冬小麦叶片气体交换参数对水分胁迫的响应
2020-12-28张玉顺路振广张明智丁鹏飞王鸿铭杨浩晨邱新强
张玉顺,路振广,张明智,丁鹏飞,王鸿铭,杨浩晨,邱新强*
(1.河南省水利科学研究院/河南省节水灌溉工程技术研究中心,郑州 450003; 2.许昌市农田水利技术试验推广站,河南许昌 461000)
0 引言
【研究意义】河南省位于我国黄淮海平原,是国家粮食战略工程的核心区,同时也是严重缺水的省份。冬小麦是该地区的主要用水作物,其生育期内的降水量仅能满足生长所需水量的25%~40%[1-2],一般需要进行2~3次灌溉才能获得较高的产量。由于生产中按作物需水适时灌溉的操作难度极大,为避免冬小麦遭受过度的水分胁迫而影响正常生长,需要对作物的水分状况进行准确判别和及时掌握,从而指导灌溉。【研究进展】准确判别和测定作物水分状况是指导农田灌溉的基础[3],为使作物不受水分胁迫影响,应在作物过度受旱之前及时灌溉。叶片是作物进行光合作用和蒸腾作用的主要场所,作物通过气孔运动调节水分的蒸腾,同时维持植株水分和CO2利用的平衡,气孔导度降低往往在叶水势尚未发生变化之前就已经发生[4]。单叶水分利用效率(Leaf water use efficiency,WUE)是光合与蒸腾之比,反映了叶片气体交换过程中相关量的变化,而胞间CO2摩尔分数(Ci)的变化方向是确定光合速率变化的主要原因是否为气孔因素的重要判断依据[5]。土壤水分胁迫下,小麦叶片光合作用对土壤水分存在一个“阈值”反应,蒸腾速率则随土壤水分递增而一直递增[6],当叶片Ci从降低转为增高时,表明水分胁迫导致的光合速率降低的主要原因由气孔限制转变为非气孔限制[7-9]。【切入点】以往相关研究,多侧重于特定生育阶段或特定干旱程度下气体交换参数的动态监测和关键阈值分析[10-11],以及干旱过程对植株形态、耗水及产量指标的生理生态学响应[12-14]。已知特定时期适度水分亏缺不仅不降低作物的产量,反而能增加产量和提高水分利用效率[15-16],在生产中基于叶片气体交换参数间量化关系对作物自身所处土壤水分状况进行准确判别,从而确定最佳的灌溉时机及灌溉水量,实现按需适时适量灌溉,尚未引起过多关注。【拟解决的关键问题】基于此,拟选择当地主栽小麦品种,通过测坑控制试验对比分析不同水分胁迫条件下冬小麦叶片气体交换参数的差异,明确水分胁迫下叶片气孔行为的响应机制及其对产量的影响,以期为小麦节水高效灌溉提供参考。
1 材料与方法
1.1 试验区概况
试验在河南省节水灌溉工程技术研究中心(河南省灌溉试验中心站)毛庄试验基地(34.27°N,112.70°E,海拔85 m)内进行,属北温带大陆性季风气候。试验区年平均气温14.0~14.3℃,年平均降雨量640.9 mm,无霜期220 d,日照时间约2400h。土壤类型为中壤土,1 m 土层的田间质量持水率为23%,干密度为1.42g/cm3,地下水埋深大于5 m。耕层有机质量为5.62g/kg,全磷量为0.44g/kg,全钾量为15.12g/kg,全氮量为0.37g/kg,碱解氮量为24.91mg/kg,速效磷量为23.89mg/kg,速效钾量为75mg/kg。
1.2 试验设计
本试验在带有电动遮雨棚的大型非称重式蒸渗仪(L×W×H=3.3 m×2 m×2m)内进行。试验材料选择当地主栽的半冬性中熟品种“周麦22”,人工开沟撒播,每坑播9 行,行距20 cm,播量为150kg/hm2。本试验共2 季,分别于2012年10月17日和2013年10月12日播种,均于翌年5月30日收获,全生育期分别为225d 和230d。播前整地,基施复合肥750 kg/km2(N、P、K 质量比为15∶15∶15),干鸡粪10 t/hm2;播后灌蒙头水,灌溉水源为井水,水量由自动流量计控制。其余农事管理同一般高产田。
试验共设置18个水分处理,分别在苗期、拔节期、抽穗期和灌浆期各设置3个土壤水分控制下限(相对含水量)水平,同时设置生育前期(苗期和拔节期)连旱、生育中期(拔节期和抽穗期)连旱、生育后期(抽穗期和灌浆期)连旱,以及生育期适宜水分、生育期中水分、生育期低水分处理,以生育期适宜水分处理作为对照。试验的计划湿润层深度为1 m,当土壤含水率达到控制下限时即灌水90 mm,具体试验处理见表1[15]。为便于观测,对于需要进行干旱处理的测坑,在上一次灌水时根据其受旱程度适度减少灌水量,尽量使各处理的土壤水分同期达到其受旱下限,受旱处理复水后,其灌水量恢复到与适宜水分处理相同。
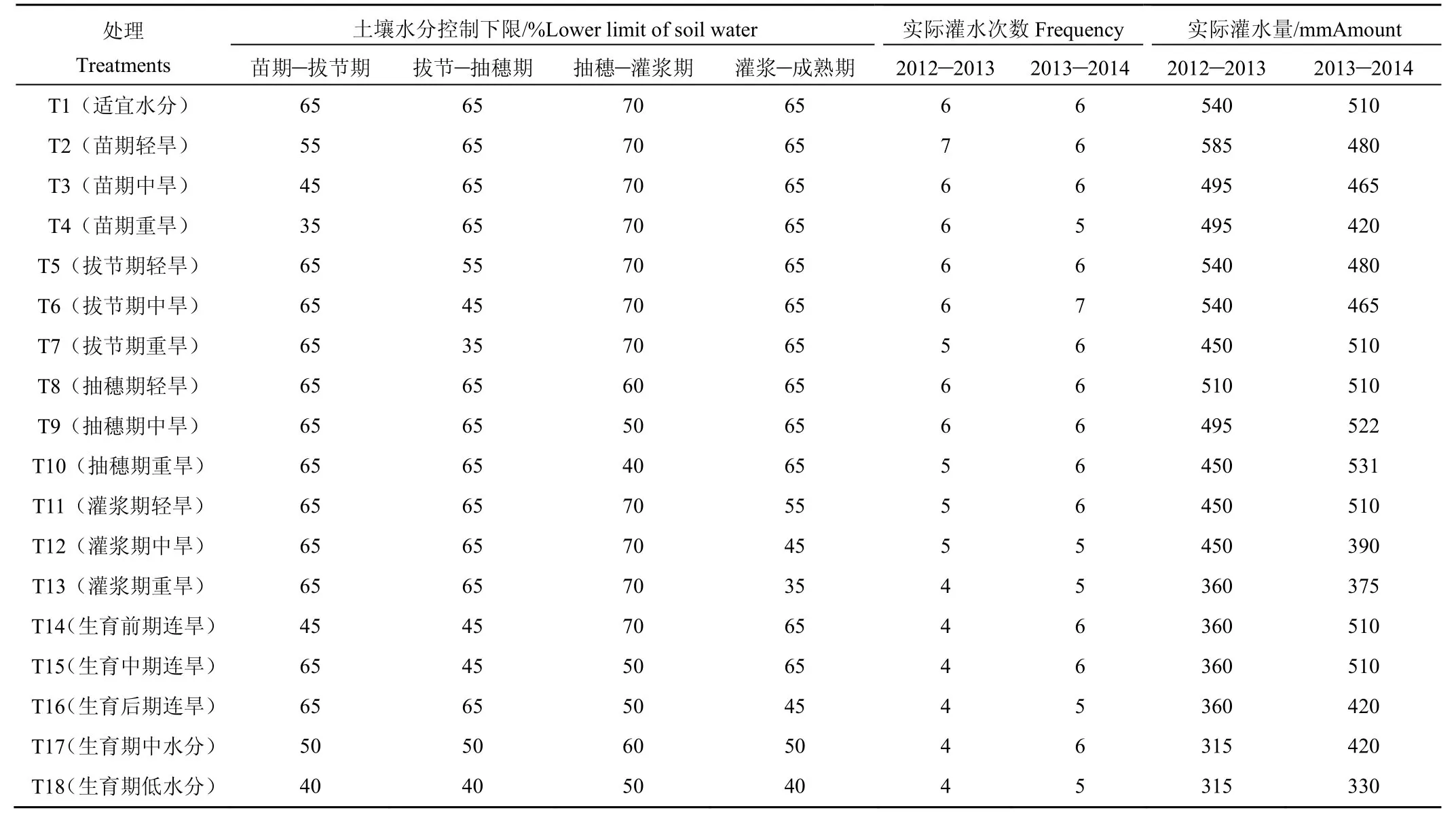
表1 试验处理Table1 Experimenttreatments
1.3 测定项目与方法
土壤含水率采用取土烘干法测定,测量深度为100cm,垂直方向每20cm为1层,每7~10 d 测1次,灌前、灌后均加测;作物实际耗水量由水量平衡公式得到,水分利用效率(WUE)由籽粒产量和耗水量相除得到;人工收割、脱粒、测产和考种,收获时对小区内中间3 行(共9 行)进行测产,同时选择小区1 m 行内有代表性的10株(10次重复)冬小麦进行考种(株高、茎粗、穗长、小穗数、无效小穗数、穗粒数和千粒质量等)。
选择晴好天气,在09:30—11:30 用Li-6400XT 便携式光合仪(美国LI-COR 公司)测定叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和Ci等指标,并计算叶片水分利用效率(WUE=Pn/Tr)和叶片气孔限制值(Stomatal limitation,Ls=1-Ci/Ca)。每处理选长势一致的3株进行测定(3次重复),挑旗前测最上一片全展叶,挑旗后测穗位叶。
利用Excel 2016 和SPSS 22.0 软件进行数据处理和分析,表中数据除特殊标注外均为所有重复的平均值。
2 结果与分析
2.1 苗期水分胁迫对冬小麦叶片气体交换的影响
2012—2014年苗期受旱各处理的Pn、Tr、Ci和Gs见表2。由表可知,拔节前(3月27日)冬小麦叶片的Pn、Tr、Ci和Gs均随干旱程度加剧而持续降低,其中T4 处理表现最弱,较对照(T1)处理分别降低35.47%、55.01%、16.51%和43.59%,与T2 处理差异显著。拔节期复水后,各受旱处理的气体交换参数均呈上升趋势,T2 处理和T3 处理的表现总体优于T1处理,T4 处理的表现则略弱于T1 处理。可见,苗期中度以上干旱在一定程度上弱化了冬小麦叶片的气体交换进程,不利于光合同化速率的提高。复水后苗期受旱各处理的气体交换参数均明显反弹,处理间差异变小,这在2012—2013年试验中同样有所体现,反映了本试验条件下的水分胁迫水平并未对其植株产生不可逆的生理胁迫。结合其主要农艺性状的表现不难看出[15],冬小麦苗期适当增加水分胁迫的程度不仅有利于节水,且能够达到“蹲苗”的目的。
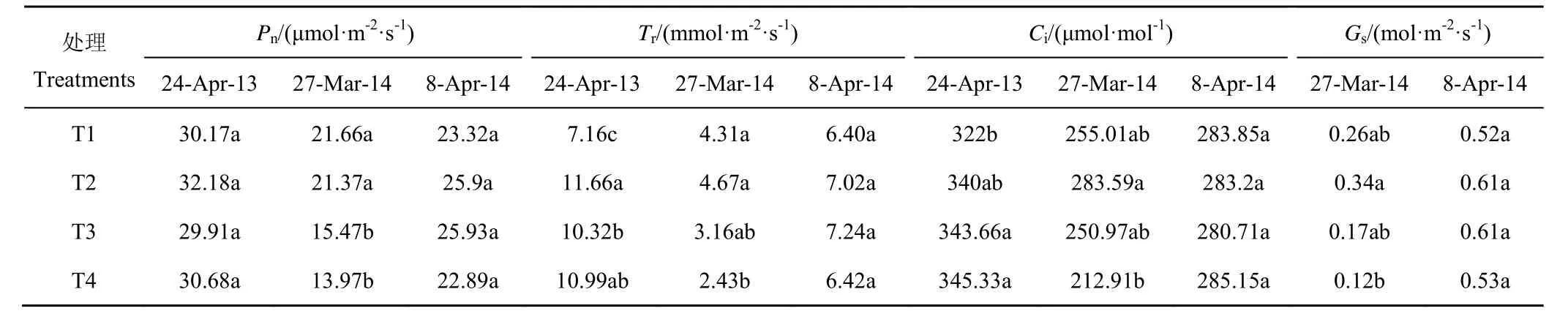
表2 苗期水分胁迫下冬小麦叶片的气体交换参数变化Table2 Changes of gas exchange parameters of winter wheat leaves under water stress at seedling stage
2.2 拔节期水分胁迫对冬小麦叶片气体交换的影响
2012—2014年拔节期受旱各处理的Pn、Tr、Ci和Gs见表3。2 季试验结果显示,冬小麦叶片的Pn、Tr和Ci在拔节—开花期总体呈先降低后上升的波状变化。2012—2013年上述各参数的峰值主要集中在挑旗后(4月24日);2013—2014年Pn和Tr均在拔节末期(4月14日)达到峰值,这可能与灌水(4月10日)有关,Ci的峰值则出现在挑旗后(4月27日),与上年度保持一致。
2012—2013年观测期内,拔节期受旱各处理的Pn较T1处理普遍降低,处理间极差值主要集中于3.31~4.67μmol/(m2·s)之间,其最大降幅(较对照)则分别达到7.08%(轻旱)、15.48%(中旱)和15.58%(重旱);2013—2014年度各处理间差异相对较小,处理间极差值不高于2.43 μmol/(m2·s)。受旱各处理复水前(4月1日)的Ci、Tr和Gs值较T1处理分别降低了约10.35%、20.17%和43.64%,复水后上述指标均快速恢复至正常水平(4月9日),T5、T6、T7处理Tr值较T1处理分别提高了16.54%、31.98%和46.79%,反映了气孔对土壤水分变化的快速响应。
2.3 抽穗期水分胁迫对冬小麦叶片气体交换的影响
2012—2014年抽穗期受旱各处理的Pn、Tr、Ci和Gs见表4。2012—2013年度,抽穗初期(4月24日)受旱各处理的Pn较T1处理均有所下降,其中T9和T10处理的降幅较大,但处理间差异均不显著。干旱持续至抽穗期末,各受旱处理的Pn、Tr和Gs间差异逐渐拉大,其中T10处理与T8处理间差异达显著水平。2013—2014年,T10处理的Pn在抽穗初期较T1处理降低约14.34%,但处理间差异不显著,T8和T9处理的叶片气体交换参数均未显著降低,其Tr较T1处理分别提高15.26%和18.50%。出现上述情况,应该与观测期内各处理间土壤水分梯度差异尚不明显有关,而随着水分胁迫的持续,抽穗末期各处理的Pn、Tr和Gs整体下行,Ci则呈上升趋势(重旱处理除外)。对比可知,T10处理的Pn和Tr明显低于其他受旱处理,其Ci和Gs则显著降低,较T1处理分别降低11.89%和31.18%。同期,T9处理Ci和Gs的降幅有限,与T1和T8处理间差异均不显著。2季试验结果显示,抽穗期重旱对冬小麦叶片的部分气体交换参数有显著负向影响,且上述各参数的变异性较大,群体内部的差异较为明显,这一现象体现了冬小麦为避免遭受更大损失相应的应对策略,亦或是品种特性的显现,有必要进一步研究验证。
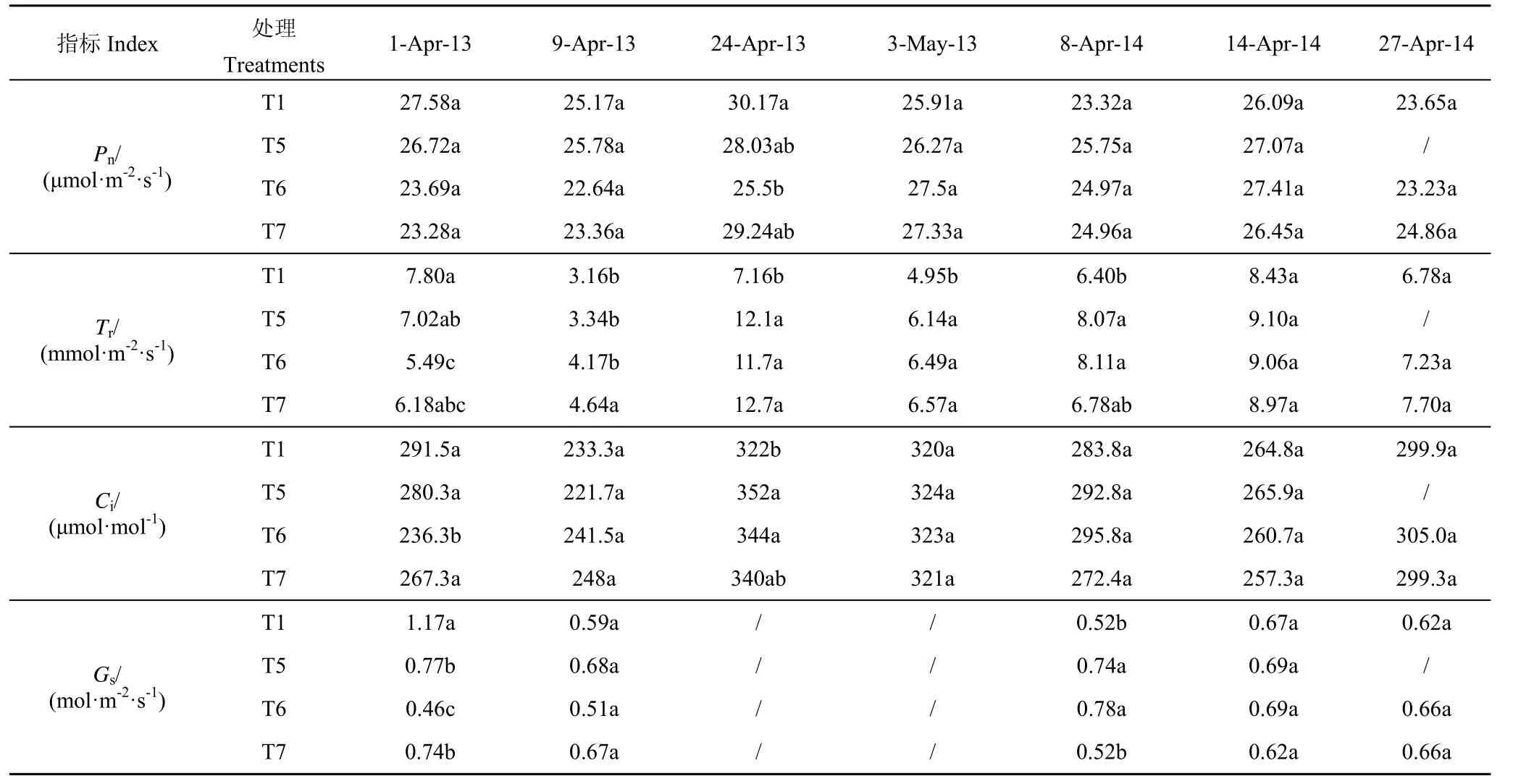
表3 拔节期水分胁迫下冬小麦叶片的气体交换特性Table3 Gas exchange characteristics of winter wheat leaves under water stress at jointing stage
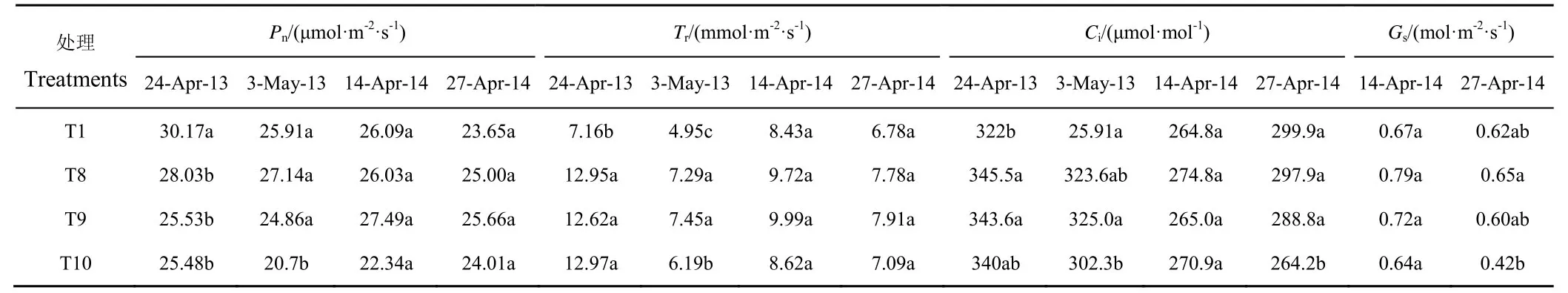
表4 抽穗期水分胁迫下冬小麦叶片的气体交换参数变化Table4 Changes of gas exchange parameters of winter wheat leaves under water stress at heading stage
2.4 灌浆期水分胁迫对冬小麦叶片气体交换的影响
2013—2014年灌浆期受旱各处理的Pn、Tr、Ci和Gs见表5。由表5可知,灌浆初期(5月8日)受旱,T13处理的Pn、Tr、Ci和Gs较T1处理分别显著降低了74.97%、73.25%、7.64%和87.17%,其他2个受旱处理的气体交换参数均未显著降低。随着水分胁迫的持续,灌浆中期(5月12日)各受旱处理的气体交换参数均显著降低,各受旱处理的平均降幅分别约为40.60%(Pn)、47.74%(Tr)、11.19%(Ci)和69.65%(Gs),同期重旱处理的Pn、Tr和Gs降幅总是最高。

表5 灌浆期水分胁迫下冬小麦叶片的气体交换参数变化Table5 Changes of gas exchange parameters of winter wheat leaves under water stress at the grouting stage
2.5 连续受旱对冬小麦叶片气体交换的影响
2012—2014年连续受旱各处理的Pn、Tr、Ci和Gs见表6。2 季试验结果显示,拔节期受旱期间T14和T15 处理的Pn均低于T1 处理,其中2012—2013年T15 处理的Pn最低,仅为T1 处理的51.97%(4月1日);2013—2014年T14 处理的Pn最低,较T1处理降低了约18.02%(4月8日)。同期,T14 处理的Tr、Ci和Gs分别降低了约19.60%、17.32%和50.43%,T15 处理的降幅更大,分别达到了45.29%、15.72%和78.92%。复水后T14 和T15 处理的气体交换参数均显著升高,其Pn、Ci和Gs均恢复至正常水平,Tr则“反弹”至T1 处理的1.92倍(2012—2013年度)、1.28倍(2013—2014年)。由表6可知,T16 处理与T1 处理间的差异自灌浆后开始凸显并随着生育进程的推延而逐步增大,至观测期末(5月12日)达到最大,T16 处理Pn、Tr、Ci和Gs分别降低了约16.90%、45.35%、25.16%和70.15%。
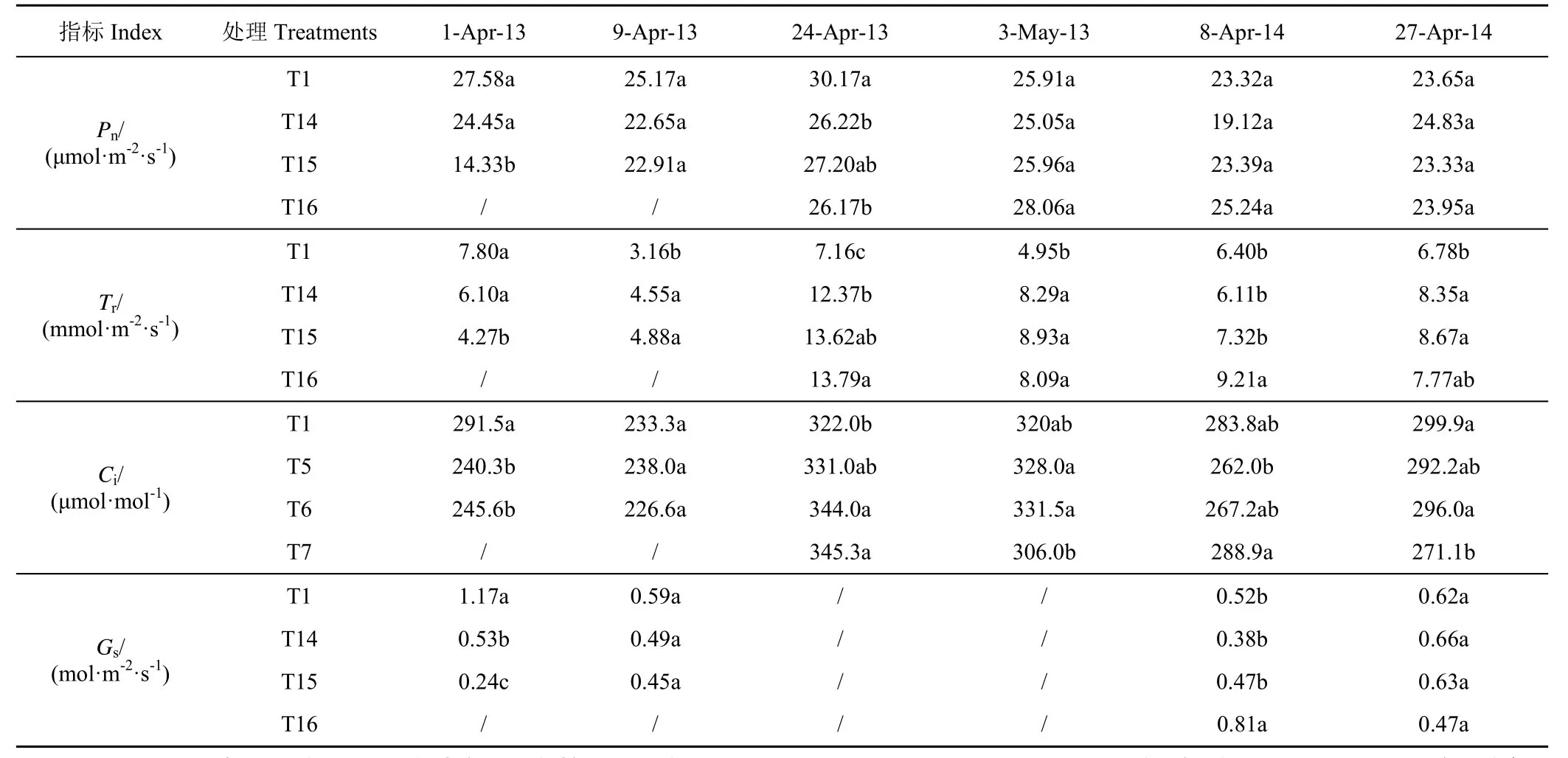
表6 连续受旱条件下冬小麦叶片的气体交换参数变化Table6 Changes of gas exchange parameters of winter wheat leaves under continuous drought conditions
2.6 全生育期受旱对冬小麦叶片气体交换的影响
2012—2014年生育期内受旱各处理的Pn、Tr、Ci和Gs见表7。随着水分胁迫程度的加剧,2 季观测期内冬小麦叶片的气体交换参数均值绝大多数呈下降变化,2012—2013年其最大降幅(即T17 处理较T1 处理)分别为25.87%(Pn)、-14.24%(Tr)、11.33%(Ci)和69.51%(Gs),相应地,2013—2014年则变为12.05%、24.12%、15.87%、38.68%。可见,长期持续受旱会明显抑制冬小麦叶片的气体交换进程,不利于光合同化产物的积累及运转分配,在生产中应优先杜绝出现此类现象。

表7 全生育期受旱条件下冬小麦叶片的气体交换参数变化Table7 Changes of gas exchange parameters of winter wheat leaves under drought conditions during the whole growth period
2.7 叶片水分利用效率(WUE)、气孔限制值(Ls)与主要气体交换参数间的关系
对WUE、Ls与Pn、Tr、Gs等主要气体交换参数进行回归分析,结果见表8。由表8可知,Ls与Pn呈二次曲线关系,与Gs、Tr呈线性关系,其中作物叶片的气孔限制值主要集中在0.3 以下;WUE与Pn、Tr呈二次曲线关系,与Gs呈非线性Ellipse曲线关系。已有研究普遍认为,当作物遭受水分胁迫时Tr即出现下降,且随着胁迫程度加剧而持续降低;随着WUE的升高,当Gs开始降低时,说明作物正在遭受中度水分胁迫阶段;当WUE达到最高后,作物将遭受重度水分胁迫,此时的Gs已显著降低,下一步WUE将明显下降,非气孔限制成为叶片气体交换的主要限制因子[7-9]。本试验中,随着WUE的提高,Pn和Tr均呈先升高后降低的趋势,其中Tr极值的出现时间要早于Pn。当WUE在2.62 μmol/mmol 左右时Tr达到极值,此时作物叶片已感知到土壤水分胁迫的反馈;随着水分胁迫的加剧,Gs持续降低,当WUE出现极值,即WUE在3.36 μmol/mmol 左右时,作物已开始由轻度水分胁迫转入中度水分胁迫阶;当WUE在3.81 μmol/mmol 左右时,Pn达到极值并开始下降,但WUE仍将保持上升态势,因为当前Pn的降幅仍小于Tr的降幅。通过联立方程计算可知,当WUE在4.11 μmol/mmol 左右时,Pn的降速开始低于Tr,此时叶片的WUE达到极值,Gs也较低,说明作物继续受旱的话将遭受重度水分胁迫。
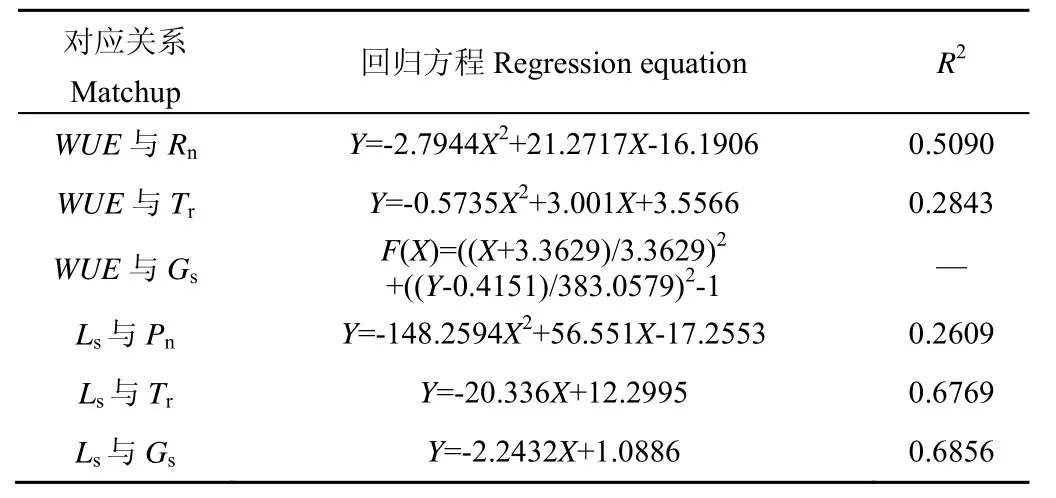
表8 叶片水分利用效率、气孔限制值与主要气体 交换参数间的回归分析Table8 Regression analysis results between WUE,stomatal limit value and main gas exchange parameters of leaves
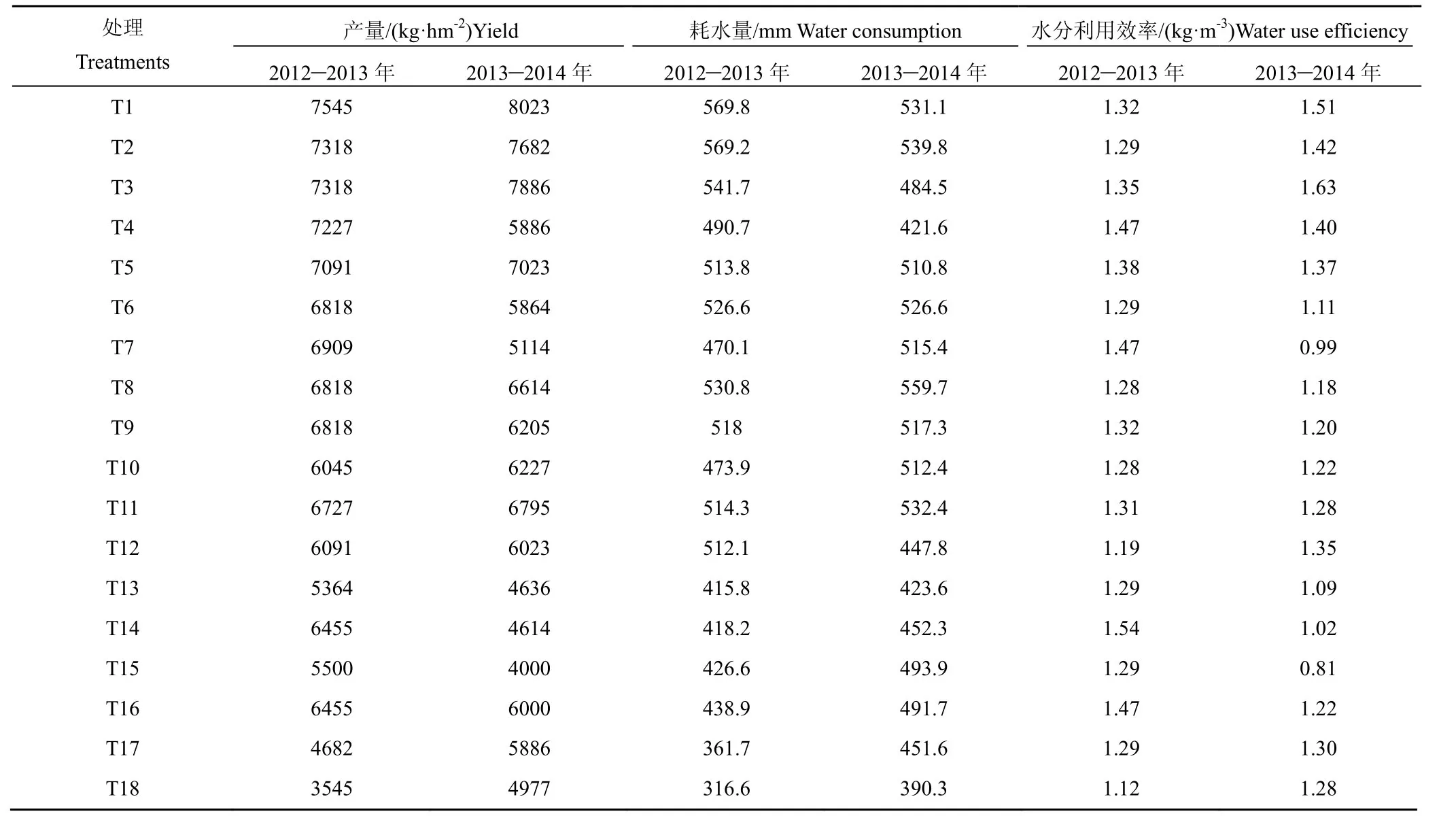
表9 冬小麦的籽粒产量、耗水量和水分利用效率Table9 Grain yield,water consumption and water use efficiency of winter wheat
2.8 冬小麦的籽粒产量、耗水量和水分利用效率
由表9可知,任一生育时期受旱均限制了冬小麦全生育期总耗水量的增加,且随干旱程度加剧呈下降趋势,各重旱处理的总耗水量总是最低。2012—2013年冬小麦的平均减产率和耗水量降幅分别约为7.38%和6.63%(轻旱)、10.39%和7.94%(中旱)、15.36%和31.87%(重旱);2013—2014年分别约为12.39%和-0.86%、19.05%和6.97%、31.87%和11.83%。全生育期干旱条件下,T18 处理较T1 处理下降约15.15%(2012—2013年)和15.57%(2013—2014年);T2和T3 处理的籽粒产量降幅在5%以内,其WUE则普遍较高,说明苗期适度干旱对提高冬小麦的水分利用效率有利,且并不会导致明显减产;抽穗期和灌浆期受旱各处理的水分利用效率较T1 处理分别下降约0.29%~3.36%和0.91%~9.89%(2012—2013年)、19.55%~21.78%和10.96%~27.54%(2013—2014年);灌浆期受旱各处理的籽粒产量和总耗水量均随干旱程度加剧呈下降变化,且其籽粒产量的降幅总是较大,并最终导致其水分利用效率低于T1 处理;连旱处理条件下,冬小麦的籽粒产量和总耗水量均明显降低,T15 处理的水分利用效率最低。尽管各处理的产量表现与上述气体交换参数的测定结果总体吻合,且相关分析结果显示不同生育阶段内各气体交换参数与产量间存在一定的相关关系(表10),但由于总体显著性不高,且存在明显的年季间差异,因此仅能通过气体交换参数来对当前的土壤水分状况进行准确判断,并不能对最终的籽粒产量水平进行准确估算。

表10 冬小麦叶片主要气体交换参数与籽粒产量间相关系数Table10 Correlation coefficient between main gas exchange parameters of winter wheat leaves and grain yield
3 讨论
气孔是植物在长期进化过程中所特有的适应陆生干旱环境的一种结构,气孔运动对植物体具有十分重要的生理意义。在正常气候条件下,气孔的开闭主要受光照和CO2调控,但在干旱条件下,水分条件就成为决定气孔开闭的主要因素。前期轻度水分胁迫可保证较高的小麦叶片光合速率,且有助于水分利用效率提高,重度亏缺则会产生显著的后效性影响[17],这一情况在玉米生产中同样有所体现[18]。本试验中,苗期受旱期间冬小麦叶片的气体交换参数均随干旱程度加剧而持续降低,拔节期复水后明显回升,甚至有所提高,且苗期轻旱和中旱处理的籽粒产量均未显著降低。可见,冬小麦苗期适当加剧土壤水分胁迫不仅有利于节水,且能够实现“蹲苗”的目的。刘琳等[19]指出拔节期水分胁迫-复水后石家庄8号的各项生理指标均快速补偿,轻度水分胁迫-复水后出现超补偿效应,提高了水分利用效率,本试验也得到了相似的结果。随着水分胁迫程度的加剧,灌浆期受旱各处理的Pn、Tr和Gs均显著降低,其中重旱处理的Ci值明显升高,这一现象在生育期低水分处理上同样有所体现,此时非气孔因素已成为叶片光合作用的主要限制因素。可见,长期持续受旱会明显抑制冬小麦叶片的气体交换进程,不利于光合同化产物的积累及运转分配,在生产中应优先杜绝出现此类现象。已有研究已基于节水高产目标提出了冬小麦不同生育阶段的适宜灌水控制下限标准[15],上述结果可作为已有研究成果的进一步深化和补充。
在土壤水分胁迫条件下,植物气孔调节机理被认为是一种反馈式调节[20]。当土壤水分充足时,随着光合有效辐射强度、大气温度的增高,气孔导度随之升高;当水分胁迫达到某一临界值时,植物通过叶片蒸腾剧烈失水,导致叶片水势产生变化,从而造成叶片气孔开度减小[8,17,21]。轻度水分胁迫下,光合速率没有下降甚至高于非胁迫处理,而由于蒸腾速率先于光合速率下降使得WUE提高;中度水分胁迫下,气孔开度明显下降,蒸腾速率的降幅大于光合速率,此时WUE最高;在严重缺水时,叶肉细胞光合活性的降低,导致了光合作用的气孔限制和非气孔限制同时存在,光合速率明显下降,WUE也显著降低。在大多数禾谷类作物中,当植株的水分状况并未受到土壤干旱的影响时,气孔即开始关闭,表现为调亏期内蒸腾作用下降超前于光合作用[22]。本试验也得到了上述结果,并基于WUE与Pn、Tr、Gs之间的相关关系,对周麦22所处水分胁迫程度进行了量化表征,即随着干旱程度的加剧,当WUE值分别在2.62、3.36 和4.11 μmol/mmol 左右时,周麦22 分别开始遭受轻度、中度和重度水分胁迫,当WUE值在3.81 μmol/mmol 时,Pn达到极值。上述结果从气孔运动的视角给出了作物所处水分状况的判定标准,为能够在生产中实现按需适时灌溉提供了简单快捷的评判思路,具有一定的实践意义。当然,明确不同生育时期内WUE与各气体交换参数间的定量关系,分阶段建立作物WUE值控制目标,对于实现小麦节水和高产的统一[23],将更具实践意义。此外,由于叶片的气体交换参数具有明显的日变化过程,而本试验的数据均是在上午获取的,该时段的气体交换过程相对活跃,但受气孔开度的影响较大,会在一定程度上影响试验成果的可靠性,因此在获取数据时应尽量选择晴好天气。
4 结论
1)拔节期复水后,苗期受旱处理的气体交换参数均明显回升,其中苗期轻旱和中旱处理均未显著减产,可见苗期适当增加水分胁迫的程度是可行的。考虑到拔节后重旱对冬小麦叶片的部分气体交换参数有显著负影响,从节水稳产角度出发,本试验建议冬小麦拔节后的土壤水分不应低于田持的55%。
2)基于WUE与Pn、Tr、Gs之间的相关关系,确定了周麦22 开始遭受轻度、中度和重度水分胁迫时叶片水分利用效率的理论计算值分别在2.62、3.36和4.11 μmol/mmol左右,当WUE值在3.81 μmol/mmol时,Pn达到极值,上述结果可作为准确判别作物自身所处土壤水分状况的数据参考。
