区,土壤主要为黄土母质发育形成的黄绵土,因此极易受到侵蚀而造成水土流失严重。该区的主要造林树种有刺槐(Robiniapseudoacacia)、油松(Pinustabuliformis),侧柏(Platycladusorientalis)等;灌木主要有沙棘(Hippophaerhamnoides)、胡枝子(Lespedezabicolor)、绣线菊(Spiraeasalicifolia)等;草本植物有铁杆蒿(Artemisiagmelinii)、大针茅(Stipagrandis)、长芒草(Stipabungeana)、白羊草(Bothriochloaischaemum)、阿尔泰狗娃花(Heteropappusaltaicus)、二裂委陵菜(Potentillachinensis)、茭蒿(Artemisialeucophylla)等;农田主要是玉米(Zeamays)等。
1.2 试验设计和采样
2015年8月,以农田弃耕后进行自然恢复的草地为研究对象,选取土壤类型和立地条件相似、受外界干扰程度较低的、分别恢复8 a、15 a、25 a和35 a的自然草地,以农田(玉米地)为对照,并将整个恢复年限划分为不同的时期,分别为恢复前期(恢复0—8 a)、恢复中后期(恢复15—25 a)和恢复后期(恢复35 a)。各恢复年限的退耕还草地分别设置3个立地相似的重复样地,每个样地内设置20 m×20 m大小的样方,在每个样方的中心和对角线设置3个1 m×1 m的小样方进行草本调查,调查项目包括物种、株数、高度、盖度及群落高度。调查结束后将小样方中所有草本植物分物种采集地上部分(枝叶)带回实验室进行养分分析。在每个样方内随机选取3个采样点,去除地表枯落物和植物残体后,用直径为5 cm的土钻取地表0—20 cm的土壤,将3个采样点的土壤混合过2 mm筛后在室温下自然风干保存用于测定土壤养分。
1.3 样品分析
草本植物地上部分养分含量为调查样方内所有草本植物地上部分养分含量的加权平均值,每个物种枝叶生物量为加权因子,计算出草地养分含量后再计算其化学计量比值。每个样地中土壤的数据是3个重复样品的平均值。采用重铬酸钾-外加热法测定植物地上部分和土壤的有机碳(SOC)含量[21];采用凯氏定氮法测定土壤和植物地上部分的全氮(TN)含量[22],用钼酸铵比色法测定全磷(TP)[23]。
1.4 功能群的划分
1.5 数据处理和统计分析1.5.1 物种多样性
重要值测度:计算样地内草本植物的物种多样性,计算公式如下:
重要值(IV)=(相对密度+相对盖度+相对高度)/3[26]
(1)
多样性测度:本研究中选用Shannon-Wienner指数(H)、Simpson指数(H′)、Margalef指数(R)和Pielou均匀度指数(Jsw)表征物种多样性[27- 29],计算公式如下:
Shannon-Wienner指数:
H=-∑Pi2lnPi
(2)
Simpson指数:
(3)
Margalef指数:
克氏原螯虾(Procambarus clarkii)俗称克氏螯虾、淡水龙虾、小龙虾,原产于美国中部和东部、墨西哥以及古巴,是最具食用价值的淡水龙虾品种,年产量占整个淡水龙虾产量的70%~80%。在我国,小龙虾人工养殖和消费的数量很大。目前,在对虾的加工中主要以冻虾仁为主,在加工中产生大量的虾头、虾壳等下脚料,约占虾体质量的30%~40%[1],如不及时处理,就会造成环境污染。
R=S
(4)
Pielou均匀度指数:
Jsw=H/lnS
(5)
式中,Pi指样方中第i种物种的相对重要值,S指样方中的总物种数。
1.5.2内稳态指数
内稳态指数(Homeostasis index,H)采用生态化学计量内稳性模型计算[30]:
(6)
式中,y表示样地中植物地上部分的C、N、P含量及其化学计量比C∶N、C∶P和N∶P,x表示对应的土壤中C、N、P含量及其化学计量比,c是常数。为方便统计,多用1/H来衡量内稳性的强弱[31],上式中1/H可表示为lg (x)和lg (y)回归方程的斜率。按照Persson等[32]的分类,当模型模拟的结果显著时,将1/H划分为四个类型:0<1/H<0.25表示内稳态;0.25<1/H<0.5 表示弱内稳态;0.5<1/H<0.75表示弱敏感态;1/H>0.75表示敏感态。而当模型模拟的结果不显著时,则认为存在绝对稳态。
1.5.3统计分析
采用Microsoft Excel 2010进行数据整理和计算;用SPSS 20.0进行统计分析,采用单因素方差分析(One-Way ANOVA)检验不同恢复年限植物和土壤化学计量特征的差异,并用Duncan法进行多重比较,显著性水平为P< 0.05;同时用逐步多元回归(STEPWISE) 方法分析不同恢复年限群落物种多样性指数与植物地上及土壤化学计量特征之间的关系;用Origin 2018作图;采用Canoco 5进行冗余分析(RDA)。
2 结果分析
2.1 退耕还草地群落特征的动态变化
2.1.1植物功能群组成的变化
各恢复年限中,物种组成比例最大的是杂草类,其次是禾草类,比例最小的豆科类,仅在恢复35 a时豆科类的比例高于禾草类(21.6% > 16.9%)(图1)。根据重要值大小发现,在植被恢复前期(恢复8 a),禾草类植物在群落中占优势,其次是杂草类,随着恢复年限的增加,杂草类逐渐在群落中占优势,在植被恢复后期(恢复35 a),重要值表现为杂草类(0.8)> 豆科类(0.11)> 禾草类(0.09)。根据不同光合途径(C3和C4)来看,在整个恢复阶段,C3植物种类在群落所有植物种中所占的比例均大于C4植物,且随着恢复年限的增加,C3植物的比例逐渐增大,而C4植物的比例逐渐减少。物种重要值的变化跟C3、C4植物种数分别占总物种数的比例呈现相同的趋势。
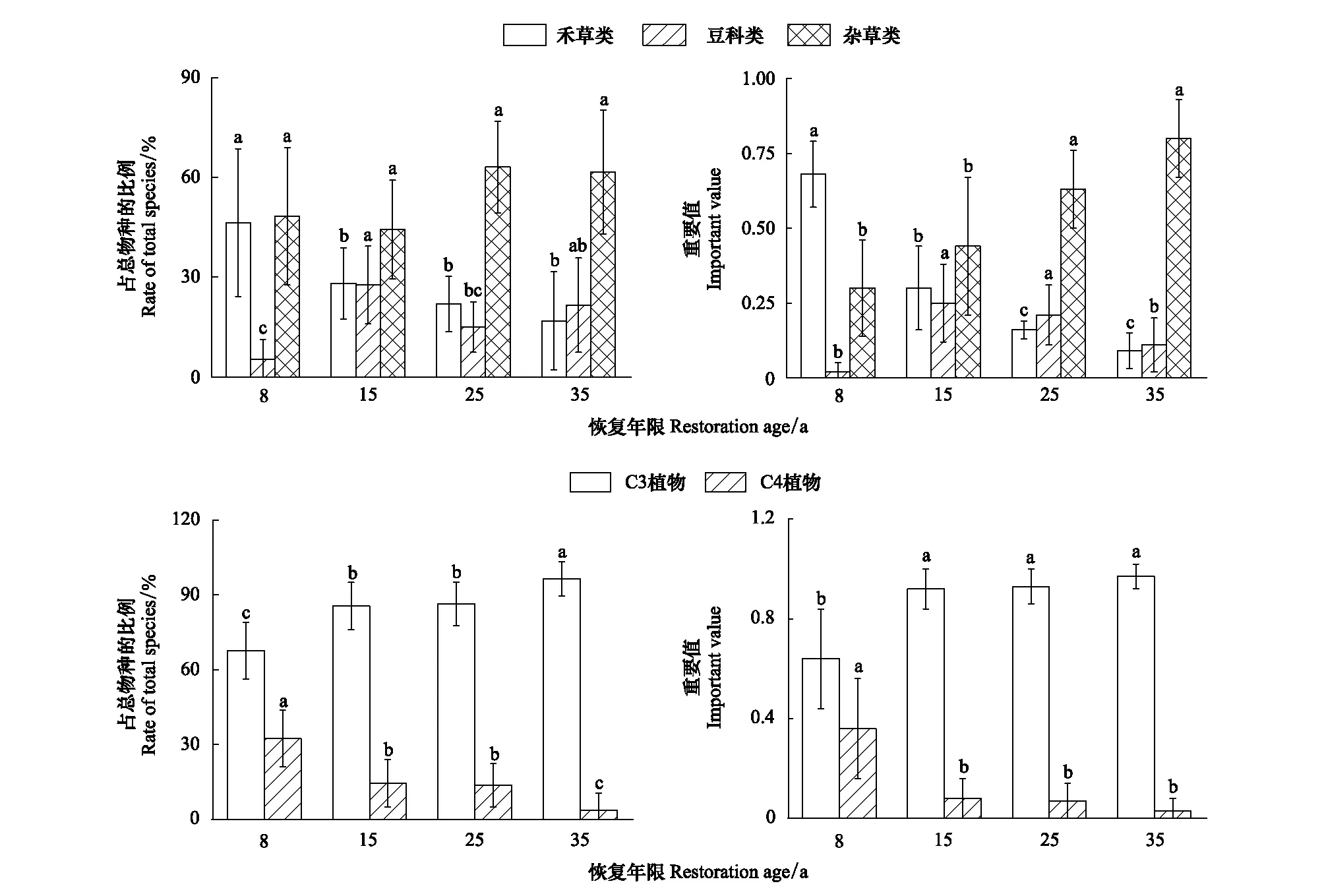
图1 退耕还草地植物功能群组成随恢复年限的变化Fig.1 The changes of the functional group composition with the recovery years since returning farmland to grassland不同的小写字母表示不同恢复年限之间差异显著(P < 0.05)
2.1.2植物群落特征的变化
随着草地恢复,群落中恢复后期的铁杆蒿(T.vulgare)、长芒草(S.bungeana)和北京隐子草(Kengiahancei)逐渐代替前期的冰草(Agropyroncristatum)成为群落的优势种(表1);植物地上部分的生物量以及群落高度、群落盖度都呈现逐渐上升趋势,在整个恢复阶段,生物量、群落高度及群落盖度分别增加了60%、34%和53%(表1);群落植物多样性指数随着恢复年限的变化而变化,具体来看,Margalef指数、Shannon-Weiner指数、Simpson指数和均匀度指数在恢复前期经过一段时间的增长后于恢复中后期(15—25 a)达到最大值,分别为8.22、1.63、0.73和0.82,至恢复后期又呈现下降的趋势,因此植物多样性指数总体呈现“抛物线”趋势。

表1 退耕还草地植物群落特征的变化(平均值±标准差)Table 1 The changes of plant community characteristics since returning farmland to grassland (mean ± standard deviation)
2.2 退耕还草地植物-土壤化学计量特征的变化
恢复年限对植物地上部分的C、N含量有极显著的影响(P< 0.001),对C∶N、N∶P有显著影响(P< 0.05)(图2)。随着恢复年限的增加,植物地上部分C、N、P含量呈现增长趋势,其中,植物地上部分C含量在恢复35 a后达到最大值(434.95 g/kg);N含量在农田阶段最高(2.29 g/kg),经过一个短暂的降低后又开始缓慢上升;P含量从农田开始也经过一段时间的降低后在恢复25 a时达到最高(1.23 g/kg);而植物地上部分的C∶N、C∶P以及N∶P呈现降低趋势,C∶N、C∶P在恢复8 a后达到最大值,N∶P在农田阶段时最高。
恢复年限对土壤的P含量以及C∶N有显著的影响(P< 0.05)(图2)。整体上,土壤C、N、P含量呈现增加趋势,其中,C、N、P含量的最大值均出现在恢复35 a,分别为10.94、0.07和0.06 g/kg;土壤C∶N整体上随草地恢复年限的增加而降低;C∶P基本保持不变,而土壤N∶P随着草地恢复整体呈增加趋势,在恢复35 a达到最大值。
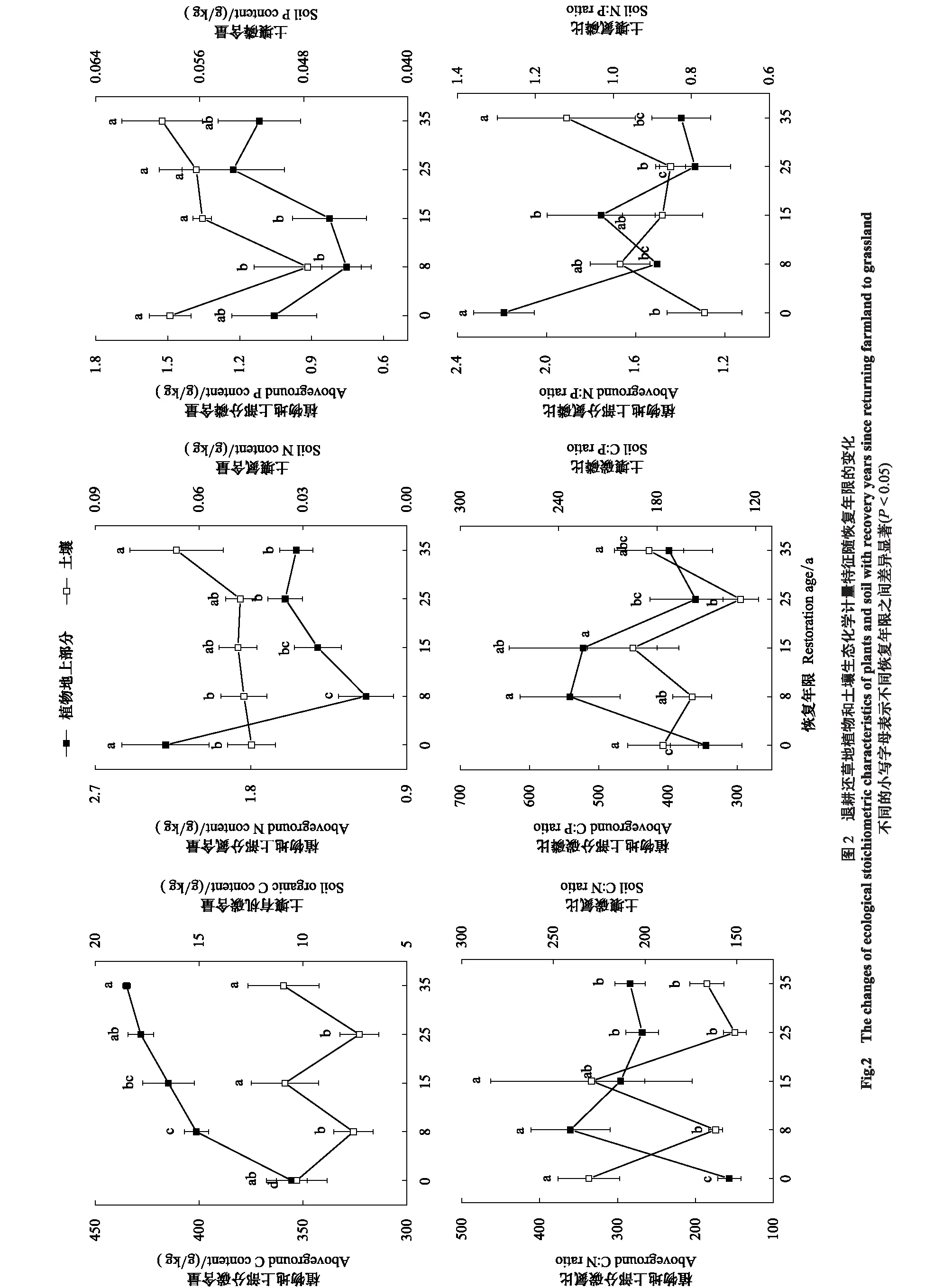
2.3 退耕还草地植物生态化学计量的内稳性特征
为了探究植物生态化学计量内稳性,本研究分析了植物地上部分C、N、P含量及其化学计量比跟相应的土壤资源之间的关系(图3)。结果表明,lg (CA)与lg (CS)、lg (NA)与lg (NS)、lg (C∶NA)与lg (C∶NS)以及lg (C∶PA)与lg (C∶PS) 之间利用稳态性模型模拟结果不显著(P> 0.05),因此认为植物地上部分的C、N、C∶N和C∶P在不同的恢复年限存在绝对稳态;而lg (PA)与lg (PS)和lg (N∶PA)与lg (N∶PS) 之间的模拟结果显著(P< 0.05),其中,lg (PA)与lg (PS)的回归方程斜率(1/H)为1.75,表明植物地上部分P属于敏感态指标,而lg (N∶PA)与lg (N∶PS)的回归方程斜率(1/H)为-0.66,因此以1/H的绝对值来表征植物地上部分N∶P的内稳性,表明其属于弱敏感态指标。

图3 植物生态化学计量的内稳性特征Fig.3 Internal homeostasis characteristics of plant ecological stoichiometry“A”:植物地上部分;“S”:土壤
2.4 植物多样性与植物、土壤化学计量特征的相关性分析
RDA分析结果显示(图4),群落物种多样性指数与植物和土壤C、N、P及其化学计量比在第1、2轴上的解释率分别为61.2%和23.1%。群落高度与植物地上部分C∶N、C∶P及土壤N含量等因子正相关,其中,植物地上部分N含量、P含量、C∶N和C∶P达到显著水平(P< 0.05);群落盖度与土壤C含量、N含量、P含量、C∶P、C∶N及植物地上部分C含量等因子均正相关,仅土壤C含量达到显著水平(P< 0.05);Shannon-Weiner指数与Simpson指数均与植物地上部分N含量、P含量及土壤P、C∶N等因子正相关,与植物地上部分N含量、土壤P的相关性较大,其中Simpson指数与土壤P显著相关(P< 0.05);均匀度指数与土壤N含量、N∶P、C∶P以及植物地上部分C含量等因子正相关;Simpson指数与植物地上部分N含量、P含量等因子正相关。
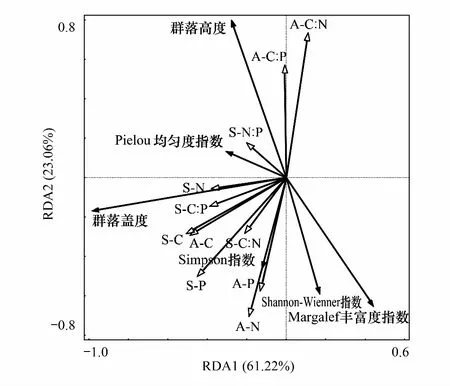
图4 植物多样性与植物、土壤化学计量特征的RDA分析Fig.4 RDA of plant diversity and stoichiometric characteristics of vegetation and soil“A”:植物地上部分;“S”:土壤;C∶有机碳;N∶全氮;P:全磷;C∶N∶碳氮比;C∶P:碳磷比;N∶P:氮磷比
为揭示各因子对植物多样性指数影响的相对重要性,对RDA分析的因子进一步进行逐步回归分析表明(表2),影响群落高度的主要因素是植物地上部分C∶P和土壤N含量;影响群落盖度的主要因素是土壤C含量和植物地上部分C含量;影响Shannon-Weiner指数的主要因素是植物地上部分C含量和C∶N;影响Simpson指数主要因素是植物地上部分C含量和土壤P含量。

表2 植物和土壤化学计量特征影响植物多样性的多元线性回归分析Table 2 Multi-factors linear regression between stoichiometric characteristics of plant and soil and plant diversity
3 讨论
3.1 退耕还草地植物群落的变化
随着退耕还草地恢复年限的增加,群落盖度和群落高度均逐渐增大,而群落植物多样性指数均呈现不同程度的变化,产生这种变化的原因在于随着恢复年限的增加,群落结构的变化以及土壤理化性质的改善影响植物的生长和种类组成,进而影响植物的多样性特征[33]。根据不同恢复年限群落中植物功能群(图1)的变化特征和物种多样性指数(表1)的变化规律,可将研究区草地自然恢复的过程大致划分为3个阶段:
第一个阶段为农田至恢复8 a左右。杂草类植物在群落所有物种中所占的比例最大,而禾草类植物冰草(A.cristatum)在群落中属于优势种,并伴有少量的豆科植物。根据不同的光合途径来看,C3植物占绝对优势,而C4植物的比例在整个恢复阶段中最高,从植物生理特征角度分析,由于C4植物具有较高的光饱和点以及较低的CO2补偿点等特点,因此其在应对相同的外界资源时往往对水分及养分等重要资源的利用效率高于C3植物[34],因此在该阶段中,草地生态系统会以较多的C4植物作为抵抗策略来维持自生的稳定发展,但此时植物种类不多,多样性指数较低。
第二个阶段为草地恢复15—25 a左右。随着恢复年限的增加,到了恢复中后期,由于环境条件趋于稳定,物种种类及其数量明显增多,包括豆类植物,但C4植物的比例逐渐减少,最终形成铁杆蒿(T.vulgare)等多年生草本群落,该结果与秦伟等[35]的研究结果基本一致。因此该阶段植物的Simpson指数、Shannon-Weiner指数、Simpson指数和均匀度指数均呈现增长趋势。
第三个阶段为草地恢复35 a左右。在第二阶段以后,由于物种增多,种间竞争增强,导致个别物种逐渐占据优势地位而部分种群逐渐退化并消失,因此到了该阶段,植物的Simpson指数、Shannon-Weiner指数、Simpson指数和均匀度指数下降,而群落优势种又开始向少数物种集中,此时形成了铁杆蒿(T.vulgare)-长芒草(S.bungeana)-北京隐子草(K.hancei)群落。李裕元等[11]在对乔木层、灌木层和草本层的研究中也发现了这一演替趋势,反映了植物在草地恢复过程中由于环境变化和种间竞争而导致种群结构的变化。
3.2 退耕还草地植物和土壤化学计量特征动态变化
土壤是植物生长发育的基础,其养分供应量对植物的生存和生长起到至关重要的作用,同时,植物枯枝落叶经过微生物的作用可将部分养分归还土壤,因此,植物-土壤系统中C、N、P的积累和转化是陆地生态系统元素循环的基础和核心环节[36]。本研究中,随着植被恢复,植物地上部分C含量增高,这与现有研究结果一致[37],说明在草地恢复过程中植物地上部分持续积累富含C的结构性物质。而N、P含量总体上呈现增加趋势,高德新等[38]在对黄土高原刺槐(R.pseudoacacia)、柠条(CaraganaKorshinskii)和草地的研究中也发现随着恢复年限的增加草地植物叶片的N、P含量显著增加。植物地上部分N含量的增加可能跟豆科类植物根系的固氮作用相关[39],而植物地上部分P含量随着恢复年限的增加而增加可能与杂草类植物的增加有关。寿命短、生长快的杂草类植物,其叶片通常需要更高的P含量来保证其细胞内有大量的核糖体,进而高效的合成植物快速生长所必需的蛋白质,而其高N含量保障了合成蛋白质所需要的原料[40- 41]。本研究中土壤C含量随着恢复年限的增加而增加,有大量的研究表明土壤C含量随恢复年限的变化而增加[42- 44],其原因可能是植物群落组成和结构随着恢复年限的增加不断变化,凋落物以及根系的数量和质量逐渐增多,因此,土壤物理、化学及生物学性质受到影响,从而影响到了土壤C的矿化过程[45]。另外,土壤N含量也随恢复年限的增加而增加,可能受到凋落物养分释放的影响,说明植被恢复促进了N积累。土壤P含量除在恢复前期呈现下降趋势外,其含量随恢复年限的增加而增加,该结果与辜翔等[43]的研究结果一致,原因在于恢复前期植物对P的高需求和凋落物的低输入导致该时期土壤P含量下降,而恢复中期以后随着植物多样性和生物量的增加,随凋落物归还到土壤中的P增多,导致土壤P含量有所提高。
植物和土壤生态化学计量比是研究恢复生态系统养分限制和平衡状态的重要方法[46]。植物叶片C∶N和C∶P可以用来反映植物的生长状况,其与植物生长的速率呈负相关关系[47]。本研究结果表明,植物地上部分C∶N和C∶P随恢复年限的增加总体呈现先增大后减小的趋势,主要是因为在恢复前期植物地上部分C含量增加的过程中N、P含量都在减少,而草地恢复后期生长相对较快,植物N、P含量均开始增加。Koerselman和Meuleman[48]提出:当植物N∶P > 16时,植物的生长受P限制;N∶P < 14时,植物的生长受N限制;而Güsewell则把10和20作为N∶P阈值的临界点[49]。尽管存在这些分歧,但本研究中,植物地上部分N∶P总体上呈现下降趋势,且比值远低于10,说明该地区植物在恢复过程中主要受N限制,可能是由于土壤N含量在草地恢复期间持续消耗所导致的。作为植物生长发育的基础,土壤的C∶N、C∶P和N∶P可作为评价其养分状况的重要指标[50]。本研究发现,土壤C∶N总体上随着草地恢复呈现下降趋势,C∶P基本不变,而N∶P呈现增大趋势,可能是因为植物通过吸收和释放土壤中的C、N、P进而改变其在土壤中的比例,亦或因为群落中物种组成和结构的变化引起土壤中C、N、P的不平衡性,进而引起土壤C∶N、C∶P、N∶P的变化[51]。
3.3 退耕还草地植物化学计量内稳性动态变化及影响因素
草地恢复后,当土壤养分等外界环境因子发生改变时,植物通过维持内部化学组成的相对稳定,使其内部环境的变化保持在较小的范围,这是一个动态的平衡,即植物形成了内稳态机制[30]。本研究中,在不同的恢复年限,植物地上部分的C、N、C∶N和C∶P存在绝对稳态,说明这些指标随着土壤养分变化而变化的趋势不显著,但是植物地上部分P属于敏感型指标,受土壤P含量变化的影响较大,而其N∶P属于弱敏感型指标。该结果与陈婵等[37]对中亚热带植物内稳性特征的研究结果不一致,原因可能是研究地区环境差异导致植物的限制性养分元素不同,湖南省中东部土壤主要受P限制。限制性养分元素稳定性假说认为:“由于生理和养分平衡的制约,限制性养分元素在植物体内的含量具有相对稳定性,对环境变化的响应也较为稳定”[30]。本研究区域土壤中N含量相对较低,因此,地上部分N的内稳性大于P。另外,本研究发现植物地上部分N∶P的内稳性指数为负值,与以往通过野外试验测算出有机体的化学计量内稳性指数为负值的结果一致[32, 52]。在这种情况下,一般都以内稳性指数1/H的绝对值来表示该有机体化学计量内稳性,因此植物地上部分N∶P属于弱敏感态指标,主要原因是草地恢复过程中,群落环境及植物组成和结构的变化,使得植物地上部分N∶P很容易随土壤养分条件的变化而变化。
3.4 退耕还草地植物多样性的变化及其影响因素
植物多样性可以反映环境与植物群落的关系[53],通过对其进行深入研究可以了解群落物种的组成、群落变化及其发展趋势[54]。总体来看,多样性指数之间大多显著相关,且植物和土壤C、N、P及其化学计量比对植物多样性存在一定的影响。本研究中,植物地上部分C∶P(P< 0.05)和土壤N对群落高度有较强正效应。由于黄土高原属于干旱半干旱地区,土壤水分缺乏,使得N、P含量较低,有研究表明植物地上部分C∶P可以用来表征植物的磷素利用效率[55],土壤N含量和植物地上部分P素利用效率越高,越有利于植物的生长,群落高度也随之增加。植物地上部分C含量和土壤C含量(P< 0.05)对群落盖度有积极影响,这是因为土壤和植物地上部分之间相互影响,不断进行物质交换,使得群落盖度也随之改变。Shannon-Weiner指数是群落复杂程度的直观反映,其值越大,群落的复杂程度就越高。在本研究中,植物地上部分C含量及其C∶N比对该指数有消极影响,该结果与杨祥祥等[15]对沙地植被C、N、P化学计量特征与物种多样性关系的研究结果一致。一般认为,C作为有机体的骨架元素,不作为影响植物多样性的关键因子[56],但研究结果与此相悖,具体原因还需进一步研究。Simpson指数可以表征群落中物种的优势程度,群落中物种数越多,各种个体分配越均匀,则指数越高。研究结果显示,植物地上部分C含量和土壤P含量与Simpson指数有较强相关性,其中植物地上部分C含量与其负相关,而与土壤P含量显著正相关(P< 0.05),该结果与杨祥祥等的研究结果一致[16]。在干旱地区,土壤通常表现为P受限制,导致了植物的生长缺乏P的供应,当土壤中P含量增多时,就会使Simpson指数增大。Simpson指数是指该草地群落中物种数目的多少,均匀度指数是估计群落中物种分布的均匀程度,本研究中,植物地上部分和土壤C、N、P含量及其化学计量比均与这两个指数未表现出显著相关性,该结果与现有的研究结果不一致[18,57- 58],其原因可能是植物地上部分和土壤中各个养分因子并不单独作用,它们之间存在影响和制约关系,且研究区域中的气候环境也制约各个因子的作用范围[59],因此使研究结果出现差异。综上所述,植物地上部分C、N含量及其C∶P以及土壤C、N、P含量均对植物多样性产生重要的影响。
4 结论
(1)随着恢复年限的增加,植物生物量、群落高度和群落盖度都呈现增加趋势,而植物多样性总体上呈现“抛物线”式变化趋势,即在恢复前期上升,中后期达到最高,恢复后期又开始下降,说明该区域草地恢复符合植被恢复的一般规律。
(2)在整个恢复阶段,植物地上部分的C、N含量及其C∶N和C∶P存在绝对稳态,P属于敏感性指标,而植物地上部分N∶P属于弱敏感性指标,可能因为干旱环境中土壤中P相对缺乏,其维持内稳态的能力也因此受限制,进而影响到N∶P的内稳性,使其内稳态属于弱敏感性。
(3)随着植物地上部分C含量和C∶N的增加,群落的Shannon-Weiner指数减小;植物地上部分C含量和土壤P含量是影响Simpson指数的主要因子,其中该指数与植物地上部分C含量呈现负相关关系,而与土壤P含量显著正相关,说明植物地上部分和土壤生态化学计量特征对植物多样性有重要的影响。
