胡萝卜YUCCA基因家族鉴定及生物信息学分析
2021-01-02闫学敏吴英华史艳武喆侯雷平李梅兰
闫学敏 吴英华 史艳 武喆 侯雷平 李梅兰

摘要:胡萝卜是重要的根菜类蔬菜之一,根部性状影响最终的产量和品质。YUCCA基因通过介导生长素调控植物根部发育,是生长素合成途径中的关键基因。利用生物信息学方法对胡萝卜YUCCA基因家族成员进行鉴定,对其理化性质、染色体定位、系统进化树、蛋白质的二级和三级结构以及保守基序进行分析。结果表明,在胡萝卜的5条染色体上共鉴定到了14个YUCCA基因家族成员,其大部分基因含有3~4个外显子;该家族编码的蛋白质为富含碱性氨基酸的亲水性蛋白质,氨基酸数量为290~424个,其蛋白质结构主要以α-螺旋和无规则卷曲构成,通过构建系统进化树可将其分为3个亚族,且亚细胞定位结果显示,YUCCA基因家族蛋白质大部分被定位在细胞质中。这为YUCCA基因家族在调控胡萝卜根发育和膨大机制研究奠定了理论基础。
关键词:胡萝卜;生长素;YUCCA基因家族;生物信息学;蛋白质结构;染色体定位
中图分类号:S631.201 文献标志码: A
文章编号:1002-1302(2021)23-0052-06
收稿日期:2021-07-16
基金项目:山西省重点研发计划重点项目子课题(编号:201703D211001-04-01);山西省重点研发计划(编号:201903D221063)。
作者简介:闫学敏(1997—),女,山西吕梁人,硕士研究生,主要从事蔬菜育种及生物技术应用研究。E-mail:464679520@qq.com。
通信作者:李梅兰,博士,教授,主要从事蔬菜育种及生物技术应用研究。E-mail:15935485975@163.com。
植物根系的生长发育存在着复杂的调控网络,受到环境、生理、激素等多种因素的影响[1]。近年来,植物激素作为调控生长发育的一大类物质而被研究者广泛关注,生长素是最早发现的植物生长类激素,其代谢调控和信号转导机制研究较为广泛[2]。其中,YUCCA基因家族已经被证明调控植物的根系发育,在生长素生物合成的吲哚丙酮酸途径中YUCCA基因家族是重要酶之一,其基因编码黄素单加氧酶(flavin-containing monooxygenase,简称FMOs)[3-4]。YUCCA基因首次被鉴定是在拟南芥生长素过量的突变体研究中[5],目前该基因家族已经在烟草[6]、拟南芥[7]、水稻[8]、草莓[9]等作物中进行了全基因组鉴定分析,但在胡萝卜中还未见类似的报道。
胡萝卜(Daucus carota var. sativa DC.)是以肉质根为食用器官的二年生草本植物,是全球性十大蔬菜中的一种。根是植物在进化过程中适应陆地生活而发展起来的营养器官,胡萝卜作为重要的根菜类蔬菜之一,研究其根部的生长发育对提高胡萝卜的品质和产量至关重要。有研究表明,生长素在植物根尖形成的梯度浓度调控其合成和运输,从而影响植物的根际发育[10]。因此,本试验利用生物信息学方法鉴定胡萝卜YUCCA基因家族,对其理化性质、染色体定位、系统进化树、蛋白质的二级结构和三级结构以及保守基序进行分析,进一步为胡萝卜YUCCA基因功能验证提供一定的理论基础。
1 材料与方法
1.1 胡萝卜YUCCA基因家族数据获取与鉴定
通过拟南芥官网TAIR(https://www.arabidopsis.org/)获取已报道过的YUCCA基因家族成员的蛋白质序列,在NCBI网站(https://www.ncbi.nlm.nih.gov/)使用BlastP检索,设置E-value值为10-5,获得拟南芥、烟草、玉米和胡萝卜中的YUCCA基因家族候选基因的蛋白质序列和基因序列的fasta格式。通过CDD和SMART数据库比对验证候选基因的蛋白质序列结构域[11],最终获得11个拟南芥YUCCA基因、20个烟草YUCCA基因、30个玉米YUCCA基因和14个胡萝卜YUCCA基因,并对其分别命名。
1.2 胡萝卜YUCCA蛋白质理化性质分析
将筛选出的14个胡萝卜YUCCA基因家族蛋白质序列通过ExPASy(https://web.expasy. org/protparam/)在线网站分析其理化性质[12]。
1.3 胡萝卜YUCCA基因的染色体定位
从NCBI上获取基因的染色体位置信息,使用在线网站MG2C_V2.0(http://mg2c.iask.in/mg2c_v2.0/)对基因进行染色体定位绘图并简单修饰。
1.4 拟南芥、烟草、玉米和胡萝卜YUCCA基因家族系统进化树分析
在软件MEGA 7.0中对比4个物种的蛋白质序列,采用邻接法构建进化树,bootstrap设为1 000[13]。用在线网站iTOL(https://itol.embl.de/itol.cgi)对进化树进行环化。
1.5 胡萝卜YUCCA蛋白质二、三级结构预测及亚细胞定位
通过在线网站SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和CELLO(http://cello.life.nctu.edu.tw/)分別对胡萝卜YUCCA基因家族的蛋白质序列进行二级结构预测和亚细胞定位。通过在线网站SWISS-MODEL(https://swissmodel. expasy.org/)对胡萝卜YUCCA基因家族的蛋白质序列进行三级结构模型分析[14]。
1.6 胡萝卜YUCCA基因家族的基因结构及蛋白质保守基序分析
将14个基因的CDS和FASTA文件导入在线网站GSDS 2.0(http://gsds.gao-lab.org/index.php)进行胡萝卜YUCCA基因结构分析[15]。使用MEME在线网站(https://meme-suite.org/meme/tools/meme)和TBtools对胡萝卜YUCCA蛋白质的保守基序进行分析[16]。
2 结果与分析
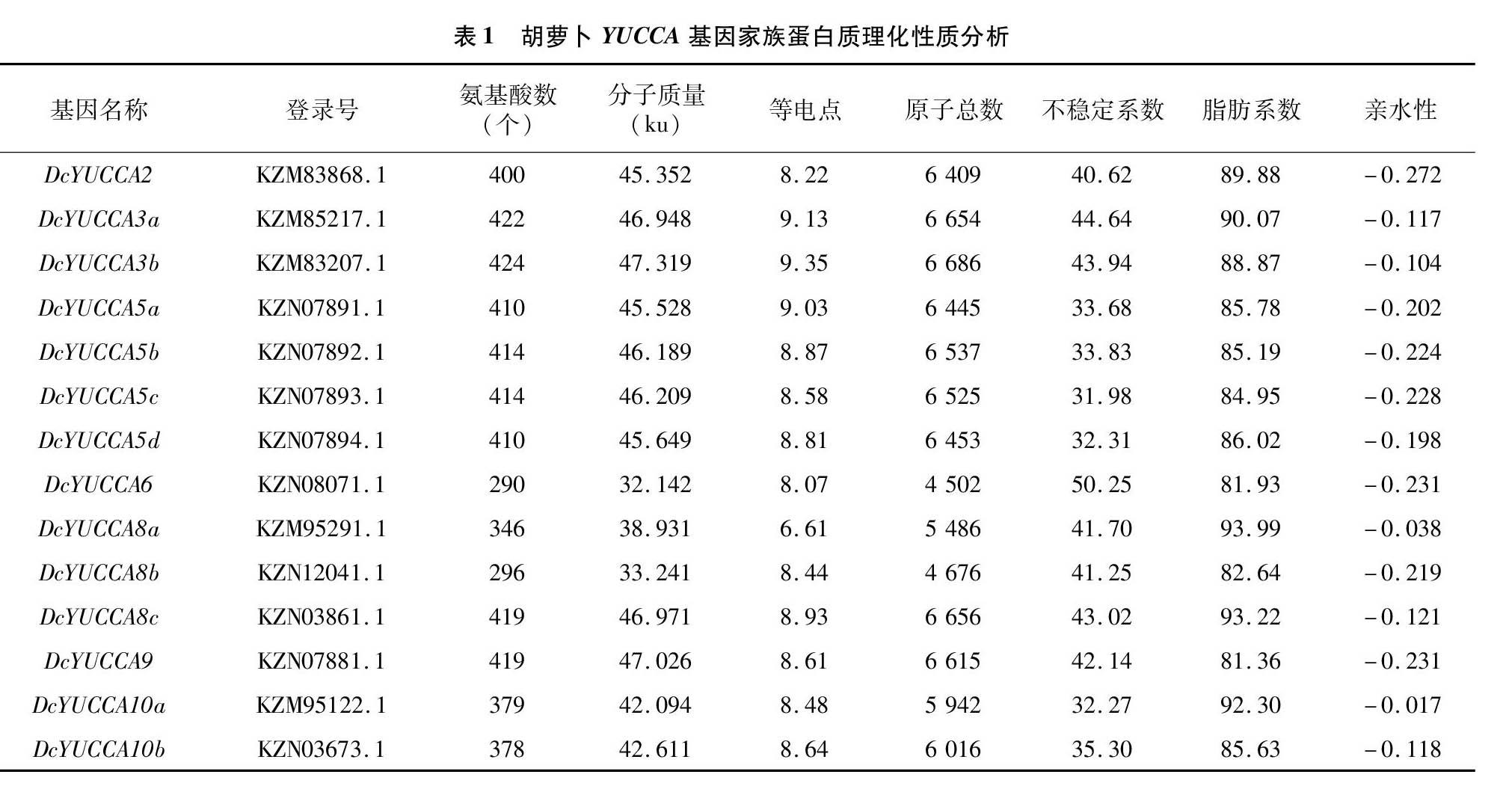
2.1 胡萝卜YUCCA蛋白质理化性质
通过对胡萝卜YUCCA基因家族的蛋白质进行理化性质分析(表1)可知,YUCCA蛋白质的氨基酸数为290~424个,相对分子质量为32.14~47.32 ku,氨基酸数和分子质量最大的均为蛋白质DcYUCCA3b,最小的均为蛋白质DcYUCCA6;等电点为6.61~935;不稳定系数为31.98~50.25,其中蛋白质DcYUCCA5a、DcYUCCA5b、DcYUCCA5c、DcYUCCA5d、DcYUCCA10a和DcYUCCA10b均为稳定蛋白,其余的蛋白质(DcYUCCA2、DcYUCCA3a、DcYUCCA3b、 DcYUCCA6、 DcYUCCA8a、 DcYUCCA8b、DcYUCCA8c和DcYUCCA9)为不稳定蛋白;亲水性均为负值,说明胡萝卜中YUCCA蛋白质均为亲水性蛋白。
2.2 胡萝卜YUCCA基因的染色体定位
胡萝卜共有9条染色体。染色体定位结果(图1)显示,胡萝卜YUCCA基因家族中有7个基因被定位在1号染色体上,分布最多,基因存在聚集现象,说明这类基因功能具有一定的相似性。其次,有2个基因被定位在3号染色体上,2个基因被定位在5号染色体上,2个基因被定位在8号染色体上,1个基因被定位在9号染色体上。
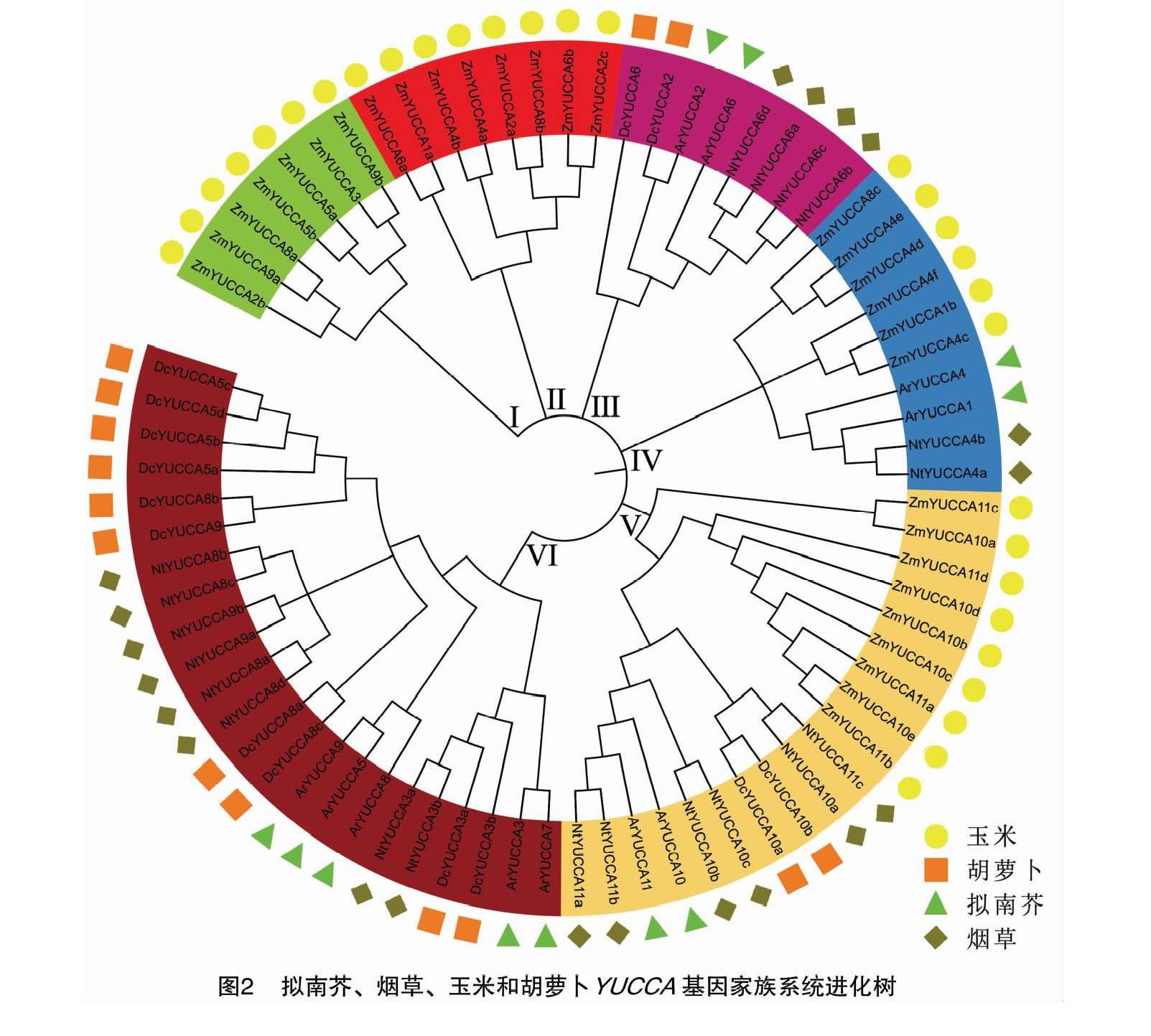
2.3 拟南芥、烟草、玉米和胡萝卜YUCCA基因家族系统进化树
为进一步了解胡萝卜YUCCA基因家族进化情况,选用拟南芥(11个)、烟草(20个)、玉米(30个)YUCCA基因家族基因为参考序列构建系统进化树(图2),结果分为6个亚族。其中胡萝卜YUCCA基因家族主要分布在Ⅵ亚族中,有10个基因,其次为Ⅲ和Ⅴ亚族中。
2.4 胡萝卜YUCCA蛋白质二、三级结构预测及亚细胞定位
对胡萝卜YUCCA蛋白质的二级结构预测结果(表2)显示,14个蛋白质主要以α-螺旋和无规则卷曲构成。为了验证二级结构的结果准确性,对14个蛋白质的三级结构进行模型预测(图3),结果显示三级结构与二级结构基本一致。蛋白质的亚细胞定位结果显示大多数的蛋白质均被定位在细胞质中(13个),还有少部分的蛋白质被定位在周质和外膜上。
2.5 胡萝卜YUCCA基因结构
为了分析胡萝卜YUCCA基因家族的功能,对14个基因进行基因结构分析(图4),结果得出基因的长度不同,外显子和内含子也存在差别。基因DcYUCCA2、DcYUCCA6、DcYUCCA10a和DcYUCCA10b均含有4个外显子和3个内含子,其中基因DcYUCCA10a无上下游;基因DcYUCCA3a、DcYUCCA3b、DcYUCCA5a、DcYUCCA5b、DcYUCCA5c、DcYUCCA5d、DcYUCCA8a、DcYUCCA8b和DcYUCCA8c含有3个外显子和2个内含子,其中基因DcYUCCA5a、DcYUCCA5b、DcYUCCA5c、DcYUCCA5d和DcYUCCA8b无上下游;基因DcYUCCA9含有2個外显子和1个内含子。
2.6 胡萝卜YUCCA基因家族蛋白质保守基序
对胡萝卜YUCCA基因家族蛋白质序列进行保守基序分析(图5),发现14个蛋白质的保守结构域为6~10个。蛋白质DcYUCCA3a、DcYUCCA3b、DcYUCCA5a、DcYUCCA5b、DcYUCCA5c、DcYUCCA5d、DcYUCCA8c和DcYUCCA9含有10个motif,结构和功能最为相似,14个YUCCA蛋白质中均含有motif 1、motif 7和motif 3。蛋白质DcYUCCA2不含motif 9、motif 6和motif 10;蛋白质DcYUCCA6不含motif 5、motif 4、motif 6和motif 10;蛋白质DcYUCCA10a和DcYUCCA10b结构基本一致,均不含motif 8、motif 9和motif 10;蛋白质DcYUCCA8a不含motif 9、motif 4和motif 10;蛋白质DcYUCCA8b不含motif 2、motif 5和motif 6。
3 讨论与结论
植物根部的生长发育是一个动态变化的过程,是由根尖分生区细胞的分裂和伸长区细胞的伸长而生长的,在这个过程中植物激素发挥了重要的作用[17]。生长素是其中一类生长调节激素,可以促进细胞的伸长、木质部和韧皮部的分化,从而使根部增粗[18]。在胡萝卜中,研究根部膨大增粗可以提高其品质和产量,提高经济效益。YUCCA基因家族蛋白是植物中第一个被鉴别的FMO类超家族,调控生长发育和植物的耐受性[19]。有研究证实YUCCA基因介导生长素的生物合成,在拟南芥中抑制YUCCA表达会抑制生根,调控根器官发育[20]。
本研究利用生物信息学方法鉴定胡萝卜YUCCA基因家族成员共14个,与拟南芥YUCCA基因家族的11个成员数量较接近。通过理化性质分析得出,胡萝卜YUCCA基因家族蛋白质基本为碱性氨基酸(除了蛋白质DcYUCCA8a),且14个蛋白质均为亲水性蛋白质,基因分别定位在5条染色体上(1、3、5、8、9号),其中在1号染色体上较聚集,结构和功能最为相似。系统进化树中聚集在一起的亚族具有相似的进化源和基因功能,通过构建系统进化树将胡萝卜YUCCA基因家族分为3个亚族(Ⅲ、Ⅴ、Ⅵ亚族),与烟草和拟南芥的亲缘关系较近,其中,Ⅲ亚族中的DcYUCCA2和DcYUCCA6与拟南芥的ArYUCCA2和ArYUCCA6亲缘关系最近,推测可能通过调控生长素信号转导介导植物的生长发育和早期花粉发育[21]。
蛋白质的结构决定其生物学功能,结构相似其功能相似,在分析蛋白质的二级结构和三级结构模型时发现,YUCCA基因家族蛋白质主要以α-螺旋和无规则卷曲构成,位于同一家族成员的结构具有一致性。在亚细胞定位中发现大部分的蛋白质被定位在细胞质中,表明这些基因合成生长素基本上是在细胞质中完成的,这与王仁汉等的结果[6,22]一致。通过分析胡萝卜YUCCA基因家族蛋白质的保守基序发现,14个蛋白质的motif有6~10个不等,说明基因家族功能的多样性,但其中有大部分YUCCA蛋白质的motif数量均为10个,说明基因家族的功能具有相似性。总体来看,位于同一基因家族的成员在植物中的结构和功能均具有相似性,为进一步研究基因家族提供参考,为胡萝卜根部膨大形成机制的研究奠定了理论基础。
本研究在胡萝卜的5条染色体上共鉴定到了14个YUCCA基因家族成员,其大部分基因含有3~4个外显子;该家族编码的蛋白质为富含碱性氨基酸的亲水性蛋白质,氨基酸数量为290~424个,其蛋白质结构主要以α-螺旋和无规则卷曲构成,通过构建系统进化树可将其分为3个亚族,且亚细胞定位结果显示,YUCCA基因家族蛋白质大部分被定位在细胞质中。这为YUCCA基因家族在调控胡萝卜根发育和膨大机制研究奠定了理论基础。
参考文献:
[1]马 凯,杨 凡,蔡毓新,等. 蔬菜不定根形成调控机制研究进展[J]. 中国瓜菜,2017,30(3):1-4.
[2]Zhao Y D.Auxin biosynthesis and its role in plant development[J]. Annual Review of Plant Biology,2010,61:49-64.
[3]Brumos J,Alonso J M,Stepanova A N.Genetic aspects of auxin biosynthesis and its regulation[J]. Physiologia Plantarum,2014,151(1):3-12.
[4]Qin M M,Wang J,Zhang T Y,et al. Genome-wide identification and analysis on YUCCA gene family in Isatis indigotica Fort.and IiYUCCA6-1 functional exploration[J]. International Journal of Molecular Sciences,2020,21(6):2188.
[5]Zhao Y,Christensen S K,Fankhauser C,et al. A role for flavin monooxygenase-like enzymes in auxin biosynthesis[J]. Science,2001,291(5502):306-309.
[6]王仁汉,宋志美,屈 旭,等. 普通烟草YUCCA基因家族的生物信息学分析[J]. 江苏农业科学,2021,49(3):61-65.
[7]Cheng Y F,Dai X H,Zhao Y D. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis[J]. Genes & Development,2006,20(13):1790-1799.
[8]Yamamoto Y,Kamiya N,Morinaka Y,et al. Auxin biosynthesis by the YUCCA genes in rice[J]. Plant Physiology,2007,143(3):1362-1371.
[9]Liu H,Xie W F,Zhang L,et al. Auxin biosynthesis by the YUCCA6 flavin monooxygenase gene in woodland strawberry[J]. Journal of Integrative Plant Biology,2014,56(4):350-363.
[10]Cao X,Yang H L,Shang C Q,et al. The roles of auxin biosynthesis YUCCA gene family in plants[J]. International Journal of Molecular Sciences,2019,20(24):6343.
[11]Wang R,Liu L,Kong Z S,et al. Identification of GhLOG gene family revealed that GhLOG3 is involved in regulating salinity tolerance in cotton (Gossypium hirsutum L.)[J]. Plant Physiology and Biochemistry,2021,166:328-340.
[12]Jiang S,Li S,Liu X A,et al. Genome-wide identification and characterization of the MdBZR1 gene family in apple and their roles in improvement of drought tolerance[J]. Scientia Horticulturae,2021,288:110359.
[13]Zheng L W,Ma S J,Zhou T,et al. Genome-wide identification of Brassicaceae B-BOX genes and molecular characterization of their transcriptional responses to various nutrient stresses in allotetraploid rapeseed[J]. BMC Plant Biology,2021,21(1):288.
[14]羅康胜,张米欢,翟兆东,等. 生菜WOX基因家族的鉴定与分析[J/OL]. 分子植物育种:1-18[2021-10-12]. http://kns.cnki.net/kcms/detail/46.1068.S.20210622.1012.002.html.
[15]Haider M S,de Britto S,Nagaraj G,et al. Genome-wide identification,diversification,and expression analysis of lectin receptor-like kinase (LecRLK) gene family in cucumber under biotic stress[J]. International Journal of Molecular Sciences,2021,22(12):6585.
[16]李明明,晋敏姗,胡海斌,等. 谷子MYB-CC基因家族的鉴定与表达分析[J]. 激光生物学报,2021,30(3):259-269.
[17]王柳萍,覃坤坚,赵立春,等. 植物根膨大的研究进展[J]. 湖北农业科学,2020,59(11):5-9.
[18]邹 禹,刘园园,张培江,等. 水稻富亮氨酸重复类受体蛋白激酶OsRPK1响应外源生长素的作用研究[J]. 江苏农业科学,2019,47(8):46-51.
[19]Cha J Y,Kim W Y,Kang S B,et al. A novel thiol-reductase activity of Arabidopsis YUC6 confers drought tolerance independently of auxin biosynthesis[J]. Nature Communications,2015,6:8041.
[20]Chen L Q,Tong J H,Xiao L T,et al. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis[J]. Journal of Experimental Botany,2016,67(14):4273-4284.
[21]Yao X Z,Tian L,Yang J,et al. Auxin production in diploid microsporocytes is necessary and sufficient for early stages of pollen development[J]. PLoS Genetics,2018,14(5):e1007397.
[22]綦 洋,王柬鈞,桑园园,等. 大白菜YUCCA基因家族的鉴定与生物信息学分析[J]. 江苏农业科学,2019,47(3):49-54.
