低磷胁迫对水稻生长及生理特性的影响*
2020-12-21朱兰保常晓梅
丁 艳,朱兰保,盛 蒂,常晓梅
(蚌埠学院土木与水利水电工程学院,安徽 蚌埠,233000)
水稻是重要粮食作物之一,全球约有50%以上的人口以稻米为主食[1]。水稻的生长过程中,磷肥是不可缺少的营养元素,磷可增强植株体内活力,促进养分合成和转运,增强光合作用,延长叶的功能期,有益于水稻增产稳产[2,3]。磷素供应不足会影响水稻的正常生长发育,进而影响其产量和品质[4,5]。
有研究表明,在低磷条件下,作物会从形态和生理等方面做出一些适应性反应,以适应低磷环境[6~8]。刘文菊等[9]研究发现,不同基因型水稻在磷胁迫下其根系形态发生了明显变化,如根长、根冠比均呈增加趋势。在低磷条件下,水稻植株分蘖数增减少,且每株根系相互之间有横向交织生长现象[10]。推测水稻在低磷处理下根系可能会通过横向生长来扩大与培养液的接触面积,从而增加对磷的吸收。此外,磷胁迫下水稻可通过提高根系APase 活性增强对磷胁迫环境的适应性[11]。综上所述,有关水稻对低磷胁迫的适应机制的研究,大多集中在苗期根系的适应性机制,针对水稻地上部组织在形态和生理等方面对缺磷的适应性反应的报道较少。本次研究是在前期研究的基础上,选取3种不同基因型水稻为研究对象,采用全营养液和低磷营养液培养水稻幼苗,以植株表型、根系形态、生物量及根系分泌物(APaes,H+)等为参数,分析低磷胁迫对水稻生长的影响,以及水稻对低磷的适应性反应,旨在为进一步从分子水平上阐明水稻对低磷胁迫的响应机制提供理论依据。
1 材料与方法
1.1 供试材料
选用郑旱6号(ZH6,耐低磷基因型)、通粳981(TJ981,磷敏感基因型)和镇稻99(ZD99,根系扩展型)3种不同基因型水稻品种为试验材料,供试水稻种子由江苏润扬种业股份有限公司提供。
1.2 试验设计
选择籽粒饱满的水稻种子,浸入100 mL/L H2O2溶液中消毒30 min,用去离子水反复冲洗后,放入盛有适量去离子水的培养皿(17 cm)中,催芽。待种子露白时,挑选发芽势一致的种子放入96孔板置于塑料容器内,采用国际水稻研究所(International Rice Research Institute, IRRI)完全培养液培养[12]。当幼苗长至3叶期时,挑选长势健壮的幼苗进行分组培养,即正常营养液培养(记为:+P)和低磷营养液培养(记为:-P),且c(+P)∶c(-P)= 25∶1。每个处理选择4株生长健壮且长势基本一致的幼苗,每个处理设置10个生物学重复,用泡沫板固定后置于12 L黑色塑料桶中,放置于塑料大棚中。自然光照和温度条件下,分别用正常供磷和缺磷培养液进行全生育期培养。幼苗生长期,每3 d更换1次培养液,拔节至成熟期,每5 d更换1次培养液。
1.3 测定项目与方法
1.3.1根系形态扫描 分别挑选正常供磷和低磷处理的水稻植株各20株,用去离子水反复冲洗根部后,利用植物根系扫描与分析系统(型号:WinRHIZO,生产商:上海临嘉科教仪器有限公司生产)扫描根系并进行分析。
1.3.2生物量、叶绿素含量及根系活力的测定 选择低磷处理15 d的水稻植株,每个处理分别取20株长势一致的幼苗,用去离子水反复冲洗根系和叶片表面,吸水纸吸干其表面水分,称其地上部和根部的鲜质量,后置于烘箱中,105 ℃杀青30 min,70 ℃烘干至恒质量,再次称量地上部和根部的质量。采用差减法计算单株相对含水率,地上部(或根部)干物质质量及根冠比等参数。采用分光光度法测定叶片中叶绿素的质量分数;TTC还原法测定根系活力。
1.3.3根系分泌酸性磷酸酶活性的测定 参考文献[6]的方法进行,略有改动。取生长健壮的水稻幼苗2株,用去离子水冲洗干净根系,置于含有100 mL摩尔浓度为1 mmol/L对硝基酚磷酸二钠(p-NPP)培养液的锥形瓶中,锥形瓶用黑色薄膜包裹,正常光照培养2 h后,吸取1 mL反应液加入到含有5 mL摩尔浓度为1 mol/L NaOH的试管中,摇匀。以无酶反应为空白对照,450 nm处测定吸光度值。根系分泌酸性磷酸酶活性以单位时间单位质量鲜根水解p-NPP生成p-NP的量来表示(单位:mg·(h·g)-1)。
1.3.4根际或根表pH值的显色观察-琼脂糖凝胶法 称取1.5 g琼脂糖分别溶于150 mL正常供磷(+P)和缺磷(-P)培养液(pH 5.4)中,于微波炉中溶解,取出后冷却至40~50 ℃时,加入甲基红和溴甲酚绿混合酸-碱指示剂,使混合酸-碱指示剂的终质量浓度为0.06 g/L,继续冷却至室温,并分装于25 mL玻璃试管中。分别选择萌发后(带胚乳)和低磷处理3 d后(去胚乳)长势一致的幼苗,植入装有10 g/L琼脂糖的试管中,自然光温条件下生长,分别在其生长24,48,72 h后观察琼脂糖的颜色变化[13]。
2 结果与分析
2.1 低磷对水稻植株表型和根系形态的影响
与正常供磷水稻相比,磷素供应不足导致水稻植株表现出显著的表型差异(图1(A))。主要表现为:地上部生长缓慢,植株矮小,叶面积减小;分蘖期延迟且分蘖数减少。此外,低磷导致根系形态发生变化:磷敏感型品种TJ981根长增加明显(1.37倍),且增长幅度高于耐低磷品种ZH6(1.15倍)和磷高效吸收型品种ZD99(1.31倍);而ZH6和ZD99根平均直径(1.13 倍和1.25倍)和根表面积(1.45倍和1.82倍)增加幅度均大于TJ981(1.07倍和0.83倍)(图2)。
低磷处理的不同基因型水稻,其主根长度均较正常供磷水稻主根长明显增长,尤其磷敏感型水稻品种TJ981主根长度增长较显著(1.67倍),其次是ZD99(1.57倍),而耐低磷水稻品种ZH6主根长增幅较小(1.41倍)(图3)。由此可见,磷胁迫导致水稻根系增长,且磷敏感基因型水稻TJ981根系增长幅度显著高于ZD99和ZH6。也就是说,根系形态变化是水稻对低磷胁迫的一种适应性响应,即低磷时水稻通过改变根系形态,增加与土壤的接触面积,从而提高其对土壤中磷的吸收和利用。
与对照相比,低磷处理使3种不同基因型水稻分蘖期叶片的长度和宽度均呈减小趋势(图3)。其中,低磷处理叶片与对照叶片长度比率分别为ZH6(0.899),TJ981(0.929),ZD99(0.799);叶片宽度比率的变化表现为ZH6(0.922),TJ981(0.843), ZD99(0.810)。本次研究结果可知,低磷胁迫导致水稻开花期较正常供磷水稻延迟,穗形偏小且每株穗数和每穗粒数减少(图1(B))。这是由于磷素匮乏致使水稻植株叶片的长度和宽度均减小,光合速率下降,光合产物积累量减少,进而影响水稻的产量。
2.2 低磷对水稻幼苗相对含水率和干物质量的影响
本次研究结果显示,缺磷水稻幼苗相对含水率下降,以ZH6植株含水率下降较大(图4(A));根部干质量增加(图4(B)),与TJ981和ZD99相比,ZH6根部干质量增加较多。3个不同基因型水稻植株根冠比变化由大到小依次为:ZH6,ZD99,TJ981(图4(D))。这是由于低磷导致水稻幼苗地上部生长受阻(图4(C)),而根系为了增加与土壤的接触面获取更多的营养,从而改变其形态。
2.3 低磷对水稻叶片叶绿素质量分数的影响
低磷条件下,水稻植株分蘖期叶片叶绿素质量分数较对照发生变化,其中TJ981和ZH6叶片叶绿素质量分数增加,而ZD99叶片中叶绿素质量分数呈下降趋势(图5(A));进入孕穗期以后,与分蘖期相比较,ZH6和TJ981植株叶片的叶绿素质量分数均下降,ZD99变化不明显;与对照相比,3种不同基因型水稻叶绿素质量分数均表现为下降(图5(B))。可见,随着低磷处理时间的延长,水稻叶片中叶绿素质量分数降低。

图1 低磷处理条件下,不同基因型水稻幼苗表型(A)分蘖期;(B)抽穗、灌浆期

图2 低磷处理2周后水稻根系形态指标的变化
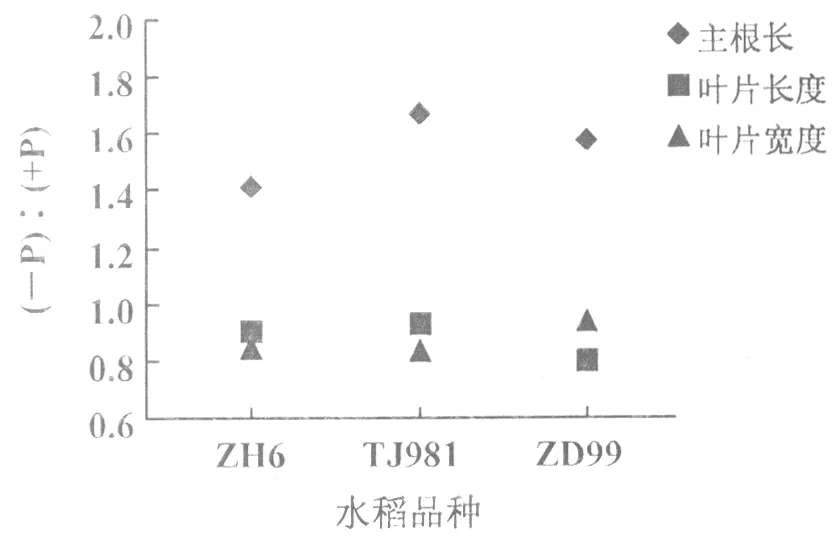
图3 水稻分蘖期的主根长及叶片变化
2.4 低磷处理对水稻幼苗根系活力的影响
低磷处理不同基因型水稻幼苗15 d后,其根系活力均呈下降的趋势(图6)。即低磷胁迫下磷敏感型品种TJ981,根系扩展型ZD99和耐低磷品种ZH6根系活力有不同程度的下降,其中ZH6和ZD99根系活力呈显著下降(p<0.05),根系活力下降率分别为42.01%和19.70%,而TJ981根系活力呈极显著下降(p<0.01)。这表明低磷胁迫影响水稻根系的活力水平,且TJ981较其它2种基因型水稻根系活力下降幅度大(下降率为51.48%),可见磷敏感型水稻TJ981对低磷的适应能力弱。
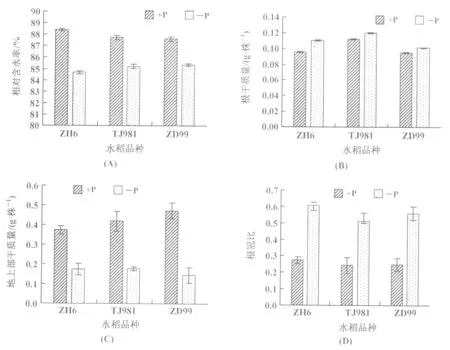
图4 低磷处理(3周)水稻植株生物量的变化(A):相对含水率;(B):根部干质量;(C):地上部干质量;(D):根冠比
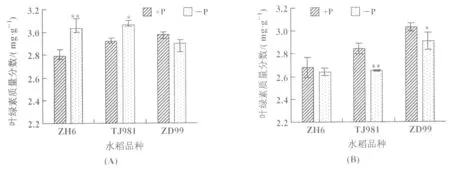
图5 叶绿素质量分数的变化(A):分蘖期;(B):孕穗期注:*和**分别代表p<0.05和p<0.01,下同。
2.5 低磷处理对水稻幼苗根系分泌酸性磷酸酶(APase)活性的影响
低磷处理3种不同基因型水稻幼苗1,8,15 d后,其根系分泌的酸性磷酸酶活性均高于对照,但不同基因型水稻幼苗根系分泌的磷酸酶活性增长表现不一(图7)。其中:耐低磷品种ZH6低磷处理1 d时,根系分泌的酸性磷酸酶活性较对照增强52.7%;低磷处理8 d和15 d时,其活性增长趋于稳定,增幅分别为9.1%和9.7%。ZD99根系分泌酸性磷酸酶的活性随处理时间的延长而增强,分别为5.5%,26.0%和40.1%;而磷敏感性品种TJ981随低磷处理时间的延长,其根系分泌酸性磷酸酶的活性增强相对稳定,增幅分别为12.5%,11.0%和11.6%(图7)。可见低磷胁迫使不同基因型水稻根系分泌酸性磷酸酶活性增强。

图6 低磷条件下水稻根系活力的变化

图7 低磷条件下水稻根系APase活性的变化
2.6 低磷处理对水稻幼苗根际或根表pH的影响
本次试验以耐低磷品种ZH6为例,采用10 g/L 琼脂糖凝胶观察根系分泌酸的变化。主要原理是:甲基红-溴甲酚绿酸碱混合指示剂的变色点为 5.1,pH > 5.1 呈绿色,pH < 5.1 呈酒红色。与对照相比,缺磷水稻幼苗根系分泌酸使琼脂糖变红色,而且随着处理时间(24,48,72 h)的延长,琼脂糖的红色加深,这表明磷胁迫促进了根系分泌酸的增多(图8)。同时,在幼苗移植时采用带胚乳(图8(A))和去胚乳(图8(B))2种情况进行对比发现,同样低磷处理48 h,移植时带胚乳的幼苗根系比去胚乳的幼苗根系分泌的酸量多,即带胚乳幼苗根系分泌酸使琼脂糖呈较深的红色。这表明幼苗生长前期胚乳可以提供部分营养供其生长发育所需,在没有胚乳营养的条件下,幼苗生长力减弱,分泌酸减少。但相比于对照,低磷处理的根系分泌酸量增加,琼脂糖红色明显。
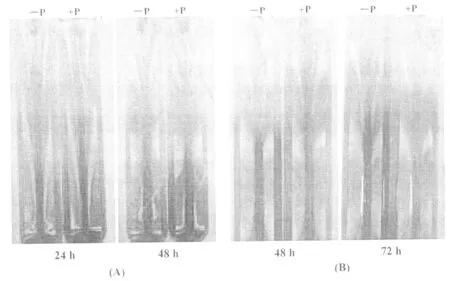
图8 低磷对ZH6幼苗根系分泌酸的影响(A)萌发后带有胚乳植入;(B):萌发后去胚乳植入
3 讨论与结论
水稻是对磷营养较敏感的作物。低磷胁迫下,为了适应逆境条件水稻会产生一系列的适应性响应。黄荣等[14]研究表明,缺磷会影响植物的生长发育且产生相应的症状,如植株生长迟缓、矮小、分枝数减少。本次试验结果表明,低磷胁迫会使水稻植株矮小、叶小、分枝、分蘖减少,这是由于低磷影响植株体内核蛋白的形成,从而抑制细胞分裂与增殖,导致植株生长发育延缓[15]。根系是最先感受养分胁迫信号并对这一逆境信号加工、处理和传递[4],其形态变化是水稻适应低磷胁迫的机制之一,如主根增长,侧根数目增多,根表面积增大[16]。本次试验结果显示,低磷处理水稻幼苗2周后,耐低磷水稻品种郑旱6号在低磷条件下其根系长度变化不明显,但其平均直径和根面积均呈增加趋势;而磷敏感型品种通粳981根系增长、根平均直径和根面积减小,根系呈细长型(图2)。因此,由于缺磷改变根系形态并使其根质量增加,从而促进根冠比增大。此外,低磷条件下,分蘖期水稻根长显著增长。郑旱6号,通粳981和镇稻99的根长增幅分别为对照的1.4,1.7和1.6倍。可见低磷胁迫时水稻可以通过增加根系长度,增大根表面积等,扩大与土壤的接触面积,进而增加对磷素的吸收和利用效率,满足自身生长所需的磷营养[17]。
低磷胁迫会诱导植物产生一系列的生理生化反应以调节植物的生长发育。本次试验结果显示,低磷处理水稻幼苗,使其叶绿素的质量分数在分蘖期和孕穗期表现不一。其中,郑旱6号和通粳981叶片中叶绿素的质量分数先增后减,镇稻99叶绿素的质量分数在分蘖期和孕穗期均减少。这是因为随着低磷处理水稻时间的延长,低磷限制了叶绿素的合成使其含量降低。此外,低磷水稻抽穗期延迟,且每株穗数及穗粒数减少,这是由于低磷水稻叶片中叶绿素质量分数的减少,减弱了水稻植株的光合能力,影响光合产物的形成和积累,进一步降低了水稻的产量。
在生理学上,植物可以通过根分泌酸性磷酸酶(APase)活性的提高和小分子量有机酸的增加,从而减小根际的pH。APase是一种诱导酶,当磷是植物生长的主要限制因子时,植物根系就向胞外分泌酸性磷酸酶,增加对土壤有机磷的水解[6],其活性受植物供磷状况的影响,低磷能诱导根系APase活性显著升高[11]。本次试验结果也显示,低磷处理3种不同基因型水稻根系分泌酸性磷酸酶活性均有不同程度的提高。根际酸化是植物,特别是双子叶植物,应对低磷胁迫的一种响应。本次研究发现,低磷促使水稻根系分泌酸增多,根系分泌的有机酸与难溶性磷酸盐进行阴离子交换或竞争性吸附,以释放或溶解难溶性磷酸盐,从而提高磷的利用率。
