水位波动的幅度与频率对刺苦草(Vallisneria spinulosa)生物量和形态特征的影响*
2021-01-12丁明明金斌松陈家宽
丁明明,黎 磊,2 ,金斌松,2,陈家宽,3
(1:江西省流域生态演变与生物多样性重点实验室,南昌大学生命科学研究院流域生态学研究所,南昌大学生命科学学院,南昌 330031) (2:江西鄱阳湖湿地保护与恢复国家长期科研基地,江西鄱阳湖湿地生态系统定位观测研究站,南昌 330031) (3:复旦大学生物多样性科学研究所,生物多样性与生态工程教育部重点实验室,上海 200438)
水位波动是影响湖泊生态系统结构与功能的主要干扰因子[1-2]. 湖泊中的水位波动有很强的时空分布特征[3-4],主要表现在水位的高低及其变动范围、频率、发生时间和持续时间等[5-7]. 近年来,气候变化(降水量、降水频率和单次降水强度的变化)和人类活动(大型水利工程建设、围垦等)的加剧改变了长江中下游地区许多湖泊水位季节性波动规律[8-11]. 例如,三峡工程运行造成鄱阳湖水位自然节律的明显改变[12,13],从而影响湖泊水生植被的生长、分布与演替[2,14-15].
水位波动幅度和波动频率是水文情势重要的组成部分[2]. 波动幅度是指一个周期内最高和最低水位之间的水位变差,波动频率是指特定时间内水位变化的周期数[2,4,16-18]. 水位波动幅度可能影响水压、水体CO2浓度、O2浓度和光照强度[2,4,19-21]. 例如,随着水位变幅增加,湖泊中高水位和低水位交替出现,在高水位条件时水体中的O2浓度减小、CO2浓度增加、水压增强、沉水植物的光截获量急剧下降[4,22]. 因此水位波动幅度的变化对沉水植物的生长、光合作用和能量利用效率有着较大的影响. 随着水位波动频率增加,水生植物受到的干扰强度大大增加,可能引致养分流失和组织损伤(如茎、叶、匍匐茎断裂或植物体被连根拔起)[23-25]. 水位波动频率增加还将频繁地改变水体光通量,对水生植物的光合结构造成破坏[2,26-28]. 此外,水位波动可引起底泥再悬浮、改变水体与沉积物的营养盐浓度和氧化还原电位[7,29-30],进而影响植物的生长和分布[7,29].
一些研究表明,较高的水位波动频率抑制水生植物生长,如喜旱莲子草(Alternantheraphiloxeroides)[31]. 然而,也有研究表明,较低的或中度水位波动频率对水生植物的建植和生长影响较小或没有显著影响[2,32]. Wang等[33]发现水位波动频率的变化不会对金鱼藻 (Ceratophyllumdemersum) 和脆轮藻 (Charafragilis) 的生长产生影响. 适度的水位波动幅度有利于黑藻 (Hydrillaverticillata) 和水车前 (Otteliaalismoides) 的生物量积累,而较大的水位变幅会产生抑制作用[34-35]. 以往关于水位变化与水生植物关系的研究大多集中在水深、水淹强度或水位波动幅度单一因素的影响[18,36-38],而有关水位波动幅度和波动频率两个因素综合影响的研究较少,尤其是对植物地下器官生长发育影响的研究更少. 目前,关于长江中下游的广布种和特有种刺苦草 (Vallisneriaspinulosa) 对水位波动的响应机制还缺乏相关研究.
我国长江中下游湖泊多浅水通江,植被以沉水植物为主,由于降雨或水闸的调节作用,水位变化十分频繁. 刺苦草是长江中下游湖泊的优势物种,一般分布在20~180 cm的水深范围内. 该物种叶基生,呈带状,光补偿点低,能够耐受一定程度的弱光环境. 鉴于长江中下游湖泊水位波动幅度为0.5~1 m、水位变化速率为5~10 cm/d的情况经常出现[22],本研究通过室外模拟实验,选取2个水位波动幅度(±30 cm和±60 cm)和2个水位波动频率处理(12 d或24 d为一个水位波动周期)(逐渐改变水位),且以恒定水位(90 cm)作为对照,回答以下问题: 1) 水位波动(波动幅度与波动频率)是否对刺苦草生长(生物量)有负面影响?2) 水位波动对刺苦草地上和地下部分形态有何影响?
1 材料与方法
1.1 实验材料
刺苦草为多年生大型沉水植物[39],是鄱阳湖湿地生态系统的重要组成部分. 它不仅能维持湖泊的清水稳态[40],还能为鱼类提供产卵场和栖息地、为浮游动物和无脊椎动物提供避难所、为植食性越冬水鸟提供食物[41-43]. 黎磊等[44]的研究表明,刺苦草可以在0.5~1.5 m水深范围生长发育,不同水深条件下刺苦草外部形态和繁殖特征存在明显差异. 在生长季节,刺苦草通过匍匐茎克隆生长,分株呈莲座型,叶最长可达2 m;根系发达,由不定根组成,没有侧根. 营养生长期结束后,该物种会产生种子和越冬繁殖体(块茎).
从江西省鄱阳湖国家级自然保护区蚌湖(29°11′~29°18′N,115°55′~116°06′E)收集刺苦草的越冬块茎,将新鲜的块茎放入盛满水的塑料盒,放在4℃的黑暗环境中保存4个月.
1.2 实验设计
本实验在位于庐山市的中国科学院鄱阳湖湖泊湿地观测研究站(29°26′N,116°03′E)的15个混凝土实验水池(2 m×2 m,水深1.6 m)中进行,实验时间为2018年5月22日-8月30日. 实验设置了2个水位波动幅度和2个波动频率,以恒定水位作为对照,设置3个区组. 选取180个鲜重为0.91 g ± 0.12 g(平均值±标准误)的刺苦草块茎,随机种在180个塑料盆(直径23 cm,高17 cm)中,并将这些块茎随机分布在3种水深处理中,以使处理之间的平均块茎质量没有显著差异. 每个栽培盆中添加15 cm厚的基质(采自鄱阳湖的底泥和细沙混匀,体积比=4∶1; 总氮含量为1.0 mg/g,总磷含量为0.16 mg/g,有机质含量为7.6%;n=3). 将这180个栽培盆平均分配到加满水(总氮浓度为1.11 mg/L,总磷浓度为0.01 mg/L,叶绿素a浓度为6.5 μg/L,n=15)的15个实验水池中. 为了方便植物栽培盆的悬挂,在水池上平行放置数根直径为6 cm的镀锌钢管,将种有刺苦草块茎的栽培盆悬挂于水池中进行预培养. 通过挂绳控制预培养水深为60 cm,预培养时间为28 d.
为了模拟洪泛湖泊水位波动的情景,选取5个水位波动处理: (1) 无水位波动(对照);(2) 低幅度低频率;(3) 低幅度高频率;(4) 高幅度低频率;(5) 高幅度高频率(图1). 在对照组中,植株一直淹没在90 cm深的水中(从水面到栽培盆内基质表面的深度),没有水深变化. 在低幅度低频率处理中,水深从60 cm逐渐变化到120 cm,然后又回到60 cm,在实验期间72 d内共实施3次这样的波动(每个波动周期为24 d). 同样在低幅度高频率处理中,水深在60~120 cm之间波动,但每个波动周期为12 d,在实验期间72 d内重复6次波动. 在高幅度低频率处理中,水深从30 cm到150 cm重复变化了3次,最后又回到30 cm(每个波动周期为24 d). 在高幅度高频率处理中,水深从 30 cm到150 cm重复变化了6次,最后又回到30 cm(每个波动周期为12 d). 4个处理的平均水深都是90 cm,因此与水位恒定的对照组(水深90 cm)有可比性. 水池水位维持在最高水位,定期查看植株生长状况. 每7天测定1次光合有效辐射,测试时间固定在中午12:00-13:00之间. 利用Li-1400数据记录器和Li COR UWQ-192SA传感器分别测定水下0、30、40、50、60、70、80、90、100、110、120、130、140和150 cm光合有效辐射(图2).

图1 水位波动处理示意图(对照(无水位波动): 植株的水淹深度一直保持90 cm; 低幅度低频率: 水深按60 cm—120 cm—60 cm的顺序逐渐变化,在实验期间72 d内重复3次波动; 低幅度高频率: 水深按60 cm—120 cm—60 cm的顺序逐渐变化,在72 d内重复6次波动; 高幅度低频率: 水深按30 cm—150 cm—30 cm的顺序逐渐变化,在72 d内重复3次波动; 高幅度高频率: 水深按30 cm—150 cm—30 cm的顺序逐渐变化,在72 d内重复6次波动)Fig.1 Schematic diagram of the five water level fluctuation treatments (Control (no fluctuations): the water depth of the plant remains at 90 cm; small fluctuation amplitude and low fluctuation frequency: water depth gradually changed from 60 cm-120 cm-60 cm,repeated 3 times within 72 days; small fluctuation amplitude and high fluctuation frequency: water depth gradually changed from 60 cm-120 cm-60 cm,repeated 6 times within 72 days; large fluctuation amplitude and low fluctuation frequency: water depth gradually changes from 30 cm-150 cm-30 cm,repeated 3 times within 72 days; large fluctuation amplitude and high fluctuation frequency: water depth gradually changes from 30 cm-150 cm-30 cm,repeated 6 times within 72 days)

图2 不同水深(0、30、40、50、60、70、80、90、100、110、120、130、140和150 cm)下光合有效辐射的变化Fig.2 Variation in photosynthetically active radiation in water column at different water depths (0,30,40,50,60,70,80,90,100,110,120,130,140 and 150 cm)
本实验采用随机区组设计,每5个实验水池为1个区组,每个区组的5个水池分别含1个水位波动处理(对照、低幅度低频率、低幅度高频率、高幅度低频率或高幅度高频率),每个水池内悬挂12个栽培盆(12个植株),因此统计过程中每个处理的样本数量为36株(12株×3个区组),每个池子的数据为12个植株的平均值. 实验过程中,通过向下释放(增加水深)或向上拉起(减少水深)挂绳控制栽培盆的水深,人工模拟水位波动. 在实验期间72天内水深每天变化一次,处理(1)~(5)的栽培盆分别以0、5、10、10和20 cm/d的速度下降或上升.
1.3 形态特征和生物量测定
实验结束时(第100天),取出所有植株. 在水池内洗净,用软毛刷小心刷掉植物表面的附着物,轻轻将植株根部的土壤洗去. 然后分别测定每个植株的地上营养器官特征(株高、分株数、叶片数、比叶面积)、地下营养器官特征(最长根长、根数、匍匐茎数、匍匐茎总长). 将植株的地上(叶、花果)和地下部分(根、匍匐茎、块茎)分开,置于75℃烘箱中烘干至恒重后称量,计算总生物量,记录叶生物量和根生物量. 株高是指植株基部到叶片顶端的最大高度; 比叶面积 (specific leaf area) 是指叶面积与叶干重的比值. 最长根长是指植株基部到根系顶端的最大长度. 总生物量是指完整植株的干重. 利用AM-350叶面积仪 (ADC Bioscientific,Hoddesdon,United Kingdom) 测定叶面积.
1.4 数据分析
数据分析使用SPSS 19.0软件 (SPSS,Inc.,Chicago,IL,US). 利用一般线性模型 (general linear model) 评估区组、水位波动(对照、低幅度低频率、低幅度高频率、高幅度低频率、高幅度高频率)以及二者间的交互作用对生物量积累和形态学参数的影响. 利用线性混合模型 (mixed linear model) 评估水位波动幅度(静止、低幅度和高幅度)、波动频率(低频率和高频率)、二者间的交互作用(以区组作为随机因子)对刺苦草地上和地下生长参数的影响. 如果处理的影响是显著的,再利用Tukey多重比较方法检验处理间的差异. 如果数据方差不齐或不满足正态分布,进行lg转换后再进行分析. 实验中显著度的水平为0.05.
2 结果与分析
2.1 水位波动幅度与波动频率对植物地上营养器官形态的影响
相比于对照组(无水位波动),水位波动处理(低幅度低频率、低幅度高频率、高幅度低频率、高幅度高频率)对刺苦草地上营养器官形态特征(株高、分株数、叶片数和比叶面积)都有显著影响(表1). 与对照组相比,水位波动能够显著降低刺苦草的分株数和叶片数(图3B,3C),但会显著增加其株高和比叶面积(图3A,3D). 然而,不同水位波动幅度(低幅度和高幅度)、波动频率(低频率和高频率)对刺苦草的地上营养器官性状(株高、分株数、叶片数、比叶面积)都没有显著影响,而二者的交互作用对株高有显著影响(表2;P<0.05). 在高幅度高频率条件下,刺苦草株高显著低于其他3个水位波动处理(低幅度低频率、低幅度高频率、高幅度低频率)(表2; 图3A).

表1 水位波动(对照、低幅度低频率、低幅度高频率、高幅度低频率或高幅度高频率)和区组对刺苦草各生长参数的影响(n=180)

图3 刺苦草地上部分形态特征(株高、分株数、叶片数和比叶面积)在5个水位波动处理下的变化(平均值 ± 标准误)(不同的小写字母表示处理间有显著差异(P<0.05))Fig.3 Variation in aboveground morphological traits (plant height, ramet number, leaf number and specific leaf area) of Vallisneria spinulosa in the five water level fluctuation treatments (mean ± SE) (Values with different lowercase letters are significantly different among treatments at the P<0.05 level)
2.2 水位波动幅度与波动频率对植物地下营养器官形态的影响
相比于恒定水位,波动水位(低幅度低频率、低幅度高频率、高幅度低频率、高幅度高频率)对刺苦草根和匍匐茎的形态(最长根长、根数、匍匐茎数和匍匐茎总长)都有显著影响(表1). 总体而言,与对照组相比,水位波动能够显著降低根数、根长、匍匐茎数和匍匐茎总长(图4). 刺苦草的根数和根长均显著受到水位波动幅度(低幅度和高幅度)的影响,但水位波动频率(低频率和高频率)的影响不显著(表2). 在同一个水位波动频率(高频率)处理下,生长在高水位波动幅度下的植株的根长(17.4 cm ± 1.3 cm) 与根数(66 ± 3)显著低于生长在低水位波动幅度下的植株(根长: 20.7 cm ± 0.7 cm;根数: 98 ± 7)(图4A,4B). 然而,不同波动频率和幅度对匍匐茎数、匍匐茎总长度都不存在显著影响(表2; 图4C,4D;P>0.05). 此外,区组对株高与根长存在显著影响(表1;P<0.01,P<0.05).

表2 水位波动幅度(恒定、低幅度、高幅度)和波动频率(低频率、高频率)对刺苦草各生长参数的影响(n=180)
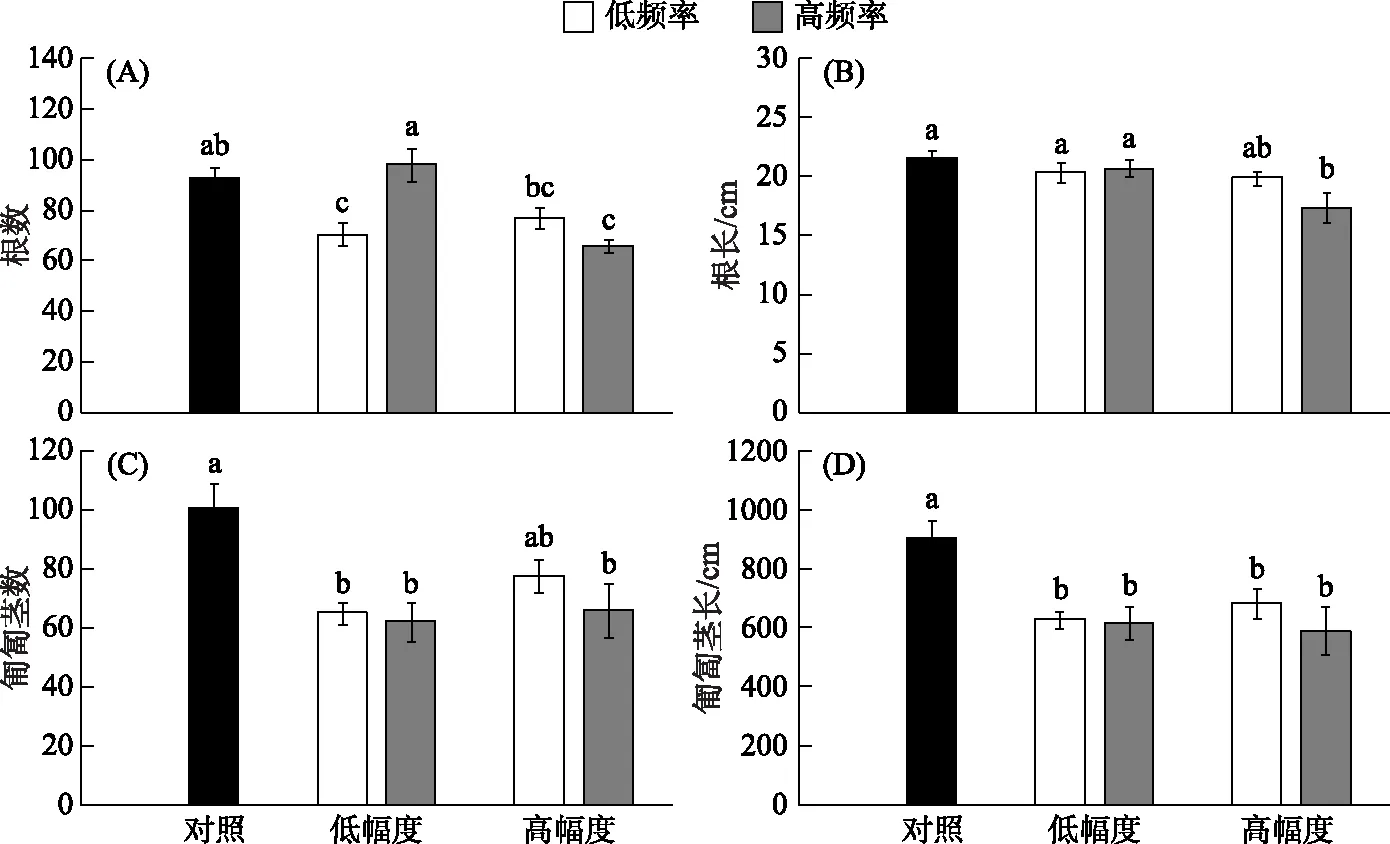
图4 刺苦草根和匍匐茎形态特征在5个水位波动处理下的变化(平均值 ± 标准误)(不同的小写字母表示处理间有显著差异(P<0.05))Fig.4 Variation in morphological traits of roots and stolons of Vallisneria spinulosa in the five water level fluctuation treatments (mean ± SE) (Values with different lowercase letters are significantly different among treatments at the P<0.05 level)

图5 刺苦草总生物量和各部分生物量在5个水位波动处理下的变化(平均值 ± 标准误)(不同的小写字母表示处理间有显著差异(P<0.05)Fig.5 Variation in total and component biomass of Vallisneria spinulosa in the five water level fluctuation treatments (mean ± SE) (Values with different lowercase letters are significantly different among treatments at the P<0.05 level)
2.3 水位波动幅度与波动频率对植物生物量积累的影响
相比于对照组,刺苦草总生物量和各部分生物量(叶生物量、根生物量、地上生物量和地下生物量)均显著受到水位波动的影响(表1). 与对照组相比,水位波动显著降低了刺苦草的总生物量及各部分生物量(图5). 水位波动幅度能够显著改变刺苦草的叶生物量和地上部分生物量,但波动频率没有显著影响(表2; 图5A,5B). 在同一个水位波动频率(高频率)处理下,生长在低水位波动幅度下的植株的叶生物量(7.8 g ± 0.5 g)和地上生物量(8.6 g ± 0.5 g)显著高于生长在高水位波动幅度下的植株(叶生物量: 5.7 g ± 0.4 g;地上生物量: 6.6 g ± 0.5 g)(图5A,5B). 然而,不同幅度、不同频率的水位波动对总生物量、地下生物量及根生物量的影响不显著(表2; 图5C~E).
3 讨论
3.1 地上部分形态变化
Zhang等[34]对沉水植物响应水位波动的研究发现,水位恒定时穗花狐尾藻 (Myriophyllumspicatum) 的分枝数大于水位波动时,相反,在水位波动时枝条长度明显增加. 顾燕飞等[45]对不同水深条件下苦草 (V.natans) 的形态响应研究表明,随着水深增加,尽管苦草单位面积株数和单位面积匍匐茎数均显著降低,但叶片长度和叶面积随水深显著增加. 同样,本实验发现,相比于对照(无水位波动),波动水位(低幅度低频率、低幅度高频率、高幅度低频率、高幅度高频率)都显著增加了刺苦草的株高和比叶面积,但分株数和叶片数都明显受到水位波动的抑制. 相对于恒定水位,波动水位实际上在一定时间内增加了植株淹水程度. 面对水位变化,耐水淹和弱光的植物一方面可以通过增加株高(茎、叶片或叶柄伸长)投资保障亲株资源(O2、CO2、光照)的利用效率,另一方面通过降低分株密度来降低亲株对子株由于非对称性竞争而产生的负面作用[33,46-48]. 许多研究表明,苦草具有较强的适应低光的能力[49-51]. 在本实验中,当水位在“高水位”和“低水位”之间波动时,一方面,刺苦草通过增加株高、比叶面积,达到接近水面获得更多光资源、提高潜在光合作用能力的目的,另一方面,降低分株密度,保障子株生长发育. 因此,刺苦草可以通过较高的形态可塑性来适应水位波动的胁迫,这种特性有可能促进其种群持续更新. 然而,本实验中水位波动幅度(低幅度和高幅度)、波动频率(低频率和高频率)对刺苦草地上形态参数都不产生影响,其原因可能是由于刺苦草本身作为一种能够忍受淹水和弱光的底层沉水植物,对本实验设置的水深范围有一定的适应能力. 如果恒定水位在1.5 m以上,刺苦草对不同水位波动频率和幅度的响应可能更明显. 这提示我们,探明不同水位波动条件下刺苦草的相关形态特征和生长策略的变化对于沉水植被恢复至关重要.
3.2 地下部分形态变化
凭借根系营固着生存的沉水植物可以从沉积物和水体中吸收养分[52]. 在本实验中,相对于恒定水位,刺苦草根的数量和长度在水位波动下表现为明显降低趋势,而且水位波动幅度对刺苦草根数和根长有显著的抑制作用,表明根系形态对水位波动敏感程度高. 本实验中水位波动处理增加了植物淹水的强度,因此随着水位波动时间的延长,植株对土壤养分利用的能力急剧减弱,根系锚定植物体的能力也会降低,这不利于刺苦草植株的定植. 同样,许多研究表明,随着淹水时间和强度的增加,沉水植物的根数和根长明显下降[37,53]. 对沉水植物而言,水位波动对其生长的影响可能主要是因为光合作用的限制,没有足够的光合产物供给根系,从而抑制了根系的生长,最终导致根系形态参数(根的数量和长度)的下降. 值得注意的是,本实验中区组对株高和根长的影响是显著的,表明刺苦草叶和根形态可能受到水位波动幅度、波动频率以外因素的影响. 由于本实验是在人工实验水池中进行的,今后有必要在自然湖泊条件下进行验证.
刺苦草通过匍匐茎的横向延伸不断产生无性系分株,从而占据更多的生存空间. 因此,匍匐茎是其种群扩展的主要方式,匍匐茎的数量与长度决定了刺苦草种群扩展能力. 在本实验中,相对于恒定水位,水位波动处理增加了植物淹水的强度,因此刺苦草分株数、匍匐茎长度、匍匐茎数明显受到抑制,表明水位波动显著影响刺苦草个体的克隆生长和克隆繁殖,降低种群密度. 然而,本实验中水位波动幅度(低幅度和高幅度)、波动频率(低频率和高频率)对匍匐茎长度和匍匐茎数基本无影响. 表明尽管波动水位与恒定水位相比显著影响了苦草种群扩展能力(即产生匍匐茎,茎顶端发生顶芽分化,产生分株),但本实验设置的不同幅度与频率对其没有显著影响. 这些结果提示我们,适当地降低水位和水位波动速率能有效地提高重建刺苦草群落的成功率.
3.3 生物量积累的变化
生物量是植物生长状态的最主要衡量指标. 相对于恒定水位,水位波动处理下刺苦草总生物量、地上部分和地下部分生物量、叶和根生物量都显著下降. 其原因与水下的光强有关. 一方面,水位波动过程中,“高水位”不仅光强弱,而且植株对CO2、O2和土壤养分的利用能力可能发生改变,从而加剧了植株本身营养物质的消耗. 另一方面,水位波动时的“低水位”可能导致植株在一定时间内暴露在强光下,造成光抑制现象,影响光和营养物质的获取. 必须指出的是,在本实验中,当水位在“高水位”与“低水位”之间波动时,沉水植物调整相关代谢活动也可能需要消耗能量,从而使生长受到限制,导致植株生物量显著降低. Wang等[18]的研究表明,相对于恒定水位(水深90 cm),±30 cm或±60 cm水位波动处理下黑藻生物量显著减小,而金鱼藻的生物量与对照组无明显区别. 因此,水位波动对不同沉水植物的生物量可能产生不同影响. Wang等[33]发现,水位波动频率增加对穿叶眼子菜 (Potamogetonperfoliatus)、脆轮藻和金鱼藻的总生物量没有明显影响. 此外,Yu等[35]对沉水植物响应水位波动的研究中发现,水位波动频率对水车前的总生物量影响不显著,这些研究结果与本实验的发现一致,即刺苦草对不同频率的水位波动适应能力较强. 相反,水位波动幅度增加造成刺苦草地上生物量和叶生物量明显降低. 比较下来,水位波动幅度对刺苦草生物量积累、形态和种群更新影响更大. 在水生态修复工程中,发育良好的沉水植物群落对维持水质、营造健康的生态系统至关重要. 通过本项研究,初步证实对受损沉水植物群落进行重建时,应充分考虑水位的调控范围和目标植物的响应特征. 对刺苦草而言,水位变化速度不超过10 cm/d有助于增强其快速恢复的能力.
4 结论
1)相对于恒定水位,波动水位对刺苦草的总生物量和获取资源相关器官的生物量有显著负面影响. 在水位波动处理下,刺苦草的分株数、叶、根和匍匐茎的生长都明显受到抑制,且刺苦草以显著增加株高和比叶面积来响应水位波动.
2)不同幅度(低幅度和高幅度)、不同频率(低频率和高频率)的水位波动对刺苦草地上部分形态特征基本上没有显著的影响,但叶生物量和地上生物量都受到水位波动幅度显著影响. 不同幅度的水位波动显著改变了根数和根长,而水位波动频率的影响不显著.
致谢:感谢南昌大学的王秀智、张宇、陈腾龙、周瑞雪、武凡、林卓义、吴申浩和陈亚松同学对本实验的帮助. 感谢中国科学院鄱阳湖湖泊湿地观测研究站提供实验场地.
