假单胞菌Pseudomonas sp. AT39反式茴脑氧化酶基因(tao)对产茴香酸的影响
2020-12-28黄罗冬华燕飞申佩弘
蒋 琼, 武 波, 黄罗冬, 华燕飞, 申佩弘,2*
1.广西大学生命科学与技术学院,广西微生物与酶工程技术研究中心,广西 南宁 530004;2.亚热带农业生物资源保护与利用国家重点实验室,广西 南宁 530004;3.广西民族师范学院,广西 崇左 532200
茴香酸(anisic acid),化学名为4-甲氧基苯甲酸(4-Methoxybenzoic Acid),是合成香料的主要原料,同时也是防腐、制药等领域方面的重要原料,是胺碘酮、茴拉西坦等的医药中间体,因而被人们关注及合成应用[1-3],反式茴脑可被微生物代谢合成茴香酸及茴香醛。国内外关于微生物利用反式茴脑合成茴香酸和茴香醛的研究报道并不多,已报道的能以反式茴脑为唯一碳源进行生长代谢的菌株主要有:AspergillusnigerZJ-9[2],Pseudomonassp.BT13[2],PseudomonasputidaWGAF10[4],Burkholderiasp. WGB31[5],且关注的是优化发酵条件提高反式茴脑的中间代谢产物茴香醛及茴香酸的产量。对反式茴脑代谢相关基因或途径进行研究的菌株主要有Arthrobactersp. TA13[6,7]和PseudomonasputidaJYR-1[8-11]。根据研究报道,Arthrobactersp. TA13代谢反式茴脑生成对-茴香醇是通过形成环氧化物或二醇的中间体而实现的,但由于其体内缺乏脱甲氧基酶,因此不能断裂苯环实现对茴脑的进一步利用。HAN Dongfei等人[9-11]对PseudomonasputidaJYR-1反式茴脑代谢途径进行了研究,首次克隆出tao基因,并于大肠杆菌体内进行异源性表达,推断反式茴脑氧化酶(TAO)为黄素蛋白单氧化酶,可将反式茴脑催化氧化成对-甲氧基茴香醛,且反应过程需NAD+和NADP+作为辅助因子。然而HAN Dongfei等人[9-11]对tao基因的功能研究是通过大肠杆菌的异源表达TAO酶来实现的,研究的范围仅限于该基因,并不能真正了解该基因对反式茴脑整个代谢途径的影响,以及与其他相关基因的作用关系。本研究拟以Pseudomonassp. AT39为目的菌株,构建tao基因的重组菌和pk18mob插入的突变菌及其互补菌,进行TAO酶比活力测定及反式茴脑代谢产物生成量的检测,以确定tao基因在Pseudomonassp.AT39反式茴脑代谢过程中的地位及作用,也为工业合成茴香酸奠定基础。
1 材料与方法
1.1 菌株与质粒
实验所用的菌株和质粒:E.coliDH5α,菌株Pseudomonassp.AT39为本实验室分离,克隆载体pMD18-T 购自TaKaRa 公司,质粒pRK2013、pBBR1MCS-5[12]和pK18mob[13]均由实验室保存。
1.2 培养基和生长条件
大肠杆菌DH5α采用LB培养基,37 ℃培养。野生型Pseudomonassp. AT39菌株采用LM9培养基和种子培养基[14],30 ℃培养,实验过程根据不同实验目的需要在LM9培养基中加入不同浓度反式茴脑。实验过程抗生素使用浓度 (mg/mL)为:氨苄青霉素(Amp)50;卡那霉素(Km)40;羧苄青霉素(Cb)100。
1.3 酶、试剂和引物
2x Taq MasterMix(含染料)和细菌基因组 DNA提取试剂盒购于北京康为世纪生物科技有限公司;Ex-Taq聚合酶和 T4-DNA ligase购自大连 TaKaRa公司;质粒 DNA提取试剂盒、胶回收试以及PCR产物纯化试剂盒均购自于西宝生物科技股份有限公司;限制性内切酶及其缓冲液均购于Fermentas公司。实验所用引物列于表1。

表1 实验所用的引物
1.4 方法
1.4.1Pseudomonassp. AT39重组表达菌株的构建
以连接在T-载体且测序结果正确的tao基因为模板, 以tao-F、tao-R为引物,PCR扩增tao基因片段,并酶切后连接至以同样酶切处理的pBBR1MCS-5质粒上,转化至大肠杆菌DH5α感受态细胞中,构建重组菌DH5α/tao-pBBR1MCS5。以野生型Pseudomonassp. AT39为受体菌,DH5α/tao-pBBR1MCS5为供体菌,DH5α/pRK2013为帮助菌进行菌体量为1∶1∶1的三亲本接合实验[15],将重组质粒tao-pBBR1MCS5导入Pseudomonassp. AT39中,并进行质粒的提取及酶切验证。
1.4.2P.putidaAT39菌株pk18mob插入突变体的构建
同样以tao-T重组质粒为模板,Δtao-F、Δtao-R为引物(分别含EcoR I、XbaI酶切位点),PCR扩增tao基因部分片段btao(约425 bp),连接同样酶切处理过的pK18mob质粒,并转化至大肠杆菌DH5α感受态细胞中,构建菌株DH5α/ btao-pK18mob。参照方法1.4.2中三亲本接合实验操作,将自杀质粒pk18mob插入Pseudomonassp.AT39基因组中,并进行PCR验证。
1.4.3互补菌株的构建
参照方法1.4.2中的三亲本接合实验操作将重组质粒tao-pBBR1MCS5导入突变株AT39/btao中,挑选具有Cbr和Kmr双抗性的重组子接种到液体种子培养基培养12~16 h,提取质粒并进行PCR或酶切验证。
1.4.4菌株生长情况测定
按1%(v/v)的接种量将过夜培养的菌液接种到新鲜的液体种子培养基中,30 ℃,180 r/min振荡培养,每隔2 h取样检测OD600值,测定菌株的生长曲线。
1.4.5TAO酶活的测定
按1%(v/v)的接种量将过夜培养的菌液接种至LB培养基中,30 ℃,180 r/min培养至OD600为0.6时,加入终浓度为1.0 mmol/L的IPTG,继续培养直至菌株的生长稳定期。离心后,以50 mmol/L磷酸钾缓冲液(pH 8.0)悬浮沉淀并超声波破碎,所用振幅39%,总时间50 min(每隔5 s停顿5 s)。离心后的上清即为粗酶液。
酶活测定方法参考文献[11,16]稍作修改。1 mL的反应体系中含有50 mmol/磷酸钾缓冲液(pH 8.0),20 mmol/L NADH,20 mmol/L FAD,100 mmol/L 反式茴脑,30 ℃,180 r/min振荡反应1 h,沸水浴10 min,以超纯水为空白对照。对酶反应液进行萃取,用高效液相色谱法对底物反式茴脑的减少量进行检测且酶活力计算。
酶活定义:在30 ℃,pH 8.0条件下,每分钟转化1 μmol/L的反式茴脑所需酶量为1个酶活单位1 U,即1 U。
酶比活力(U/g)=A/(t×m)
注:A为样品中反式茴脑减少的量(μmol),t为反应时间(min),m为酶的质量(g)。
1.4.6高效液相色谱法(HPLC)分析
按1%(v/v)的接种量将过夜培养的菌液接种到以1%(v/v)反式茴脑为唯一碳源的LM9培养基中,30 ℃,180 r/min振荡培养。待菌株OD600值达到0.6~0.8时,加入终浓度为1.0 mmol/L的IPTG,30 ℃,180 r/min继续振荡培养36 h。进而对菌液进行萃取以及高效液相色谱的测定,具体实验操作及数据处理方法参考文献[4,14]。
2 结果与分析
2.1 Pseudomonas sp. AT39重组表达菌株、突变菌株及互补菌株的构建
利用两端分别含有XhoI和EcoR I酶切位点的引物tao-F和tao-R扩增tao基因,双酶切纯化后与同样处理的pBBR1MCS-5质粒连接,转化至大肠杆菌DH5α感受态中,通过含有Gmr抗性的X-gal和IPTG培养基进行筛选重组转化子。并使用限制性内切酶XhoI和EcoR I进行酶切验证(图1)。以野生菌P.putidaAT39为受体菌,DH5α/tao-pB-BR1MCS5为供体菌,DH5α/pRK2013为帮助菌进行三亲本接合实验,将重组子质粒导入野生菌中,构建重组菌AT39/tao-pBBR1MCS5。
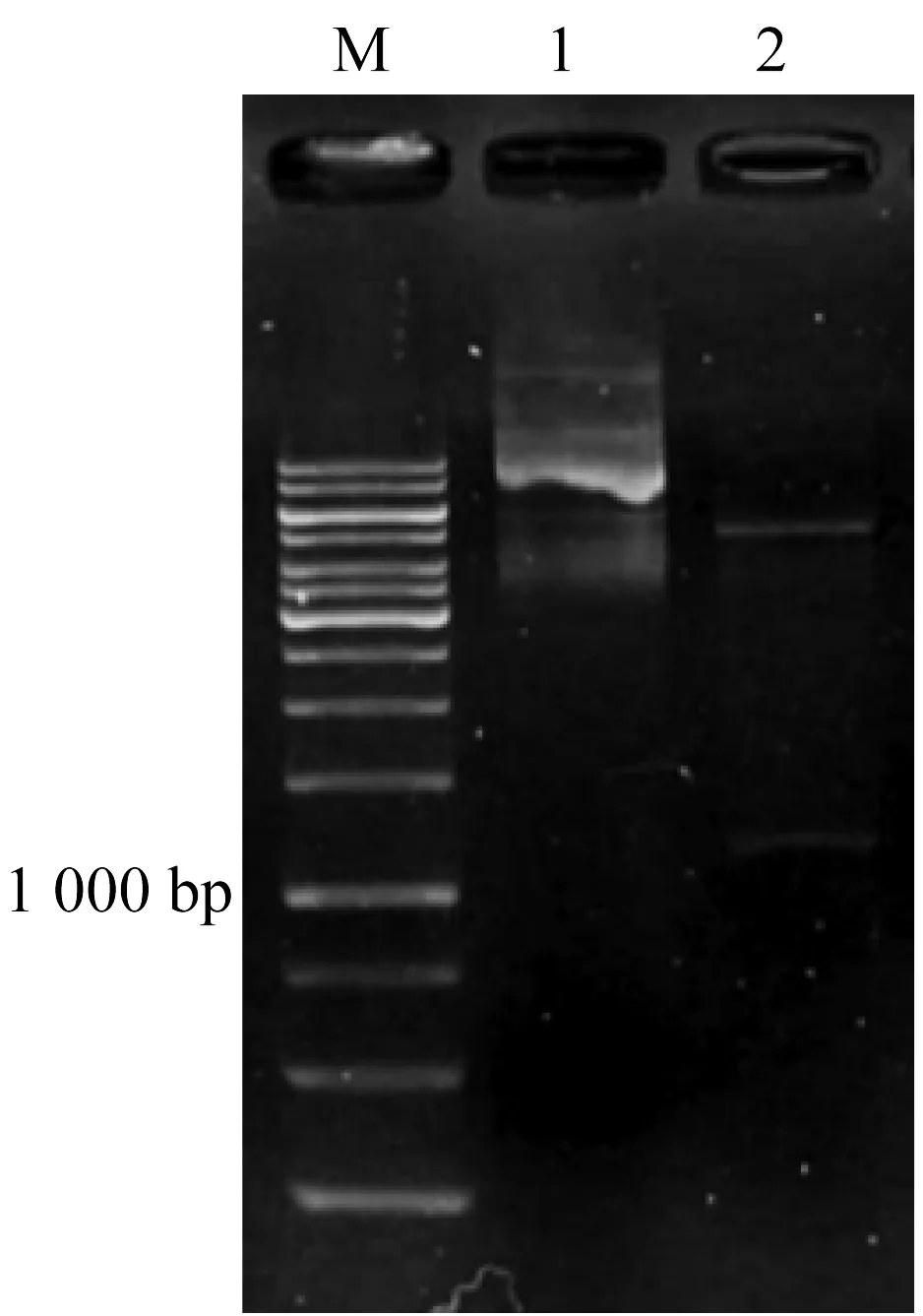
M: 1 000 b ladder DNA; 1: tao-pBBR1MCS5质粒; 2: tao-pBBR1MCS5质粒的Xho I 和EcoR I酶切图1 重组质粒tao-pBBR1MCS5 的Xho I和EcoR I酶切验证
以野生菌Pseudomonassp.AT39为受体菌,DH5α/tao-pBBR1MCS5为供体菌,DH5α/pRK2013为帮助菌进行三亲本接合实验,将重组子质粒tao-pBBR1MCS5导入野生菌中,构建重组菌AT39/tao-pBBR1MCS5。
按实验操作,得到转化子DH5α/btao-pK18mob,提取重组质粒酶切验证(图2)。参照方法1.4.1三亲本实验操作,将自杀质粒pK18mob插入到野生型AT39菌株基因组中,提取基因组DNA进行PCR验证。
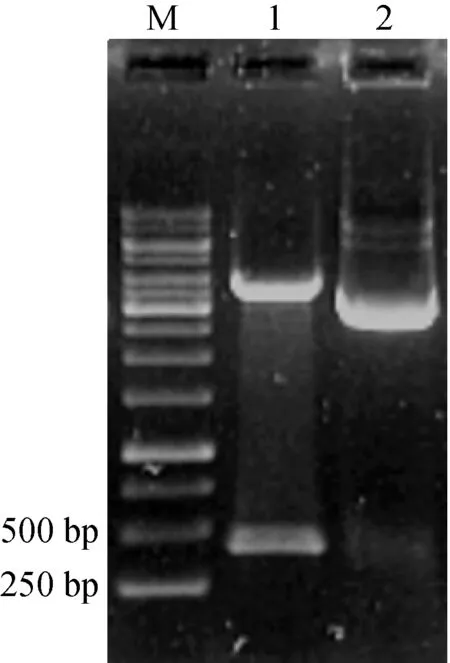
同样参照1.4.1三亲本实验操作,将重组质粒tao-pBBR1MCS5导入突变菌AT39/btao中,构建互补菌HAT39/btao。提取互补菌重组质粒tao-pBBR1MCS5并进行酶切验证。
2.2 菌株生长情况分析
对菌株AT39、AT39/pBBR1MCS5、AT39/btao、AT39/tao-pBBR1MCS5、HAT39/btao的生长曲线进行测定。结果表明,在种子培养基中,突变菌、重组菌以及突变互补菌到达稳定生长期的时间与野生菌的相差不大,与野生菌间的差值约为2 h。
2.3 酶活力测定
测定并计算菌株AT39、 AT39/btao、AT39/pBBR1MCS5、AT39/tao-pBBR1MC S5、HAT39/btao中TAO酶的比活力。结果如表2所示。

表2 各菌株中酶比活力的测定
根据表2中数据显示,重组菌AT39/pBBR1MCS5 的TAO酶比活力同比野生菌AT39并未有显著性差异。而重组菌AT39/tao-pBBR1MCS5比野生菌AT39酶比活力增加了35.23%,表明tao基因在lac启动子作用下表达量增加,对反式茴脑的代谢作用增强,是反式茴脑代谢途径中一个关键基因。突变菌AT39/btao比野生菌AT39酶比活力下降了19.67%,其互补菌HAT39/btao比野生菌AT39酶活力增加了26.79%,而相对于突变菌AT39/btao,酶比活力增加了57.83%。
2.4 茴香酸产量分析
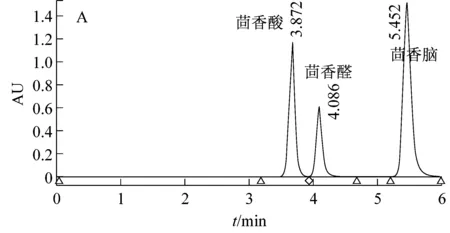
利用高效液相色谱法对Pseudomonassp. AT39野生型、重组型及突变型菌株发酵液中茴香酸进行检测,如图3所示。
图3显示野生菌AT39和空质粒重组菌AT39/pBBR1MCS5代谢反式茴脑36 h后茴香酸产量分别是0.706 g/L和0.695 g/L,茴香酸摩尔生成率分别为6.87%和6.77%,茴香脑的摩尔利用率分别为78.73%和88.48%。数据显示两者之间并未有显著性差异,表明转入的空质粒pBBR1MCS5对实验结果影响不大。与此同时,重组菌AT39/tao-pBBR1MCS5代谢生成的茴香酸的量达到3.55 g/L,茴香酸摩尔生成率为34.56%,茴香脑摩尔利用率为86.42%。与已报道的P.putidaWGAF10[4]发酵条件优化后产茴香酸的量相比增加了45.5%。茴香酸摩尔生成率同比野生菌AT39增加了约4倍,比P.putidaWGAF10增加了42.22%。茴香脑的摩尔利用率同比野生菌AT39并未有显著差异,而同比P.putidaWGAF10增加了46.18%。


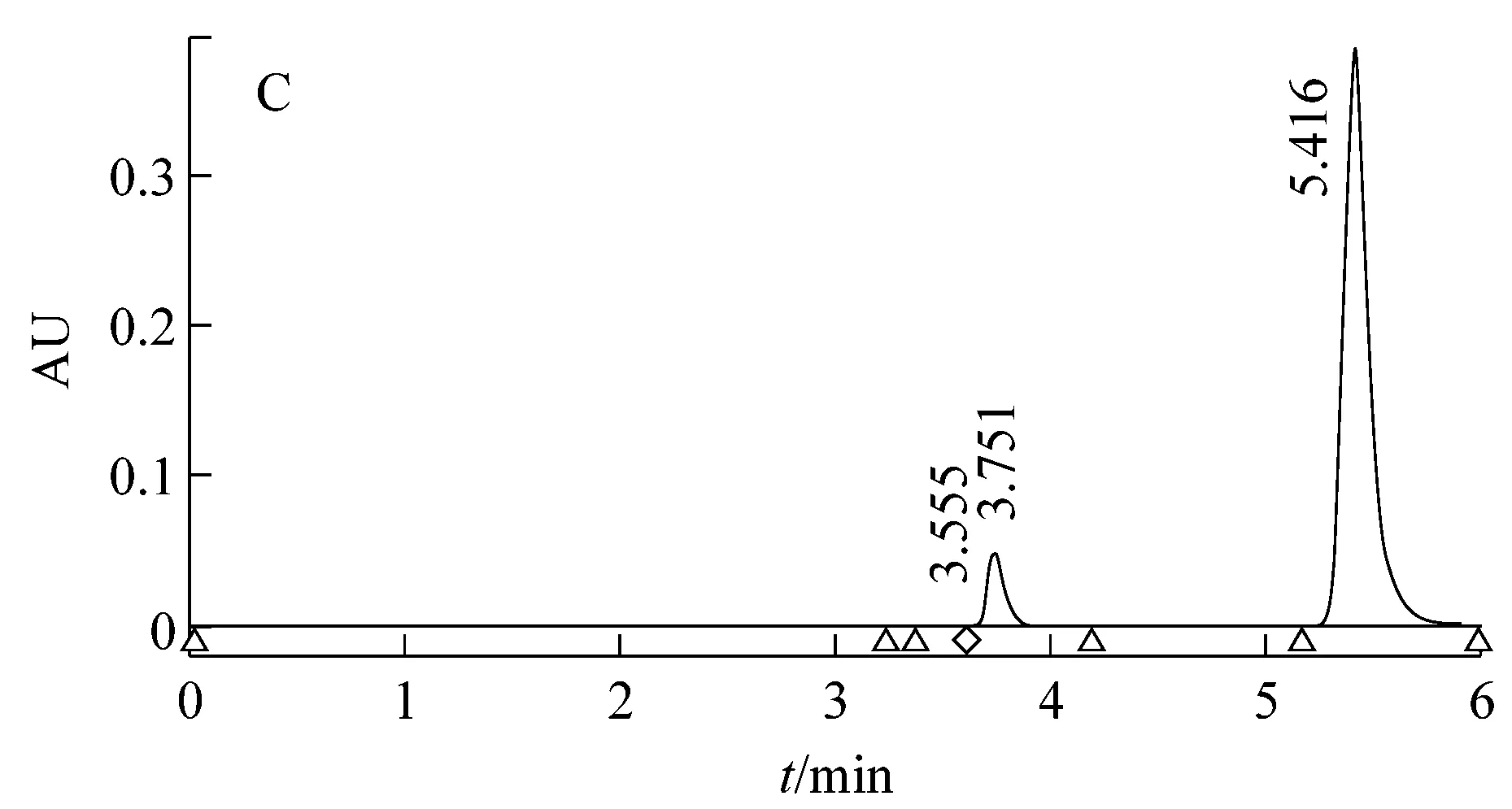
根据实验结果分析得出突变株AT39/btao反式茴脑代谢过程中茴香酸生成量为0.028 g/L,比野生菌AT39茴香酸产量下降了96.10%。且其互补菌HAT39/btao代谢过程茴香酸产量为0.843 g/L,比野生菌AT39增加了19.4%,说明对突变菌的回补效果较好。
本次实验中所用质粒为pBBR1MCS-5,所用的启动子为低水平表达的lac启动子,虽然相对野生型茴香酸的产量及茴香酸摩尔生成率均得到很大程度的提高,但为了后期能更好地应用于工业生产,可基于本次研究结果进行质粒强启动子的构建,使tao基因进行高水平表达从而获得更高产量的茴香酸。
3 结论
本研究对Pseudomonassp. AT39的tao基因进行了突变和表达,以研究该基因在菌株中对茴脑代谢及合成茴香酸的功能,发现tao基因的过量表达可以增加重组菌AT39/tao-pBBR1MCS5的酶活力,并提高茴香酸的转化能力。确定了tao基因是Pseudomonassp. AT39利用反式茴脑代谢生产茴香酸的重要基因。
