急尖长苞冷杉叶光合特性及营养元素对海拔的响应
2021-01-12李琬婷程小毛黄晓霞
王 菲 李琬婷 程小毛 黄晓霞
(西南林业大学园林园艺学院,国家林业与草原局西南风景园林工程技术研究中心,云南 昆明 650233)
光合作用是反映植物对环境适应的一个重要指标,而海拔梯度可以反映出环境因子的综合变化,随着海拔梯度的变化,环境因子(光照、温度、水分、土壤等)均呈现出规律性的变化,因此研究植物光合作用对不同海拔的响应对阐明环境因子对植物生长发育的影响有重要价值。有研究表明生活在高海拔地区的植物其光合作用随海拔的增加而减弱,这是由于高海拔的低温环境使得植物叶片控制水分和进行气体交换的能力降低,其蒸腾作用减弱,从而导致光合能力下降[1]。植物叶片营养元素含量不仅能够反映植物的营养水平,还可以反映植物在其自身生长发育过程中的资源利用与分配特征以及对生境的有效利用状况[2-3]。有研究发现植物叶片的氮含量和碳同位素均随海拔高度的增加而变大,温度和大气CO2分压是引起这种现象的主要原因[4-5];但陈立同[6]在对中国西部森林树种的研究中指出青海云杉叶氮含量随海拔梯度的上升呈明显的非线性差异,且在中间海拔3 100 m处达到最小值。上述研究有的是在喀斯特地区自然条件下进行,而有的是在地理环境极为复杂的青藏高原进行,其环境条件相差很大,研究树种也不相同,说明不同植物会采取不同的适应机制以应对随海拔引起的垂直气候变化及土壤变化。
滇西北亚高山地区是典型的横断山区,此地区特殊的地形地貌和复杂多样的气候,使得该地区成为我国生物多样性最丰富、自然环境和生态系统分布极为复杂的地区之一[7]。急尖长苞冷杉(Abies georgeivar.smithii)是我国一种特有的松科冷杉属植物,主要分布区域位于中国云南西北部、四川西南部和西藏东南部。目前为止,关于急尖长苞冷杉的研究区域主要集中在藏东南地区,研究报道多关于其群落结构、物种多样性、养分循环、生长规律以及小气候特征等[8-12]方面。而对于该物种对环境因子的生理生态适应性研究还较少,所以,本次研究以滇西北玉龙雪山保护区内不同海拔梯度的急尖长苞冷杉为对象,对其针叶叶片光合参数及叶营养元素进行测定分析,比较其在不同海拔下针叶的营养元素累积和光合特性的差异,探讨植被营养元素含量特征和光合性质,研究结果对于了解该区域急尖长苞冷杉种群的适应能力、养分的利用、生态系统的养分循环及生物地球化学过程等具有重要的意义。
1 研究地区
玉龙雪山位于云南省丽江市西北部,其地理坐标为27°10′~27°40′ N,100°10′~100°20′ E,是我国纬度最南的一座雪山,也是欧亚大陆最靠近赤道的雪山[13]。其西北临长江石鼓第一湾和金沙江虎跳峡,东麓是海拔约3 km的干海子盆地,南面是丽江盆地。主峰海拔达到5 596 m,海拔梯度极大,形成了暖性、暖湿性、温凉性、北亚热带和寒温性等山地垂直气候带,再加上水文、地形等条件的差异性,导致玉龙雪山植物类群呈现有规律的带状分布。总体上,该区域垂直气候差异显著,随海拔高度的增加,气温下降,湿度增加,降雨量增大[14]。
2 材料与方法
2.1 野外采样
根据急尖长苞冷杉的分布区域,在玉龙雪山保护区内3 100~3 800 m的海拔梯度上每间隔150 m分别设置1个海拔点,总共设置5个取样样地为A1(3 100 m),A2(3 250 m),A3(3 400 m),A4(3 550 m),A5(3 700 m)。在每个样地内,水平距离间隔50 m随机选择5个采样点,在每个采样点内随机选取1株生长基本一致且无病虫害的成年树,树高>10 m,胸径>15 cm,每棵树上均采摘1年生枝上的成熟针叶,统一编号[15]。将编号的针叶实地测定完光合作用后再进行取样,用于叶营养元素及碳同位素组分(δ13C)的测定,采摘后先用液氮冷冻,再放到-80 ℃超低温冰箱保存待用,每项指标的测定至少3次重复。
2.2 指标测定
2.2.1 叶光合参数的测定
在设置好的每个海拔梯度生长地内分别选择3株长势良好且具有代表性的急尖长苞冷杉植株,取5枝生长一致、无病害的针叶,然后用LI-6400光合作用仪测定其光合作用,测定的光合参数包括胞间CO2浓度(Ci)、瞬时最大净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(E),测定时采用6400-05簇状叶室,叶室内温度为23~28 ℃,相对湿度为36%~55%,设置CO2浓度为400 μmol/mol,光强为1 600 μmol/(m2·s)[16]。急尖长苞冷杉叶光合参数的测定在2014年9—10月完成,所有的指标测定都选择在晴朗天气的早上9:00—11:00完成,每个数据3个重复,每个重复测定3~5次,取平均值。
2.2.2 叶营养元素的测定
1)叶元素含量测定:先用自来水冲洗植物样品叶片2次以上,去掉灰尘及泥沙等杂物,然后用去离子水冲洗3次。于105 ℃下杀青10 min后在65 ℃下烘干至恒质量。采用高速万能粉碎机(天津市泰斯特仪器有限公司)将样品粉碎磨成粉末状。用C、N元素分析仪(Vario macro,elementar,Germany)测量样品中的C、N元素含量;P元素的测量则先将样品加入浓HNO3和H2O2,再在微波消解仪(MARS Xpress,CEM,American)中进行微波消解,最后用ICP-OES(ICP-OES 7300DV,PerkinElmer,American)测量样品P元素含量[17]。
2)叶面积及叶干质量测定:把采摘好的植物针叶整齐平整地黏到透明胶上,用Epson perfection V700 photo扫描仪扫描针叶后,通过Leaf Auto Compute软件计算取其平均值,再将叶烘干,称取干质量[18]。最后分别计算单位叶面积的C、N、P含量和单位叶质量的C、N、P含量。
3)叶碳同位素组分(δ13C)测定:根据文献[19],测定不同海拔生长的急尖长苞冷杉针叶样本的δ13C。具体步骤如下:先将植物叶片洗净后置于70 ℃恒温48 h以上烘干至恒质量,粉碎样品,过200目筛后制成供试样品。再将预处理过的样品7~8 mg封入真空的燃烧管,并加入催化剂和氧化剂,550 ℃燃烧1 h。冷却后收集纯化的CO2,然后用质谱仪测定样品碳同位素比率,以PDB(Pee Dee Belemnite)为标准,根据下面公式计算:δ13C ={[(13C/12C)sample-(13C/12C)standard] /(13C/12C)standard}×1 000‰。
4)光合氮利用效率(PNUE)和长期氮利用效率(NUE)测定:NUE用叶总碳含量与总氮含量之比得出。PNUE用叶最大净光合与叶总氮含量之比得出[20]。
2.3 数据分析
采用SPSS分析软件和Excel表格进行数据的计算、统计分析和图表绘制,采用Duncan法进行不同数据组间的差异分析。
3 结果与分析
3.1 不同海拔急尖长苞冷杉针叶的光合特性
从图1可知,急尖长苞冷杉针叶光合参数都受到海拔的显著影响:瞬时最大净光合速率在海拔3 250 m及3 550 m点高于其他海拔点,而在最高海拔点3 700 m时降至最低;气孔导度、蒸腾速率以及胞间二氧化碳浓度均随海拔的增加先升高后下降,且在3 250 m增至最大,而在3 700 m海拔点这几个参数值均处于最低水平。

图1 急尖长苞冷杉叶光合参数随海拔梯度的变化Fig.1 Photosynthetic parameters of A.georgei var. smithii needles at different altitudes
3.2 不同海拔急尖长苞冷杉针叶的营养元素差异性
由表1可知,急尖长苞冷杉叶营养元素碳、氮、磷的含量都受到海拔的显著影响。随海拔的升高,单位叶面积碳、氮、磷含量出现非线性变化,碳含量在3 250 m点降至最低,而在3 400 m及3 700 m点处于显著增加水平(P<0.05);而单位叶面积的氮含量在海拔3 550 m点最低,而在海拔3 400 m点增至最大;单位叶面积磷含量在海拔3 550 m点处于最低水平,而在海拔3 700 m点处于最高水平。单位叶干质量的碳、氮、磷的含量均在海拔3 400 m点处于显著增加水平,而在海拔3 550 m点处于显著降低水平(P<0.05)。
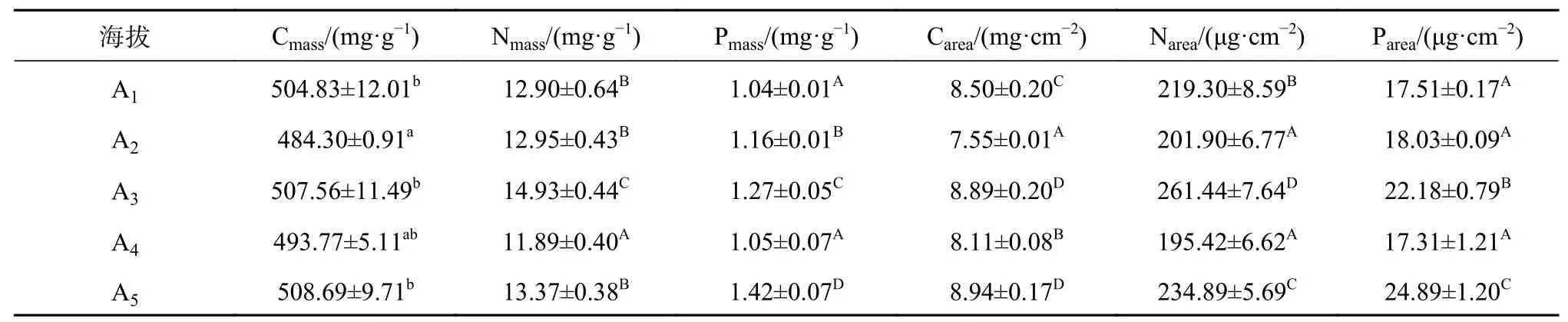
表1 急尖长苞冷杉针叶营养元素含量随海拔梯度的变化Table 1 Content of nutrient elements of A.georgei var. smithii needles at different altitudes
3.3 不同海拔急尖长苞冷杉针叶δ13C差异性
从图2中δ13C的变化可以看出,随海拔增加,急尖长苞冷杉的长期水分利用效率先下降后增加,在海拔3 250 m处其值最低,然后再逐渐增加,在海拔3 700 m时达到最大值,且在其他海拔间均差异显著。δ13C的变化可以代表植物长期用水效率,急尖长苞冷杉长期水分利用效率在高海拔处的显著增加,也是其对高海拔环境的一种适应。
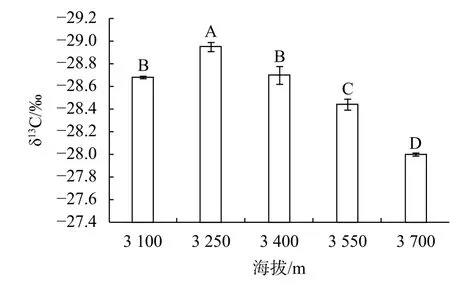
图2 急尖长苞冷杉碳同位素组组分(δ13C)随海拔梯度的变化Fig.2 Content of δ13C of A.georgei var. smithii needles at different altitudes
3.4 不同海拔急尖长苞冷杉叶NUE和PNUE差异性
从表2可知,长期氮利用效率NUE及光合氮利用效率PNUE都受到海拔梯度的显著影响,随海拔增加呈现降低-增加-再降低的趋势,且在海拔3 400 m点降至最低,而在3 550 m点却处于最高水平。
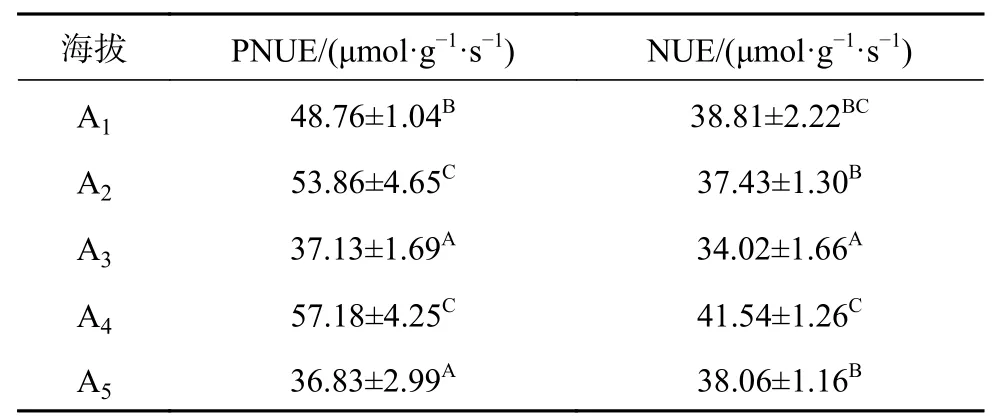
表2 急尖长苞冷杉针叶叶片中PNUE和NUE含量随海拔梯度的变化Table 2 Content of PNUE and NUE of A.georgei var. smithii needles at different altitudes
4 结论与讨论
随着海拔梯度的不断上升,环境条件会逐渐变得恶劣,植物的生长发育逐渐受到水分、温度及其他生态因子的限制,导致其适应性发生显著的变化[15],植物会通过调整内在的生理生化及外在的形态结构等来应对环境的变化。叶作为植物与环境直接接触的器官,其光合特性可以体现出植物对环境变化的适应性[21]。有研究发现不同植物的光合生理差异性是自身生理因素和不同海拔下的环境因素共同作用的结果[22]。金高明[23]对富士苹果的研究发现其净光合速率、蒸腾速率和气孔导度均随着海拔的升高而逐渐增大,胞间CO2浓度随着海拔升高表现为先增大后减小再增大的趋势。韦玉等[24]对不同海拔梯度矮嵩草(Kobresia humilis)的光合响应差异进行研究时,结果显示,随着海拔的不断升高,矮嵩草的最大净光合速率和光合能力均逐渐增大。但何涛等[25]的研究指出随海拔梯度从2 300 m升高至3 800 m,火绒草(Leontopodium leontopodioides)叶片的最大净光合速率呈下降趋势。火绒草在光合特性上的变化是对逆境的一种适应,是青藏高原特殊生态条件长期胁迫的结果。有关专家对意大利一些植物叶片的研究发现,海拔升高时,叶片的气孔导度减小[26]。本研究中,随着海拔的升高,急尖长苞冷杉的气孔导度、蒸腾速率以及胞间CO2浓度总体上呈现先增后降的变化趋势,且在海拔3 250 m处最大,而净光合速率在3 250 m海拔点及3 550 m海拔点无显著区别,且均大于其他各海拔。说明急尖长苞冷杉的光合碳同化在这两个海拔点更强,其光合能力较强,可以制造更多的碳水化合物,供给自身生长发育。此外,从其他几个光合参数的变化还可以看出3 250 m海拔点净光合速率的上升更多受气孔的调控,而在3 550 m海拔点净光合速率的上升更多受到非气孔因素的影响;而在高海拔3 700 m处,净光合速率、气孔导度、蒸腾速率以及胞间CO2浓度都降至最低,产生这一变化的原因可能是由于在高海拔区域,低温限制了植物的生长,且光照强度的增加,大气、水分状况的改变,使得植物处于不良生境条件下,当蒸腾速率过大,为防止植株自身针叶水分过度蒸发,植株通过调控针叶表面的气孔开闭合状态,使得叶气孔开度降低,胞间二氧化碳浓度也随之降低,光合能力也受到一定影响,从而导致光合速率下降。
叶营养元素是植物生长发育的物质基础,能反映植物营养元素吸收水平。祁建等[27]对辽东栎(Quercus liaotungensis)特性变化的研究也发现,叶氮、磷的含量随着海拔的升高而增大。而吴尚明等[28]对珙桐(Davidia involucrata)的研究时发现其叶片碳氮代谢并未随海拔的升高而显著改变,其研究地点的海拔范围为1 400~1 800 m,分布区海拔梯度范围较窄。叶氮、磷含量沿海拔梯度的升高说明叶的水分和能量代谢及抗性也在增强,这也体现了植物为了适应高海拔地区恶劣的生存环境,对高海拔地区的一种响应机制。本研究对象急尖长苞冷杉的海拔分布范围在3 100~3 800 m,叶碳、氮含量受海拔的影响显著。急尖长苞冷杉叶单位叶干质量的C、N、P的含量在海拔3 400 m点处于显著增加水平,而当海拔升高到3 700 m时,单位叶面积的C、N、P含量处于显著增加水平这也间接说明该树种对海拔的响应积极。植物体内75%的氮都集中于叶绿体中,且大部分都用于光合器官的构建,因此氮素是光合物质代谢和植物生长的关键性因子[29]。有研究表明高海拔处土壤有机碳和全氮含量明显高于低海拔地区,且土壤全磷含量随海拔的升高呈现上升趋势[30]。李长燕等[31]研究发现随着海拔梯度的增加土壤中有机碳、全氮、全磷含量也相应增加,为珙桐群落的正常生长发育提供必要的营养。此外,低温、高光合能力以及含氮量丰富的土壤也能促使植株积累较高的氮素含量。与以上研究结果相似,本实验中急尖长苞冷杉单位叶面积的C、N、P含量在3 700 m处显著增加,可能与土壤中碳氮磷含量的变化密切相关。
植物叶的δ13C值与植物长期的水分利用效率和自身叶的气孔导度有密切的关系,由于随着海拔升高,气温下降和CO2浓度降低,导致CO2扩散能力降低,从而使叶气孔导度下降,使得叶的δ13C分馏能力降低,δ13C值增大[32]。有研究表明,随着海拔的升高温度逐渐下降,植物可用的土壤中的有效水降低,造成高山植物生理干旱,从而使叶气孔导度降低,δ13C值增大[33],这也间接说明了用叶δ13C值来表征植物水分利用效率的可靠性。本研究结果表明,急尖长苞冷杉叶δ13C值随着海拔梯度升高而呈现先下降后增加的变化趋势,这与冯秋红等[34]对异型柳(Salix dissa)叶片δ13C值的研究结果类似。但林玲等[35]在研究急尖长苞冷杉与川滇高山栎(Quercus aquifolioides)叶δ13C值沿海拔梯度的变化后发现其δ13C值与海拔梯度之间并不是一个线性增加的关系,而是不规则的变化关系。这可能是水分、温度以及降水等因子在不同树种之间的共同作用导致的结果。研究结果显示急尖长苞冷杉叶δ13C的值在海拔3 250 m最小,这可能是由于该海拔条件下水分较充足,使得比叶面积,气孔导度及蒸腾速率都处于最大值,因此其长期水分利用效率最低。而随着海拔的逐渐增加,海拔越高,δ13C值越大,且差异显著,说明长苞冷杉随海拔的增加,长期水分利用效率也增加,这也可反映出急尖长苞冷杉对高海拔环境的一种适应。叶光合氮利用效率可以较叶氮含量更好的诠释光合作用与氮素之间的关系,从而揭示不同物种间光合能力差异的原因[21]。研究叶光合氮利用效率的特征以及光合氮利用效率与其他叶性状之间的联系,对深入探讨叶氮在不同植物体内的重要性有很大的好处。本研究发现急尖长苞冷杉长期氮利用效率C/N(NUE)和光合氮利用效率PNUE均受到海拔梯度的显著影响,在海拔3 550 m点均处于最高水平,而在海拔3 400 m及3 700 m点都显著降低,且在海拔3 400 m处于最低水平,说明在海拔3 550 m该树种有着更高的氮利用效率,这主要归因于碳含量的增加和氮含量的减少,在海拔3 550 m,光合速率最大,在叶中累积了更多的碳水化合物,同时也能够引起一系列代谢水平的变化。长期氮利用效率及光合氮利用效率在海拔3 700 m点都显著降低,这可能是由于受到高海拔的极端环境如低温、干旱、贫瘠土壤及强UV-辐射等的影响,使得气孔开度减小,胞间二氮化碳浓度减小或光合器官受损,使得植株光合能力减弱。张石宝等[36]对帽斗栎(Quercus guyavaefolia)的研究发现其水分利用效率和氮利用效率在海拔中部比低海拔或高海拔处的都要高,这和本研究得出的结果类似。
综上所述,滇西北玉龙雪山急尖长苞冷杉天然种群的光合生理特性及叶营养元素累积对海拔变化有积极响应,也说明该物种能较好的适应亚高山环境。3 550 m以下海拔环境对其生长无限制影响,其净光合速率及叶营养元素水平、氮利用效率等均处于中间水平;而在海拔上升至3 550 m时,急尖长苞冷杉叶最大净光合速率、光合氮利用效率及长期氮利用效率都处于最高水平,说明海拔3 550 m是急尖长苞冷杉的最适生长区;而当海拔进一步上升到3 700 m时,单位叶面积的C、N、P含量及长期水分利用效率δ13C值都处于最高水平,以应对高海拔的逆境胁迫,但同时净光合速率、气孔导度、蒸腾速率、光合氮利用效率、长期氮利用效率等都降至最低水平,说明3 700 m海拔区域由于环境条件愈发严厉,限制了急尖长苞冷杉的生长发育,同时可促使其调节营养代谢分配及提高其用水效率等来应对高海拔的恶劣环境,也体现了急尖长苞冷杉对高海拔地区生存环境的适应,以延长和保存种群生命。
