横断山不同地区大绒鼠冬季体重调节
2021-01-13韩春艳王政昆朱万龙
韩春艳 贾 婷 杨 忌 王政昆 朱万龙*
(1.云南省高校西南山地生态系统动植物生态适应进化及保护重点实验室,云南师范大学生命科学学院;生物能源持续开发利用教育部工程研究中心;云南省生物质能与环境生物技术重点实验室,昆明,650500;2.云南经济管理学院,昆明,650106)
小型哺乳动物通过调节能量的摄入和消耗来调节体重以适应不同环境的变化[1-2]。动物的消化道具有容纳、处理食物和吸收营养物质的能力,食物的质量等外界环境因素影响消化道长度[3]。研究表明,小型哺乳动物消化道长度随季节变化,是表型可塑性的一种[4],指动物为适应食物质量和环境温度,消化道在形态、结构和生物化学上发生的可逆性变化[5],是动物适应环境的重要特征[6]。研究发现,小型哺乳动物会通过降低体重和改变消化道长度来提高在低温和食物匮乏条件下的生存率[7]。
瘦素是白色脂肪分泌的一个16kDa的蛋白类激素,含167个氨基酸[8],是一个重要的脂肪细胞因子[9],中枢神经系统中瘦素可抑制下丘脑弓状核促食神经肽Y(neuropeptide Y,NPY)和刺鼠相关蛋白(agouti-related peptide,AgRP)的基因表达,同时,刺激弓状核抑食神经肽前阿黑皮素(pro-opiomelanocortin,POMC)和可卡因和安非他明调节转录肽(cocaine and amphetamine regulated transcript peptide,CART)的表达,从而抑制摄食、促进产热、降低体重[10]。研究发现,高水平的瘦素可以抑制食欲,增加能量消耗[11],低浓度的瘦素促进小型哺乳动物的进食,影响动物的体重及能量平衡[12]。
横断山位于中国西南部,青藏高原的东南部,作为长江上游重要的生态屏障区,是中国珍稀濒危动植物的避难所,也是世界上生物多样性最丰富的地区之一[13],对周边区域乃至中国西部地区气候和生态环境有着深远的影响[14]。该地区随海拔升高,气温降低,降水增加,具有干湿两季分明,年温差小,日温差大,立体气候显著等特点[15]。大绒鼠(Eothenomysmiletus)为横断山固有种[16],主要分布于我国云南省的横断山地区中部,营地表浅层洞道生活,多在夜间活动,以含水量高的鲜嫩植物、草的根部以及茎部为主要食物[17]。本研究组之前的研究结果表明,夏季香格里拉和德钦地区的大绒鼠体重显著小于丽江、剑川和哀牢山3个地区,但是肝脏和小肠重显著高于其它3个地区,表明不同地区大绒鼠通过调节体重和消化道形态等生理指标来适应不同的环境条件[18]。而冬季横断山区的环境因子更为胁迫,因此本研究采集冬季横断山区自北向南分别从德钦、香格里拉、丽江、剑川和哀牢山5个不同地区的大绒鼠,测定和比较其体重、身体组成、消化道、血清瘦素以及下丘脑神经肽含量的变化,为探究大绒鼠在冬季对横断山不同地区适应机制提供证据。
1 研究方法
1.1 实验动物
实验动物于2019年冬季分别捕自横断山地区的5个地点,自南向北依次为:哀牢山、剑川、丽江、香格里拉和德钦。实验动物均为非繁殖期的健康成年个体,每组6只动物。
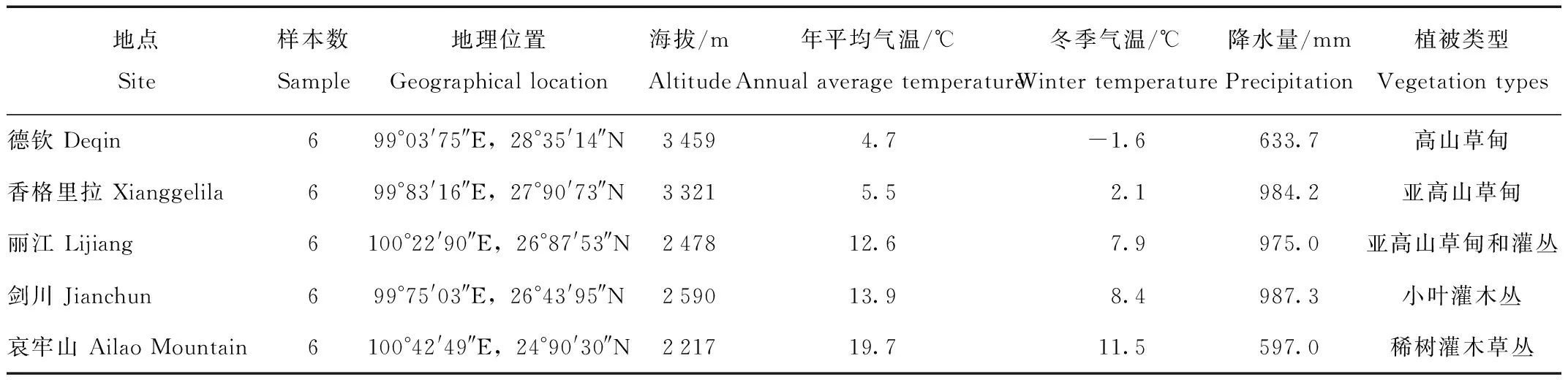
表1 横断山区大绒鼠采样点Tab.1 Features of the sampling site in E.miletus from Hengduan Mountains
1.2 动物处理
用捕鼠笼捕捉动物,当天在地方病防治站称量体重后处死,记录繁殖状态等。将心、肺、肝、脾、肾、胃、小肠、大肠及盲肠完整取出,仔细剔除黏连的结缔组织,在滤纸上蘸干,称量鲜重(精度为0.1 mg)。取血清和下丘脑,血清瘦素浓度和下丘脑神经肽基因表达量分别用放射免疫法和实时荧光定量方法测定,详见Zhu等[19]2017研究报道。
1.3 数据分析
数据采用SPSS 22.0软件进行分析,所有数据均符合正态分布。本研究组之前发现大绒鼠雌雄之间生理数据差异不明显[20],故数据合并分析。5个地区的各种指标差异性采用单因素方差分析(one-way ANOVA)或协方差分析(ANCOVA)进行分析,以体重作为协变量。结果以平均值±标准差(Mean±SE)表示,其中P<0.05为差异显著,P<0.01为差异极显著。
2 结果
2.1 体重、身体组成及消化道重量
冬季5个地区大绒鼠体重差异极显著(F=22.23,P<0.01,图1),其中香格里拉和德钦地区的大绒鼠体重显著低于其他3个地区。肝脏重量5个地区之间有显著差异(F=3.25,P<0.05,表2),其余身体组成重量均差异不显著(P>0.05)。5个不同地区大绒鼠的胃重有极显著差异(F=4.26,P<0.01),盲肠重量5个地区之间差异显著(F=2.69,P<0.05),其余消化道重量差异不显著。
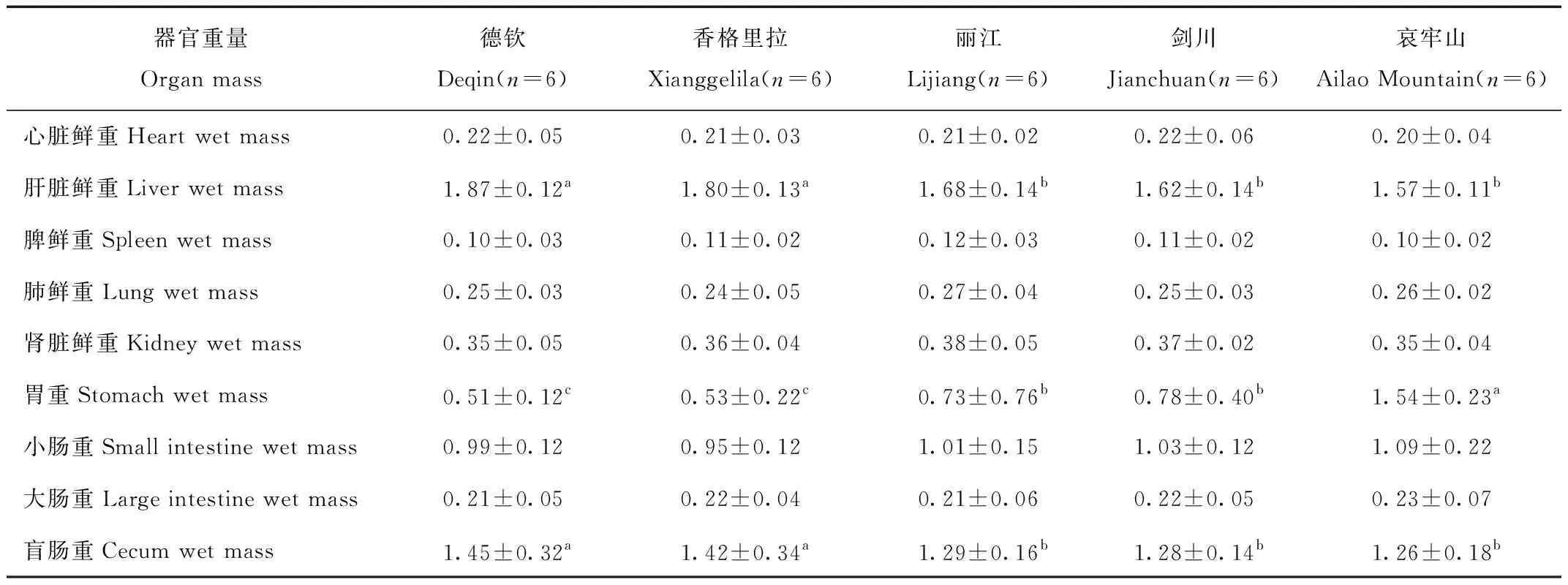
表2 5个地区大绒鼠内脏器官重量和消化道重量变化Tab.2 Changes of body composition and digestive tract masses in E.miletus at five regions
2.2 血清瘦素含量和下丘脑神经肽表达量
冬季5个地区大绒鼠血清瘦素含量差异极显著(F=8.69,P<0.01,图2),其中香格里拉和德钦地区的大绒鼠血清瘦素含量显著低于其他3个地区。冬季5个地区大绒鼠下丘脑神经肽NPY和AgRP表达量地区之间差异显著(NPY:F=4.21,P<0.05;AgRP:F=2.89,P<0.05),但是POMC和CART表达量没有地区差异。
3 讨论
小型哺乳动物的体重变化受环境温度、海拔及食物的影响,动物为适应不同的栖息环境,形成各自不同的获能策略[21]。在自然条件下,小型哺乳动物必须通过行为或生理调节来满足能量需求[22]。本次采样点从北向南分别在德钦、香格里拉、丽江、剑川和哀牢山,年平均气温和采样期气温由北向南依次升高,本研究组之前的研究结果表明夏季横断山5个采样地大绒鼠体重由南向北依次降低[18],这可能和不同地区环境因素的渐变有关。本研究中,德钦和香格里拉的大绒鼠体重明显低于其他3个地方,这可能和植被类型与高海拔低温度有关。
肝脏是小型哺乳动物的产热的器官之一[23]。在本研究中,德钦和香格里拉两个地区的大绒鼠肝脏重量明显高于其他3个地区,表明大绒鼠可能通过增加肝脏重量,增加产热,来抵抗两个地区的低温环境。胃作为小型哺乳动物消化道的重要组成部分,其形态常因食性的不同而变化[24]。动物胃的大小与温度、食物质量和繁殖状态等很多因素有关[25]。胃越大,储存的食物越多,食物在胃里消化时间变长,增加食物消化吸收的效率[26]。本研究组之前的研究结果表明夏季哀牢山大绒鼠较剑川、丽江、德钦和香格里拉地区的胃重大[18]。冬季哀牢山大绒鼠胃重高于其他4个地区,剑川和丽江的大绒鼠胃重也显著高于德钦和香格里拉地区,这可能是与温度和植被类型有关。由于冬季哀牢山、剑川、丽江地区大绒鼠所食用的草根及嫩叶在冬天未被冰雪覆盖,大绒鼠有充足的食物资源和相对较大的胃,具有重要的生态学意义,它既可提高觅食效率和获得足够食物,又可减少暴露时间,从而降低被捕食的风险[27]。盲肠是纤维素的发酵部位,纤维素经盲肠分解后的营养物质主要由大肠吸收[28]。盲肠对于食物质量做出灵敏反应,当在低温条件下,食物质量下降,纤维素含量升高时,盲肠会变重[29-30]。
瘦素由白色脂肪组织分泌的,作为一种信号通过血液循环作用于下丘脑[31]。动物在低温时瘦素降低,低浓度的瘦素防止进一步消耗能量。保持能量输入和输出的平衡[32]。本研究组之前结果表明,低温诱导大绒鼠降低瘦素,低浓度的瘦素促进大绒鼠进食,增加能量收入,从而产生热量适应低温环境[33]。本研究中,香格里拉和德钦的血清瘦素含量明显低于其他3个地方,与之前结果一致,大绒鼠可能通过降低瘦素和体重适应高海拔低温度地区(香格里拉和德钦)恶劣的环境。通过比较4种小型哺乳动物不同季节的血清瘦素和下丘脑神经肽表达量变化(表3),可见4种动物下丘脑NPY表达量均在冬季增加,大绒鼠和高山姬鼠(Apodemuschevrieri)的AgRP表达量冬季比夏季高,其余两种的AgRP不变。但是5个地区大绒鼠的下丘脑抑食类神经肽POMC和CART相对表达量没有显著差异,可能和低温环境中动物的产热增加有关,因为POMC和CART的表达水平越高,冬季产热就越明显[19]。这和中缅树鼩(Tupaiabelangeri)的变化不同,但是和其他物种变化相似(表3),说明不同季节不同物种的下丘脑神经肽调节具有种属特异性。本研究中,香格里拉和德钦NPY和AgRP相对表达量显著高于其余其他3个地区,表明高海拔低温度地区大绒鼠通过增加NPY和AgRP相对含量,增加食欲,促进大绒鼠的进食,增加产生热量,抵抗低温带来的胁迫[34]。以上结果说明血清瘦素和下丘脑神经肽表达量可能参与了大绒鼠的体重调节。
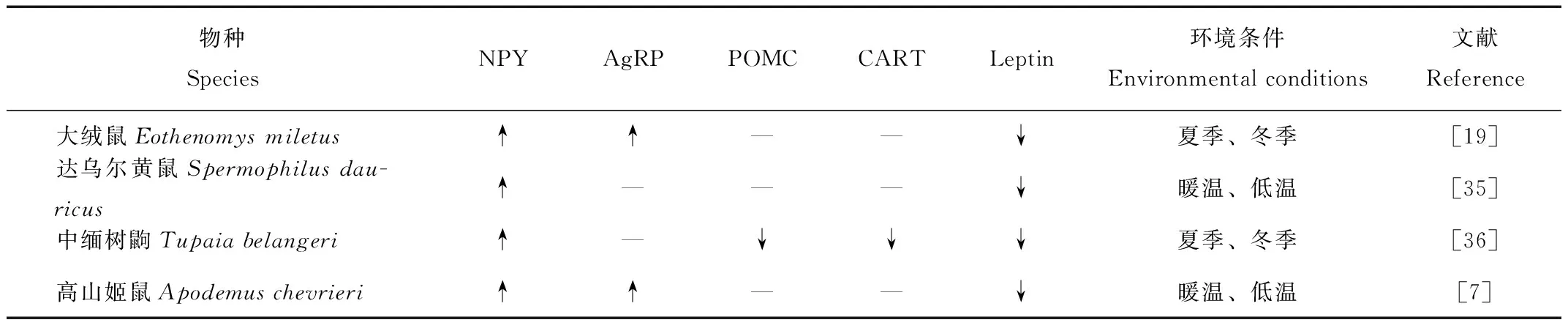
表3 4种小型哺乳动物不同环境条件下的血清瘦素和下丘脑神经肽表达量变化Tab.3 Changes of serum leptin levels and hypothalamic neuropeptide expressions in four small mammals under different environmental conditions
综上所述,冬季横断山不同地区的大绒鼠通过改变体重、肝脏重量、大肠重量、盲肠重量、血清瘦素、下丘脑促食类神经肽相对表达量,最终适应该地区不同的环境变化。
