不同磷水平外源独脚金内酯和独脚金内酯抑制剂对油菜根系形态和产量的影响
2021-01-04唐长生王煜潇丁广大徐芳森
唐长生,王煜潇,丁广大,徐芳森,石 磊
(华中农业大学作物遗传改良国家重点实验室,农业农村部长江中下游耕地保育重点实验室/微量元素研究中心,湖北 武汉 430070)
油菜是我国主要的油料作物,种植面积和总产量均约占世界三分之一。菜籽油是国产食用植物油的第一大来源,年均产量1 300 多万t,在我国食用油市场中具有举足轻重的地位[1]。推动油菜产业发展是我国植物油供给安全的重要保障。
磷是植物生长发育所必需的第二大矿质营养元素,它是核酸、磷脂、植素、ATP 等多种化合物的重要组成部分,积极参与植物体内的生化合成、能量转移、信号转导等代谢过程,同时提高作物的抗逆性和适应能力,在产量和品质形成过程中起着重要的作用[2]。油菜是早期磷营养类型作物,早期缺磷所受的损害,即使后期有充足的磷营养,也无法补救。适量施用磷肥能大幅度增加油菜分枝数、每株角果数和每角果籽粒数,并在一定程度上提高籽粒千粒重,为高产稳产打下基础[3]。
独脚金内酯(Strigolactones)是近年来发现的一种植物激素或其前体,它是一类天然的独脚金醇化合物和人工合成类似物的总称,它存在于所有植物中,在根中合成,一部分向地上部运输以调节植物的生长,另一部分直接释放到土壤中以介导植物与土壤微生物及寄生植物间的信号交换[4-5]。独脚金内酯作为一类抑制植物分枝发育的植物激素,可与生长素和细胞分裂素协同调控植物的分枝生长[6-9]。
独脚金内酯的合成除了受遗传调控外,还受环境条件比如土壤营养状况的调控[4]。与正常磷处理相比,低磷处理的豆科植物红苜蓿、非豆科植物西红棉及水稻独脚金内酯的合成分泌显著增加,当转入正常磷的营养液后独脚金内酯的合成分泌迅速下降[10-14]。独脚金内酯缺乏能诱导拟南芥侧根的形成[15]。拟南芥施用独脚金内酯后根毛伸长,花青素积累,酸性磷酸酶活性增强,干重降低,表现出典型的缺磷症状[16]。低磷和低氮胁迫条件水稻野生型和独脚金内酯合成缺陷突变体(D10、D27和D3)种子根长均增加,根系密度均减少,正常磷和正常氮处理外源施用GR24 抑制了野生型和独脚金内酯合成缺陷突变体根系密度,症状与低磷、低氮胁迫相似,说明独脚金内酯参与水稻根系调控[17-18]。此外,氮、磷缺乏和外源施用GR24 都减少了水稻生长素在野生型D10和D27突变体中的运输[18]。
油菜缺磷苗期老叶出现紫红色,根系生长受阻,成熟期分枝数显著减少,可能与低磷时独脚金内酯合成增加和向地上部的分配增强有关。本课题拟通过营养液培养和全生育期根箱土培试验,研究外源独脚金内酯及其抑制剂对不同磷条件下油菜根系形态和地上部生长的影响,研究结果可为磷肥减施增效条件下通过独脚金内酯及其抑制剂调控油菜根系形态、地上部分枝,提高作物产量提供理论依据。
1 材料与方法
1.1 试验材料
供试油菜品种为长江中下游主推品种“中双11 号”,该品种由中国农业科学院油料作物研究所选育,为半冬性常规甘蓝型油菜品种。试剂为GR24(人工合成的独脚金内酯)和TIS108(人工合成的独脚金内酯抑制剂)。
1.2 试验方法
1.2.1 营养液培养试验
进行营养液培养的聚乙烯塑料盆规格为35 cm×22 cm×5 cm(长×宽×高),上方为打有12 孔的黑色KT 幼苗定植板,盆体外壁四周用黑油漆涂黑以避光。播种前,先用去离子水将种子浸泡过夜,然后将纱布覆在育苗盆上并用枪头和细绳固定边缘,装满去离子水后将种子均匀播在纱布上,待发芽后子叶完全展开、主根长度4 ~6 cm 时(约6 d),选取长势较一致的幼苗用海绵轻轻裹住幼苗下胚轴(避免夹到根部)将其根部通过黑色KT 板上的圆孔放入盛装3 L 营养液的塑料盆中。试验采用霍格兰大量营养元素营养液和阿农微量元素营养液[19],具体配方如下,大量元素(g/L):KNO30.51,KH2PO40.14,MgSO4·7H2O 0.49,Ca(NO3)2·4H2O 1.18,K2SO40.09; 微 量 元 素(mg/L): 铁盐(EDTA-Fe)32.5,H3BO32.84,CuSO4·5H2O 0.08;ZnSO4·7H2O 0.22,MnCl2·4H2O 1.81,Na2MoO4·2H2O 0.09。初始营养液的浓度为全营养液浓度的1/4,每5 d 更换一次,浓度依次为1/2 全营养液、全营养液。GR24 和TIS108 浓度梯度试验均在正常磷条件下进行,试验设置5 个浓度处理,分别为0、0.01、0.1、1 和5 μmol/L,每个处理3个重复,每个重复12 株,采取完全随机设计。分别在培养8、15 和22 d 取样,测定根系形态性状。通过上述筛选试验确定了适合本研究的GR24 和TIS108 的浓度分别为5 和0.1 μmol/L,再设置低磷(5 μmol/L)、低磷添加GR24、低磷添加TIS108、正常磷、正常磷(250 μmol/L)添加GR24、正常磷添加TIS108 共6 个处理,每个处理3 个重复,每个重复共12 株油菜,采取完全随机设计。KH2PO4为试验用磷的来源,低磷不同梯度处理添加一定量的K2SO4以保证各处理钾肥量相同。每次更换营养液时均根据各处理要求添加GR24 或TIS108。
试验在光照培养室中进行,光照培养室光照强度为300 ~320 μmol/(m2·s),空气相对湿度为65%~80%,光照周期为16 h/8 h(光照/黑暗),温度22℃。
1.2.2 根箱土培试验
采用自制根箱进行全生育期土培。试验用土壤为华中农业大学校内实验实习基地的黄棕壤,其基本理化性状如下:pH 值6.84(1∶5),有机质为5.2 g/kg,全氮为0.25 g/kg,有效磷(Olsen-P)为2.2 mg/kg。根箱规格为67 cm×18 cm×98.5 cm(长×宽×高)[20],根箱下层78.5 cm(从底部往上)填装无施肥处理的土壤,根箱上部20 cm 填装30 kg 与肥料混匀的土。根层土施肥量按每千克土施N 0.2 g、P2O50.15 g(正常磷)/0.005 g(低磷)、K2O 0.20 g,以(NH4)2SO4、KH2PO4和KCl 为 肥源。并按每千克土施1 mL Anon 微量元素营养液和EDTA-Fe 5 mL(浓度为0.05 mmol/L)(以上试剂均为化学纯)。2014 年10 月24 日播种,播种后覆盖薄膜2 d,种子萌发约1 ~2 cm 后揭开薄膜。两周后间苗,每个根箱按相同株距留苗9 株,油菜生长期以去离子水浇灌,5 周后定苗3 株。试验设低磷、低磷喷施GR24、低磷喷施TIS108 和正常磷4 个处理,每个处理3 个根箱,每个根箱3 株油菜。分别在五叶期(35 d)、抽薹期(140 d)和成熟期(210 d)取样考察相应指标。GR24 浓度为5 μmol/L,TIS108 浓度为0.1 μmol/L,五叶期取样后开始进行喷施处理,开第一朵花后停止喷施,期间每周喷施一次,以叶面布满雾滴为度。
1.3 相关指标测定
1.3.1 根系形态
营养液培养油菜根系形态:油菜从营养液中取出,用剪刀将根系和地上部剪断分开,用蒸馏水将油菜根系洗干净,用直尺直接测量主根长度,再将根系放入盛有蒸馏水的透明浅盘中,用镊子小心把根系展开,将浅盘置于根系扫描仪上,扫描根系,图像存入计算机,用根系图像分析软件(WinRhizo Pro,Régent Instruments,Canada)对根系形态进行分析。
根箱土培油菜根系形态:五叶期将油菜根系从土壤中取出,用直尺直接测量主根长度,然后用根系扫描仪进行扫描。抽薹期和成熟期根系很大,取样时尽可能保持根系完整,然后按一级侧根分开分别用根系扫描仪进行扫描,尽量减少根系重叠,获取图片,用WinRhizo 软件分析根系形态,最后得出主根长、总根长、根系总表面积、根系总体积、根尖数等指标。
1.3.2 干重和磷含量
不同生育期地上部和根系样品,105℃杀青30 min,80℃烘干至恒重,称量地上部干重和根系干重,计算根冠比,之后用植物粉碎机磨碎测定植物全磷含量。把磨碎的样品(约为0.1 g)转入50 mL消化管,H2SO4−H2O2消煮后采用钼锑抗比色法、流动注射分析仪(FIAstar5000 analyzar,FOSS,丹麦)测定磷浓度[21]。
1.3.3 根系和地上部生长素含量
营养液培养21 d 时进行油菜根系和地上部取样。取0.5 g 以上样品放入1.5 mL 离心管,液氮冷冻,-70℃超低温冰箱保存至激素测定。激素测定时,样品用液氮研磨、摇床脱色、加buffer 液处理,然后放置于质谱专用上样瓶内,采用超快速液相色谱法进行测定[22]。
1.3.4 成熟期油菜分枝数、株高、产量
成熟期测定不同处理每株油菜株高和分枝数,油菜种子收获后晒干(含水量不高于10%)称取种子产量。
1.4 数据统计
采用Excel 2010 软件计算平均数和标准误;采用SPSS 18.0 进行单因素方差分析和差异显著性分析。
2 结果与分析
2.1 GR24 和TIS108 对油菜苗期根系形态和地上部生长的影响
2.1.1 正常磷不同浓度外源GR24 和TIS108 对油菜苗期根系形态的影响
正常磷(NP)以及NP 添加不同浓度GR24 处理油菜主根长、总根长、根系总表面积、根系总体积和根尖数均表现为GR24 浓度越大,对根系的抑制效果越强,其中8 d 时NP 处理主根长、总根长、根系总表面积、根系总体积和根尖数均显著长(大)于NP+1 μmol/L GR24 和NP+5 μmol/L GR24处理;15 d 时NP 处理总根长、根系总表面积、根系总体积显著高(大)于NP+0.1 μmol/L GR24,主根长、总根长、根系总表面积、根系总体积和根尖数显著高(大)于NP+1 μmol/L GR24 和NP+5 μmol/L GR24 处理;22 d 时,NP 处理主根长、总根长、根系总表面积、根系总体积、根尖数显著高(大)于NP+1 μmol/L GR24 和NP+5 μmol/L GR24处理(表1)。
NP 以及NP 添加不同浓度TIS108 处理油菜主根长、总根长、根系总表面积、根系总体积和根尖数均表现为低浓度TIS108 促进根系生长,高浓度TIS108 抑制根系生长,其中8 和15 d 时 0.01 μmol/L TIS108 促进作用最强,22 d 时0.1 μmol/L TIS108对根系的促进作用大于0.01 μmol/L TIS108。8 d时NP+0.01 μmol/L TIS108 处理根系总表面积、根系总体积和根尖数显著高于NP,而NP+5 μmol/L TIS108 处理主根长、总根长、根系总表面积、根系总体积和根尖数均显著低于NP 处理。15 d 时NP+0.1 和0.01 μmol/L TIS108 处理根尖数均显著高于NP 处理,但主根长、根系总表面积、根系总体积无显著差异;NP+ 1 和5 μmol/L TIS108 处理总根长、根系总表面积和根系总体积均显著低于NP 处理。22 d时NP+0. 1 μmol/L TIS108 处理主根长、总根长、根系总表面积、根系总体积和根尖数均显著高于NP 处理,而NP+5 μmol/L TIS108 处理上述根系参数除根系总体积外均显著低于NP 处理(表2)。
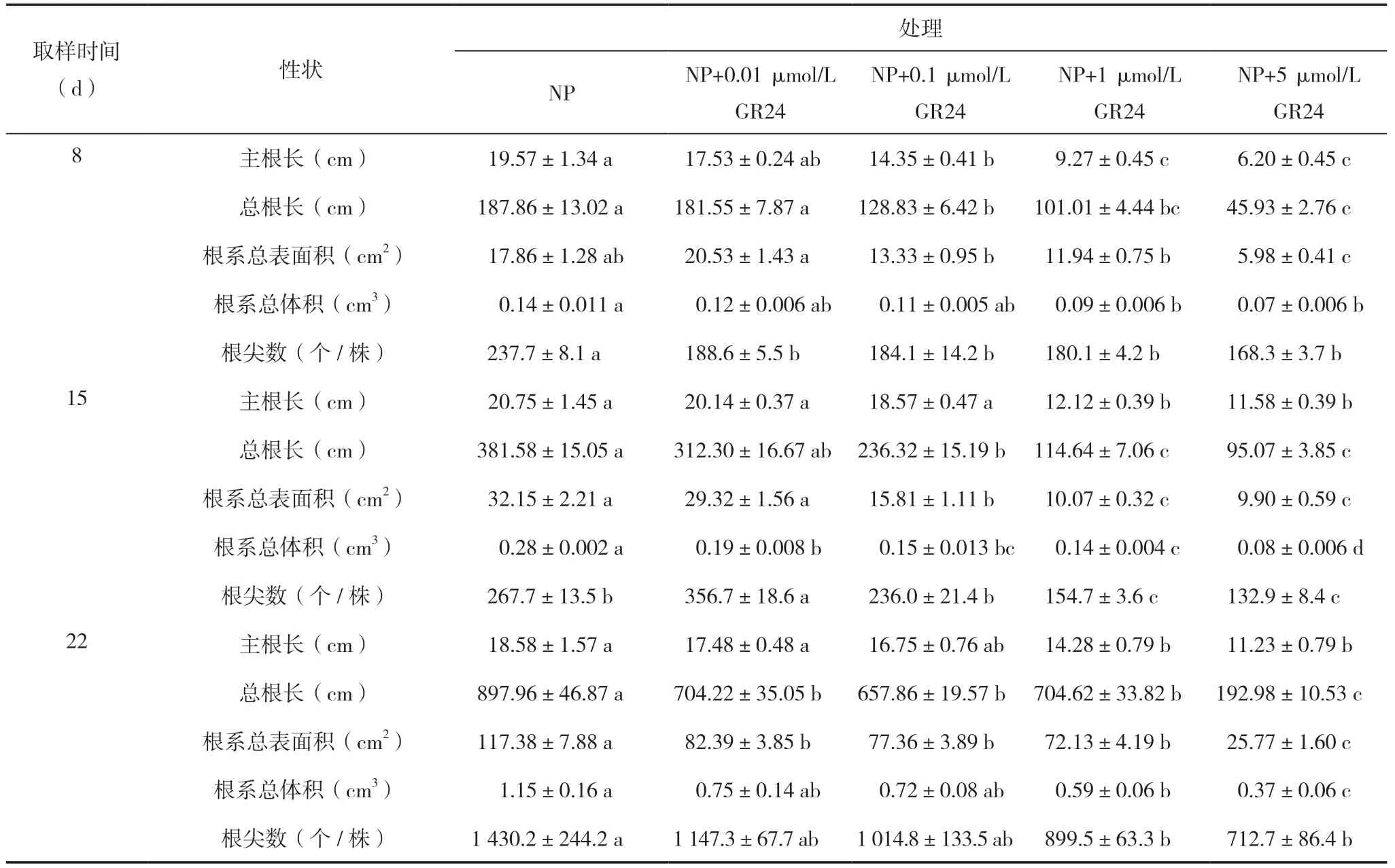
表1 不同浓度GR24 处理中双11 号的主根长、总根长、根系总表面积、根系总体积、根尖数
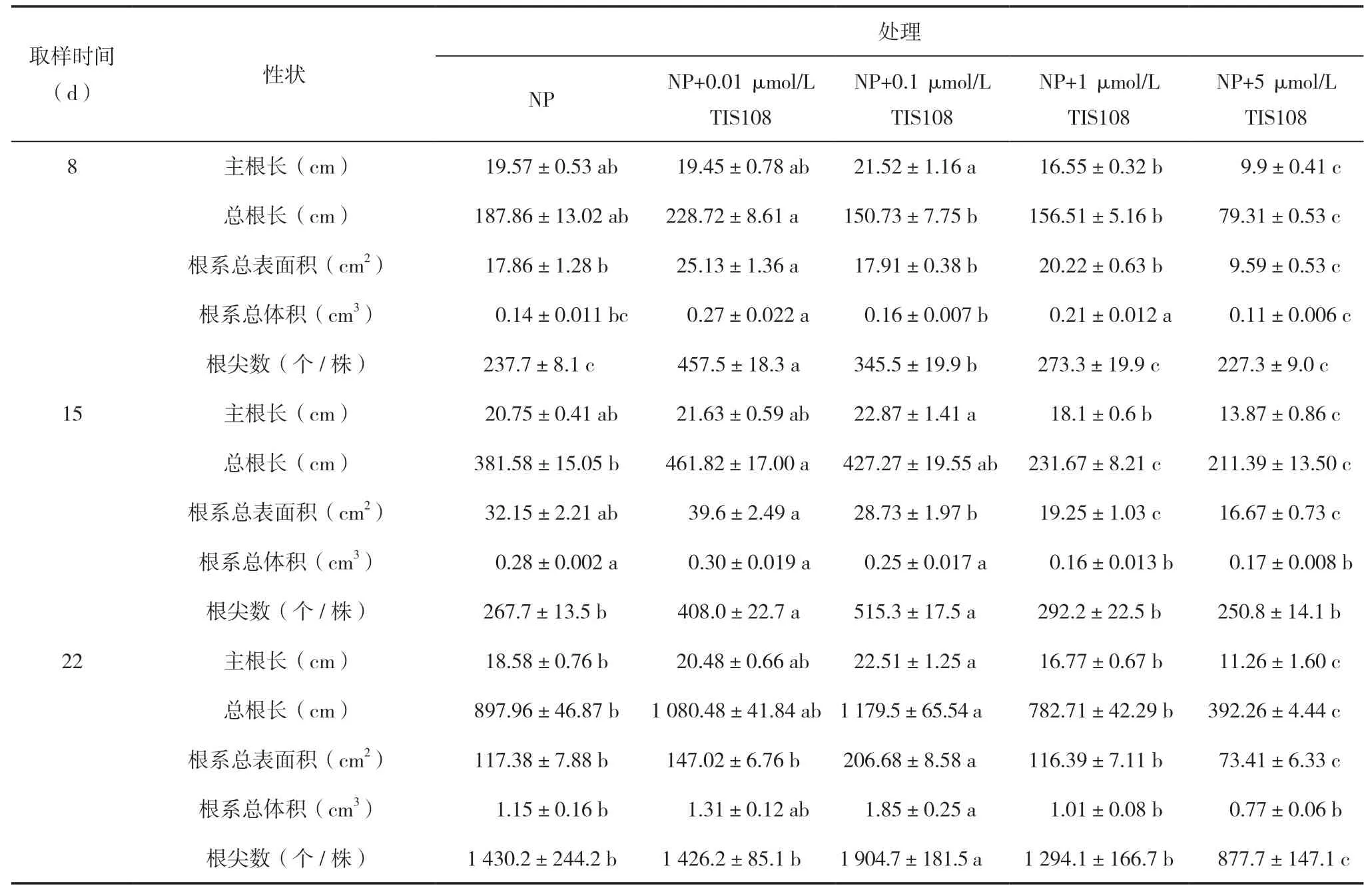
表2 不同浓度TIS108 处理中双11 号的主根长、总根长、根系总表面积、根系总体积、根尖数
根据苗期不同浓度GR24 和TIS108 处理油菜根系和地上部生长的差异,确定后续试验中采用的GR24 浓度为5 μmol/L,TIS108 浓度为0.1 μmol/L。
2.1.2 不同磷水平外源GR24 和TIS108 对油菜根系形态的影响
7 d 时NP 处理油菜根系总体积和根尖数均显著大(多)于LP 处理。两个磷水平,TIS108 均表现为促进根系生长,GR24 均表现为抑制根系生长。其中,NP 处理施用TIS108 主根长、总根长、根系总表面积、根系总体积和根尖数均显著高于GR24;LP 处理施用TIS108 主根长、总根长、根系总表面积和根系总体积均显著高于GR24(表3)。
14 d 时NP 处理油菜总根长、根系总表面积和根系总体积均显著长(大)于LP 处理。两个磷水平施用TIS108 均表现为促进根系生长,施用GR24 均表现为抑制根系生长。其中,NP 处理施用TIS108 总根长、根系总表面积、根系总体积、根尖数显著长(大、多)于施用GR24;LP 处理施用TIS108 主根长、总根长和根系总体积显著长(大、多)于施用GR24(表3)。
21 d 时NP 处理油菜总根长、根系总表面积、根系总体积和根尖数均显著长(大)于LP 处理。不同磷水平施用TIS108 均显著促进了上述根系指标,而施用GR24 均显著抑制了上述根系指标(除根尖数)(表3)。NP 处理,施用TIS108 和GR24主根长均表现为抑制,但GR24 的抑制作用更强;而LP 处理,施用TIS108 促进了主根伸长,施用GR24 抑制了主根伸长(表3)。
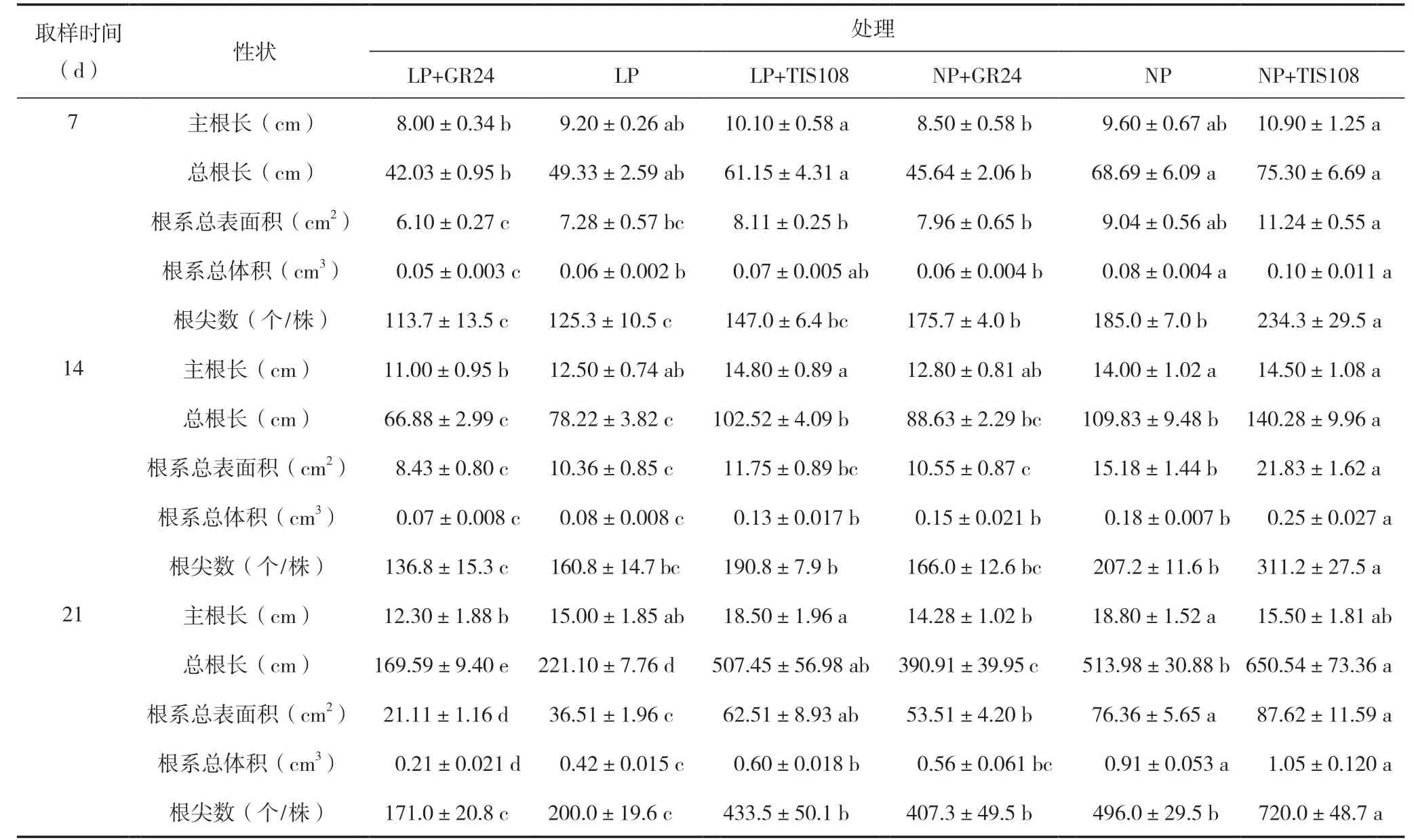
表3 低磷和正常磷GR24 和TIS108 处理中双11 号主根长、总根长、根系总表面积、根系总体积、根尖数
2.1.2 不同磷水平外源GR24 和TIS108 处理油菜根系和地上部干重
7、14 和21 d,油菜NP+TIS108 和NP 处理地上部干重均显著大于LP 和LP+GR24 处理,根冠比则相反(表4)。两个磷水平,TIS108 均促进了油菜地上部和根的生长,而GR24 则表现相反。其中,7 d 时LP 3 个处理地上部干重:LP+TIS108 显著大于LP,LP 显著大于LP+GR24;NP 3 个处理中NP+TIS108显著大于NP 和NP+GR24;两个磷水平TIS108 处理根干重均显著大于GR24 处理。14 和21 d 时,LP 3 个处理中LP+TIS108 地上部干重显著大于LP和LP+GR24,NP+TIS108 和NP 处理地上部干重显著大于LP(表4);两个磷水平TIS108 处理根干重均最高,GR24 处理根干重均最低(表4)。7、14和21 d,LP+GR24 处理根冠比均大于NP+GR24,LP 处理根冠比均大于NP;7 和14 d,LP+TIS108处理根冠比均大于NP+TIS108。但21 d 两个处理根冠比无显著差异(表4)。
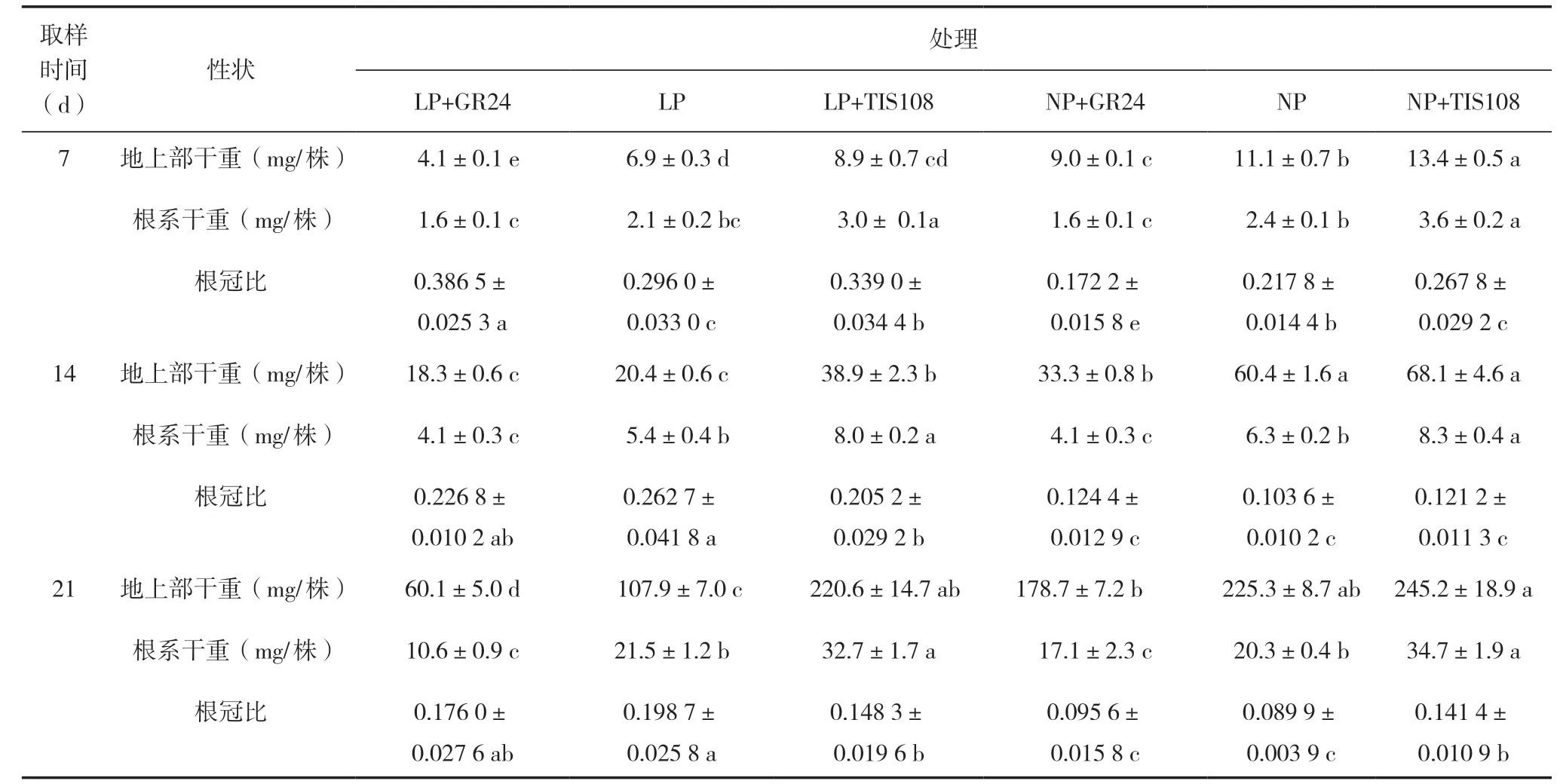
表4 低磷、正常磷GR24 和TIS108 处理中双11 号地上部和根系干重、根冠比
2.1.3 不同磷水平外源GR24 和TIS108 处理油菜根和地上部磷含量
两个磷水平,油菜地上部和根系磷含量均表现为TIS108 处理最高,GR24 处理最低,施用GR24显著降低了地上部和根系磷含量,而施用TIS108显著提高了地上部和根系磷含量(图1)。
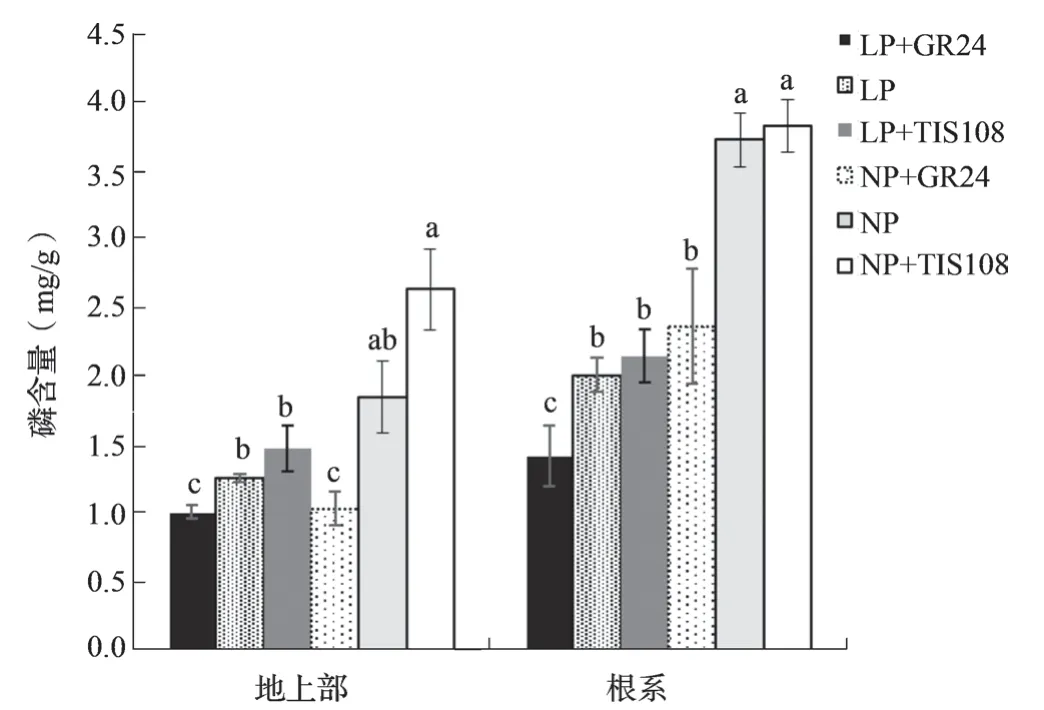
图1 低磷和正常磷GR24 和TIS108 处理中双11 号根系和地上部磷含量(21 d)
2.1.4 不同磷水平外源GR24 和TIS108 处理油菜地上部和根系生长素含量
与NP 处理比较,LP 处理油菜根系生长素浓度显著降低,但两个磷处理地上部生长素浓度没有显著差异(图2)。低磷时,TIS108 处理地上部生长素浓度显著低于LP 和LP+GR24 处理;正常磷时,TIS108 处理和GR24 处理地上部生长素浓度均显著降低,其中TIS108 处理降低幅度显著大于GR24 处理(图2 A)。低磷时,TIS108 处理根系生长素浓度显著增加,GR24 处理根系生长素浓度显著降低;正常磷时,GR24 处理根系生长素浓度显著降低,但NP 和NP+TIS108 处理生长素浓度无显著差异(图2 B)。
2.2 低磷胁迫喷施GR24 和TIS108 对油菜全生育期根系和地上部生长的影响
2.2.1 低磷胁迫喷施GR24 和TIS108 油菜根系形态的差异
抽薹期和成熟期NP 处理油菜主根长、总根长、根系总表面积、根系总体积和根尖数均显著大(长、多)于LP+GR24、LP 和LP+TIS108 处理(表5)。其中,抽薹期LP+TIS108 处理总根长和根系总表面积显著大于LP;LP+TIS108 处理主根长、总根长、根系总表面积、根系总体积和根尖数均显著大(长、多)于LP+GR24;LP 处理主根长、总根长和根尖数均显著大(长、多)于LP+GR24(表5)。成熟期LP+TIS108 处理主根长、根系总表面积、根系总体积和根尖数均显著大于LP+GR24 处理;LP处理主根长和根尖数均显著大于LP+GR24 处理(表5)。
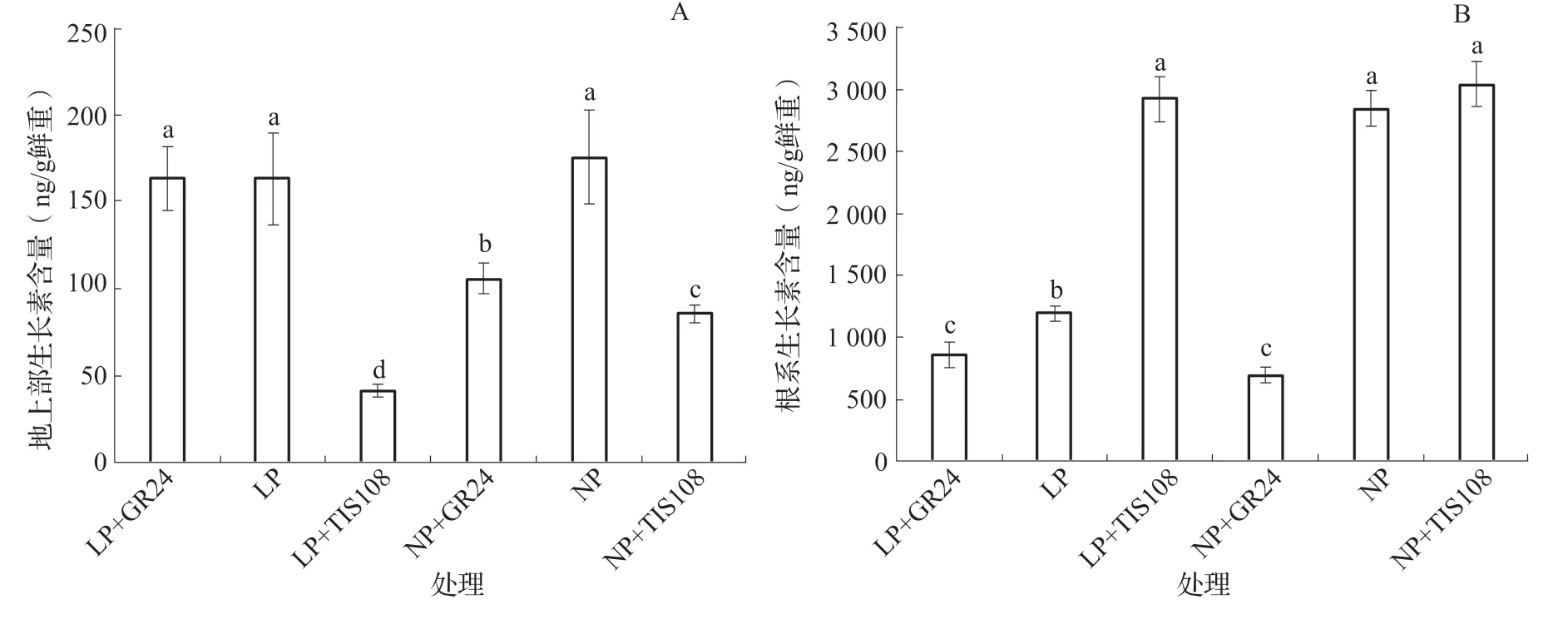
图2 低磷和正常磷水平GR24 和TIS108 处理中双11 号地上部和根系生长素含量(21 d)
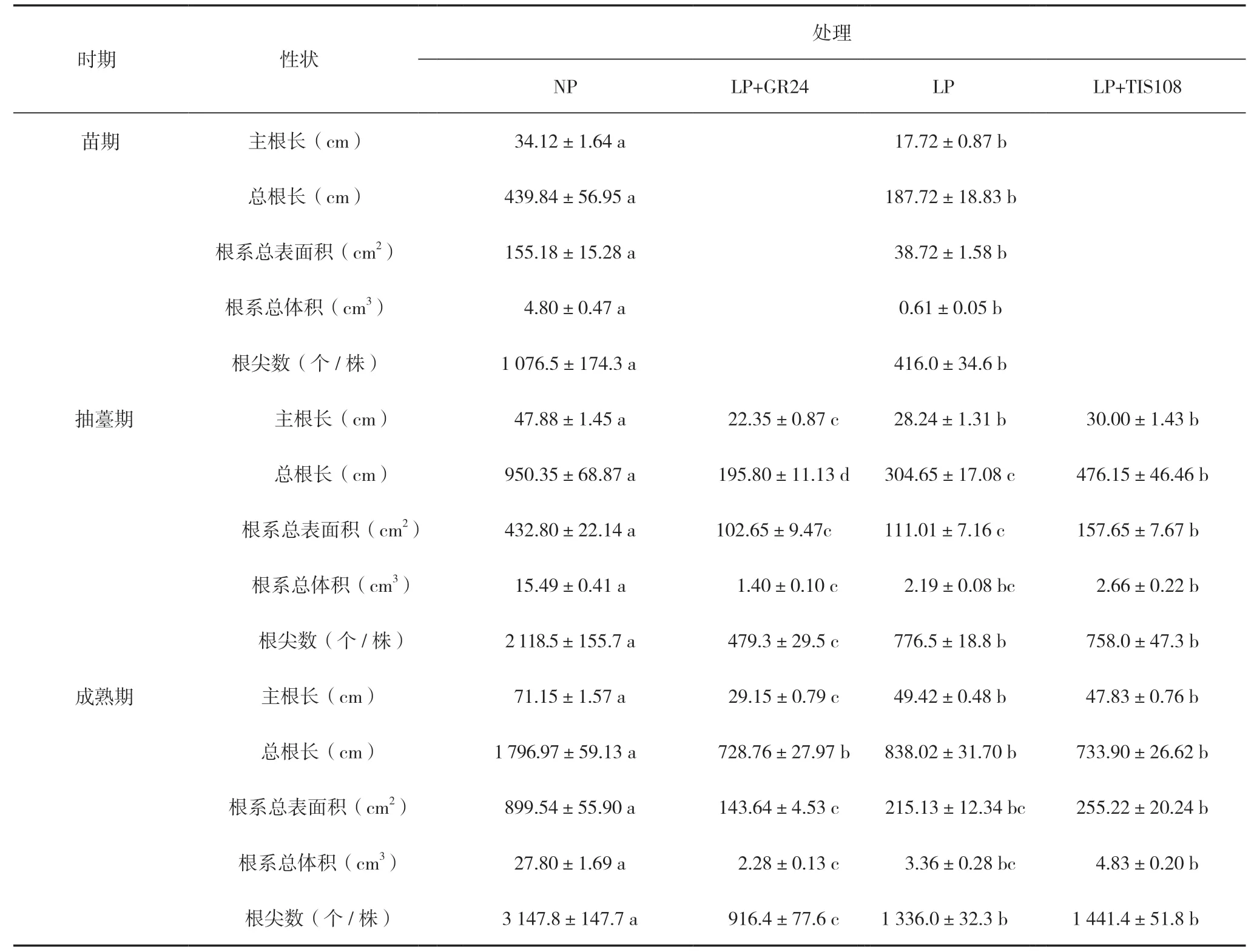
表5 低磷胁迫喷施GR24 和TIS108 中双11 号主根长、总根长、根系总表面积、根系总体积、根尖数
2.2.2 低磷胁迫喷施GR24 和TIS108 油菜地上部和根干重以及磷含量
苗期NP 处理油菜地上部干重1.63 g/株,显著高于LP 处理(0.98 g/株),但根干重NP 和LP 处理分别为0.10 和0.09 g/株,无显著差异。抽薹期和成熟期油菜NP 处理地上部和根系干重均显著大于LP+GR24、LP 和LP+TIS108 处理(图3);LP+TIS108 处理地上部干重大于LP 处理,LP 处理显著大于LP+GR24处理(图3 A);LP+TIS108 处理根系干重显著大于LP 处理,LP 处理大于LP+GR24 处理(图3 B)。
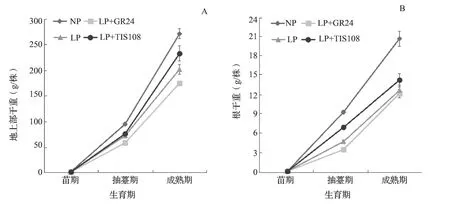
图3 低磷胁迫喷施GR24 和TIS108 中双11 号地上部和根系干重
苗期NP 处理油菜地上部和根系磷含量均显著大于LP 处理。抽薹期以及成熟期NP 处理油菜地上部和根系磷含量均显著大于LP+TIS108、LP和LP+GR24LP 处理。抽薹期地上部磷含量LP+TIS108 处理显著大于LP 和LP+GR24 处理,但LP和LP+GR24 处理磷含量无显著差异(图4 A);抽薹期根系磷含量NP 和LP+TIS108 处理之间、LP+TIS108 和LP 处理之间均无显著差异(图4 B)。成熟期地上部磷含量LP+TIS108 和LP 处理之间无显著差异,均显著大于LP+GR24 处理(图4 A)。成熟期根系磷含量LP+TIS108、LP 和LP+GR24 3 个处理之间没有显著差异(图4 B)。
苗期至抽薹期NP、LP 处理油菜地上部磷含量均呈现逐渐降低的趋势,但抽薹期至成熟期不同处理地上部磷含量均表现为增加趋势(图4 A)。苗期至抽薹期,抽薹期至成熟期不同处理根系磷含量均呈现逐渐降低的趋势(图4 B)。
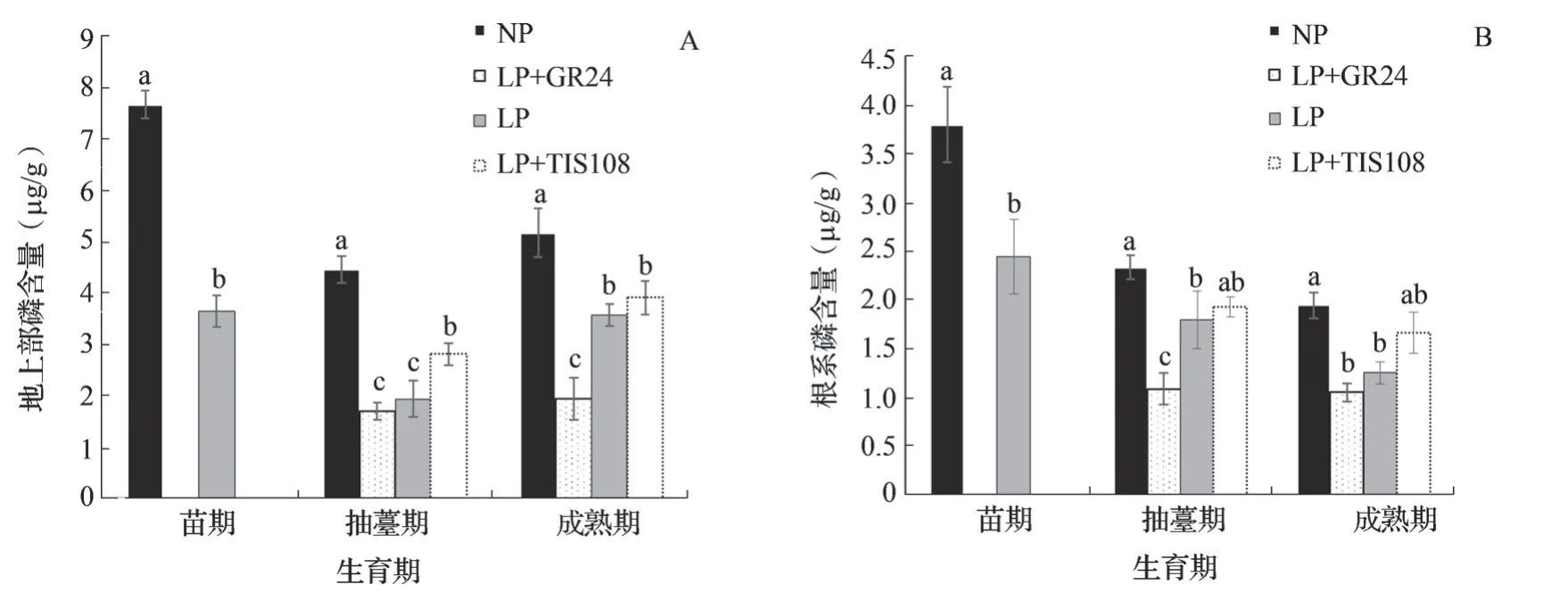
图4 低磷喷施GR24 和TIS108 中双11 号地上部和根系磷含量
2.2.3 低磷胁迫喷施GR24 和TIS108 油菜株高、分枝数和产量
NP 处理油菜株高、分枝数和产量显著高(多)于LP+GR24、LP 和LP+TIS108 处理(图5)。LP+GR24处理株高显著高于LP+TIS108 处理,但LP+GR24和LP 处理之间,LP 和LP+TIS108 处理之间株高均无显著差异(图5A)。LP+TIS108 和LP 处理分枝数无显著差异,但均显著大于LP+GR24 处理(图5 B)。 LP+TIS108 处理油菜产量显著大于LP+GR24,但LP+TIS108 和LP 处 理,LP 和LP+GR24 处 理 之间产量均没有显著差异(图5 C)。

图5 低磷胁迫喷施GR24 和TIS108 中双11 号株高、有效分枝数和产量
3 讨论
3.1 独脚金内酯对根系发生发育的影响
根系是植物吸收养分和水分的主要器官,从农业生产的角度来看,一个优良的作物品种应拥有合理的根系形态构型,才能有效利用土壤中的养分和水分,从而实现地上部的优良性状(株型、叶型、光合效率、收获指数等)[23]。独脚金内酯在植物根系的发生和发育中起着重要作用[15,24-26]。
本研究正常磷和低磷处理GR24 均抑制主根伸长和侧根发育,根系磷含量减少,根系干重下降,而TIS108 处理促进了主根伸长和侧根发育,进而增加了根系干重和根系磷含量(表3~5;图1、3、4),该研究结果与拟南芥[15,24-26]和水稻[17-18]中的研究结果一致。正常磷和低磷处理分别添加GR24 和TIS108,均表现为GR24 处理根系生长素含量降低,TIS108 处理根系生长素含量增加;并且,低磷时GR24 处理地上部生长素含量增加,TIS108 处理地上部生长素含量降低(图2),该研究结果与冯凡等水稻中的研究结果一致[14],说明独脚金内酯可能通过调控生长素从地上部向根系的极性运输进而调控植物根系生长发育。
此外,本研究营养液培养,在0 ~5 μmol/L范围内,随着独脚金内酯(GR24)浓度的增加,油菜根系和地上部生长受到的抑制越来越严重(表1)。胡超等[27]研究表明1 μmol/L GR24 能够有效缓解渍水影响,促进油菜根系和地上部干物质积累。Ma 等[28]研究表明,1.8 μmol/L GR24 能促进盐胁迫下油菜生长、光合效率,减轻氧化胁迫。本研究与该研究结果不完全一致,原因可能为两者油菜培养方法、独脚金内酯类型及其施用方式不完全一致。此外,本研究结果表明0.1 μmol/L 独脚金内酯合成抑制剂(TIS108)能够显著促进油菜根系和地上部生长(表2)。
3.2 独脚金内酯对地上部分枝、株高和产量的影响
土壤磷缺乏时,根中独脚金内酯增加,主根生长受到抑制,侧根和根毛密度和长度增加,增强根系对磷的吸收;同时,独脚金内酯也会运输到地上部,抑制地上部分枝,根冠比增大[29]。此外,在植物响应缺磷胁迫的信号通路中,独脚金内酯是能够从根向地上部运输的系统信号[30]。中双11 号成熟期NP 处理的株高、分枝数、单株产量均显著大于LP 处理(图5)。低磷胁迫喷施GR24 地上部干重和磷含量减小,喷施TIS108 地上部干重和磷含量增加(图3 和5)。油菜株高LP+GR24 和LP+TIS108 处理分别显著高于和低于LP 处理(图5),表明磷胁迫条件下喷施GR24 对油菜株高有促进作用,喷施TIS108 对株高有抑制作用。油菜分枝数和产量LP+TIS108 和 LP+GR24 处理显著多(高)于LP 处理(图5),表明低磷胁迫喷施GR24油菜分枝减少,产量降低;喷施TIS108 油菜分枝数增加,产量增加。该结果与水稻中的研究结果一致[31-32],说明独脚金内酯能够通过抑制分枝作用降低产量。分枝是植物株型决定的重要因素,生理上主要由生长素、独脚金内酯和细胞分裂素3 类激素调节。独脚金内酯的分枝抑制作用主要通过两种方式实现:一种是在水稻、豌豆中通过 TCP 转录因子OsTB1/PsBRC1 在独脚金内酯下游作用抑制腋芽外生;另一种是在拟南芥中独脚金内酯触发茎木质部薄壁细胞质膜上的生长素外运载体PIN1 快速移动来减少生长素极性运输[9,33-35]。低磷胁迫、喷施独脚金内酯抑制剂TIS108 可以通过增加分枝数来减少低磷胁迫对油菜产量的影响。
4 结论
不同磷水平施用外源GR24(5 μmol/L)均抑制油菜根系生长,表现为主根长变短,总根长、根系总表面积、根系总体积、根尖数减小(少、短),根干重减小,根系磷浓度和生长素浓度降低,地上部生长素浓度增加,独脚金内酯可能通过抑制生长素从地上部向根系的极性运输,进而抑制植物根系生长。不同磷水平外源施用TIS108(0.1 μmol/L)均能促进油菜根系生长,主要表现为主根长变长,总根长、根系总表面积、根系总体积、根尖数增大(加、长),根干重增加,根系磷浓度和生长素含量增加,地上部生长素浓度减小,独脚金内酯合成抑制剂可能通过促进生长素从地上部向根系的极性运输,进而促进植物根系生长。低磷胁迫喷施GR24 株高增加,分枝数减少,产量降低;喷施TIS108 株高降低,分枝数增加,产量增加。分离克隆油菜中独脚金内酯合成相关基因及其调控因子,揭示不同时期这些基因的功能,将进一步明确减磷条件下油菜独脚金内酯的作用及其合成抑制剂的应用前景。
