切除眼柄与触角对波纹龙虾存活、蜕壳、摄食和生长的影响
2020-12-17卓宏标吴佳俊梁华芳罗嘉俊温崇庆
卓宏标,吴佳俊,梁华芳,罗嘉俊,温崇庆
(广东海洋大学 水产学院,甲壳动物遗传育种与增养殖实验室,广东 湛江 524088)
在印度-西太平洋地区分布的10种龙虾属种类当中,有8种在中国东南沿海分布[1]。由于国内龙虾市场需求强劲,中国大部分龙虾都是由国外进口,中国是当前世界首屈一指的龙虾消费大国,因此,龙虾是继对虾之后又一富有市场潜力的开发品种。近年来,波纹龙虾Panulirushomarus作为新兴养殖品种越来越受到青睐,在越南和印尼等地,野生波纹龙虾幼苗丰富且易捕捉,在当地已形成一套成熟的网箱养殖模式[2-3]。目前,国内已开展了波纹龙虾的网箱与工厂化养殖[4-5]、人工繁育[6-8]和养殖生物学研究[9-11],为龙虾养殖业的发展奠定了丰富的理论基础。
眼柄中X-器官窦腺复合体是甲壳动物多种激素的合成和分泌源[12],分泌多种激素,如性腺抑制激素和蜕皮抑制激素。眼柄微结构观察显示,其生长纹的数目与虾蟹类的生长有密切关系[13]。切除眼柄被证实能有效诱导卵巢成熟和发育,最初由国外学者Panouse[14]发现,随后便相继应用于对虾和龙虾卵巢促熟[15-16]。但切除眼柄对虾蟹的生长影响研究尚较少,陈政强等[17]研究了切除单侧眼柄对中国龙虾Panulirusstimpsoni的影响, 有关波纹龙虾眼柄切除的研究目前尚未见报道。
甲壳类动物的自切行为是一种常见高效的反射性反应,由种间因素、高养殖密度、人为因素等所导致[18],附肢自切损伤与再生对虾蟹类成活率和生长的影响在国内外已被广泛研究,如杂色龙虾Panulirusversicolor[19]、中华绒螯蟹Eriocheirsinensis[20-21]、三疣梭子蟹Portunustrituberculatus[22]等。第二触角是波纹龙虾区别于其他甲壳动物的显著特征,在养殖生产中因捕捉龙虾操作不当而导致其挣脱并失去第二触角的情况时有发生。目前,国内对虾蟹类触角的研究仅限于形态学观察[23-25],第二触角的缺失对波纹龙虾养殖性能的影响尚不清楚。为此,本研究中探讨了眼柄和第二触角切除对波纹龙虾幼虾存活、蜕壳、摄食和生长的影响,以期完善波纹龙虾的人工养殖技术,并为今后深入开展相关研究提供参考。
1 材料与方法
1.1 材料
试验用波纹龙虾于2019年4月购自海南省琼海市长坡镇,为自然海区捕获的不同规格的幼龙虾。采用干运法运回广东海洋大学东海岛生物研究基地,暂养于20 m3的水泥池中,用黑幕布遮光,24 h 连续充气。试验海水源自自然海区,经沉淀、沙滤,盐度为26~30,pH为 8.1~8.3,日换水量为1/3~1/2,每天投喂一次菲律宾蛤仔Ruditapesphilippinarum、杂色蛤仔Ruditapesvariegata等鲜活饵料,驯养15~20 d后,挑选健康活泼、规格合适的个体开展正式试验。
1.2 方法
1.2.1 蜕壳期的判定 为明确试验用波纹龙虾所处的蜕壳期,剪取少量龙虾的腹肢置于载玻片上,并于显微镜(×10)下观察刚毛的形态特征,参照Radhakrishnan[26]对蜕壳期的划分标准,将龙虾按蜕壳前期、蜕壳间期和蜕壳后期区分开来。
1.2.2 试验设计及日常管理 选取体质量为(18.15±1.85)g、大小相近、健康无损伤的波纹龙虾135尾,随机分为3组,分别为对照组、眼柄切除组和触角切除组,每组设3个平行,每个平行15尾虾,放养于体积为0.5 m3的塑料圆桶中,所有试验桶内的养殖条件均一致。对照组不做任何处理,眼柄切除组和触角切除组均在第1个蜕壳周期(蜕壳前期)采用镊烫法处理龙虾的眼柄和第二触角(图1(a)、(b))。在后续养殖试验阶段,若龙虾在蜕壳后重新长出触角(图1(c)),则在蜕壳后期以同样的方法处理。
试验从2019年5月10日开始,期间维持自然光周期,用黑幕布遮光,每天16:00投喂蛤肉一次,每次投喂前用电子天平(精度0.01 g)称量蛤肉湿质量,每天早上捞起残饵称重,计算当天的摄饵量。每天换水1/3,每天观察记录各试验桶龙虾的存活数和蜕壳数,将试验虾完成第1次蜕壳所经历的时间记录为第1个蜕壳周期,第2个蜕壳周期以此类推。试验期间,每隔15 d测定一次所有龙虾的体质量和体长,试验时间为120 d。
1.2.3 生长指标的测定与计算 使用电子天平(精度0.01 g)称量龙虾体质量和去掉贝壳的蛤仔肉湿质量;用游标卡尺测量龙虾的体长(眼柄基部到尾节末端)。
在此次低应变实验中,楼盘项目试验点的复合地基承载力特征值满足具体施工的设计要求,即实验所得特征值大于国家技术要求的650kPa。此外,在单桩竖向抗压实验环节,3个试验点基础桩的最大加载值都小于最大允许沉降值,且其沉降曲线基本符合标准桩的沉降趋势,即满足单桩单桩承载力950kN的使用标准。但需要注意的是,低应变实验桩检测过程中,一类桩和二类装之外的基础桩,其在质量上或多或少存在问题,需另作处理。
存活率(%)、再生率(%)、平均蜕壳周期(d)、饵料系数、摄食率(%/d)、体质量增长率(%)、体长增长率(%)和特定生长率(%/d)计算公式分别为
存活率=N2/N1×100%,
再生率=M2/M1×100%,
平均蜕壳周期=(t×N2)/N0,
饵料系数=c/(W2-W1),
摄食率=c/[t×(W1+W2) /2]×100%,
体长增长率=(L2-L1)/L1×100%,
特定生长率=(lnW2-lnW1)/t×100%。
其中:N1、N2分别为试验初始和终末龙虾存活数量(ind.);M1为切除眼柄或触角的龙虾数量(ind.),M2为眼柄或触角再生的龙虾数量(ind.);W1、W2分别为试验初始和终末龙虾的湿质量(g);L1、L2分别为试验初始和终末龙虾的体长(cm);N0为存活龙虾的蜕壳次数;c为试验期间龙虾的摄食总量(g);t为试验时间(d)。
1.3 数据处理
试验结果用平均值±标准差表示(mean±S.D.)。采用SPSS 17.0软件进行单因素方差分析,采用Duncan法进行多重比较,显著性水平设为0.05。
2 结果与分析
2.1 切除眼柄与触角后波纹龙虾的再生和存活情况
从表1可见:波纹龙虾第二触角重新长出仅出现在第2、3个蜕壳期,其他蜕壳时期未发现有再生的迹象;第1次蜕壳后触角再生率为0,第2、3次蜕壳后触角再生率达100%,表明在蜕壳前期切除触角时,波纹龙虾须经历2次蜕壳后才能完成触角再生,在进行第2次的触角切除时,龙虾还处于蜕壳后期,距离下一次蜕壳的时间间隔较长,经历1次蜕壳便获得触角再生;波纹龙虾的眼柄切除后,经历3次蜕壳还是未能获得再生;试验结束时,眼柄切除组的存活率最高(P<0.05),而对照组和触角切除组间的存活率无显著性差异(P>0.05)。

表1 切除眼柄与触角对波纹龙虾再生率和存活率的影响
2.2 切除眼柄与触角后波纹龙虾蜕壳周期的变化
从图2可见:波纹龙虾的蜕壳周期随着试验时间的增加而逐渐延长;切除触角会延长波纹龙虾第1次蜕壳的时间,但与对照组相比,触角的再生显著缩短了第2次蜕壳的时间(P<0.05),第3个蜕壳周期中各试验组无显著性差异(P>0.05)。
从表2可见:各试验组的前2个蜕壳周期总和无显著性差异(P>0.05);试验结束时,触角切除组的平均蜕壳周期最长,显著大于其他两组(P<0.05),说明频繁切除触角会延长波纹龙虾的蜕壳周期。
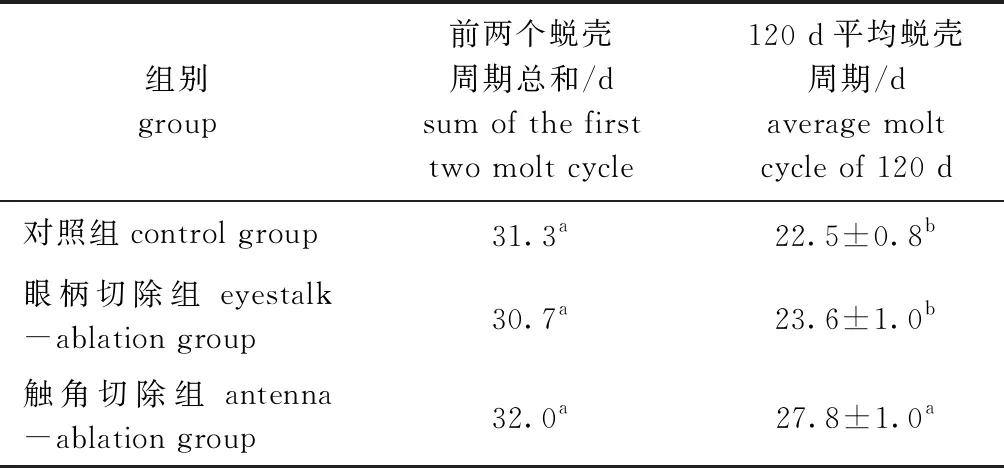
表2 切除眼柄和触角对波纹龙虾平均蜕壳周期的影响
2.3 切除眼柄与触角后波纹龙虾饵料系数和生长性能的变化
从表3可见:随着试验时间的延长,波纹龙虾的饵料系数整体呈现上升的趋势;触角切除组的饵料系数在整个养殖阶段均显著高于眼柄切除组和对照组(P<0.05);试验前60 d,对照组和眼柄切除组的饵料系数无显著性差异(P>0.05),但试验60 d后,眼柄切除组龙虾的饵料系数显著低于其他两组(P<0.05);试验结束时,各试验组的平均饵料系数存在显著性差异(P<0.05),且眼柄切除组的平均饵料系数最低(P<0.05)。

表3 切除眼柄与触角对波纹龙虾饵料系数的影响Tab.3 Effects of eyestalk and antenna ablation on food conversion ratio of scalloped spiny lobster Panulirus homarus
从表4可见:眼柄切除组龙虾的终末体质量、终末体长及体质量特定生长率均显著高于其他组(P<0.05);与对照组相比,触角的切除对波纹龙虾的各项生长指标均未产生显著性影响(P>0.05)。

表4 切除眼柄与触角对波纹龙虾生长的影响Tab.4 Effects of eyestalk and antenna ablation on growth of scalloped spiny lobster Panulirus homarus
从图3、图4可见:眼柄切除组龙虾的体质量和体长增长率在30、60、90、120 d时均显著高于其他组,但触角切除组与对照组相比无显著性差异(P>0.05)。
2.4 波纹龙虾生长、蜕壳和摄食之间的关系
从图5可见:眼柄切除组龙虾的平均蜕壳周期与对照组无显著性差异(P>0.05),但其通过提高摄食率,促进了波纹龙虾的生长;切除触角延长了波纹龙虾蜕壳周期,同时也提高了摄食率,但对龙虾的生长速度无明显影响(P>0.05)。
3 讨论
3.1 切除眼柄与触角对波纹龙虾存活的影响
在虾蟹类养殖或运输过程中,自残行为普遍存在,是影响养殖生产力和利润的重要因素之一[27-28]。自残行为受多种生物和非生物因素影响,包括食物、放养密度、蜕壳状态、光照和遮蔽物等。凡纳滨对虾Litopenaeusvannamei仔虾自残现象少,可忍受身体的相互接触,攻击频率低,是高密度工厂化养殖的首选品种[29],但在许多蟹类、龙虾类和螯虾类中,自残现象高发[30]。与其他甲壳类一样,波纹龙虾的生长过程伴随着周期性蜕壳[31],在完成蜕壳时,龙虾的新壳还未完全钙化,极易受到同类残食。据观察,本试验中对照组龙虾在试验期间的死亡均是由夜间蜕壳时的同类残食所引起的。相比之下,切除眼柄和第二触角后的龙虾感觉不灵敏、反应迟缓、攻击频率低。因此,波纹龙虾幼虾阶段眼柄和第二触角的切除有利于减少自残行为,这一结论也在相关研究中得到印证[32]。另一方面,Yang等[20]研究发现,螯足切除显著降低了中华绒螯蟹的免疫功能和抗细菌能力。本试验中也发现,触角切除组龙虾成活率低是由于频繁切除触角,伤口较大,导致病原菌入侵进而引起死亡[33]。
3.2 切除眼柄与触角对波纹龙虾蜕壳周期的影响
甲壳动物眼柄中的X-器官窦腺能够分泌性腺抑制激素(GIH)、蜕皮抑制激素(MIH)等多种神经肽类激素,蜕皮抑制激素对甲壳动物的蜕壳和生长影响较大[34-35]。眼柄的切除在多种虾蟹类的研究中被证实能够缩短蜕壳周期,包括日本沼虾Macrobrachiumnipponense[36]、短沟对虾Penaeussemisulcatus[37]、中国龙虾[17]、眼斑龙虾Panulirusargus[38]等,但在本试验中,切除单侧眼柄的波纹龙虾的蜕壳周期与对照组无显著性差异,这与对阿拉斯加帝王蟹Paralithodescamtschatica单侧眼柄切除的研究结果一致[39]。究其原因:一方面蜕壳周期与不同物种、个体大小和试验时间长短等有关;另一方面,也可能单侧眼柄的切除,不足以有效减少蜕皮抑制激素的分泌[40]。
甲壳动物在断肢后通过蜕壳可以再生,有关断肢对蜕壳周期的影响已有许多研究。岳武成等[21]研究表明,断肢延长了中华绒螯蟹的第1个蜕壳周期,而第2个蜕壳周期却相对缩短,整体蜕壳周期与对照组无显著性差异,这与本试验结果相近。波纹龙虾触角的第1次切除发生在蜕壳前期,距离下一次蜕壳的时间间隔相对较短,因此,当波纹龙虾无法通过第1次蜕壳完成触角再生时,便需要延长蜕壳周期,为下次蜕壳触角的再生做好能量储备,而第2个蜕壳周期的缩短也是生理调控的结果,以及对环境的适应[21]。第2次的触角切除发生在蜕壳后期,距离下一次蜕壳的时间间隔较长,波纹龙虾经历一次蜕壳便重新长出触角,且蜕壳周期与对照组相比无显著差异,这说明触角切除对波纹龙虾蜕壳周期的影响与其所处的蜕壳期有关。He等[41]还发现,自切附肢缩短了幼蟹的蜕壳周期,并且蜕壳周期与自切附肢的数量呈负相关。其他相关研究表明,失去附肢对蜕壳周期的影响不仅取决于蜕壳期和失去附肢数量,还与附肢类型及受损程度有关[32, 42-43]。
3.3 切除眼柄和触角对波纹龙虾生长性能的影响
与其他甲壳动物一样,波纹龙虾通过蜕壳实现快速生长,蜕壳间期的营养物质积累是影响蜕壳后生长的重要因素[44]。切除眼柄的龙虾行动和反应迟缓,代谢水平降低,减少了生长过程的能量损耗,而且其摄食率高,饵料系数低,有利于在下次蜕壳前储备更多的营养物质,使其生长性能要优于对照组。王芳等[45]对凡纳滨对虾稚虾的研究表明,去除眼柄对凡纳滨对虾稚虾摄食率的影响无统计学意义。除了由于波纹龙虾和凡纳滨对虾分属十足目的不同下目外,也可能与试验时间的长短有关。
附肢损伤会降低甲壳类的觅食效率和交配成功率,并降低种内和种间的资源竞争力,影响程度取决于失去附肢的类型和数量[46]。甲壳动物的摄食主要依靠化学感受器[47],龙虾嗅觉感受器存在于第一对触角上,而味觉感受器存在于口器和颚足上,据此可推测,切除第二触角并不会影响龙虾的摄食活动。较高的饵料系数主要是因为第二触角频繁切除与再生所造成的摄食成本的增加[48]。附肢再生需要消耗能量,这部分能量来源于储存的营养[49],失去触角的龙虾通过增加进食量以满足能量需求,具有较高的摄食率,但频繁切除触角导致动物机体用于生长的营养储备减少,最终未能提高其生长速度。
4 结论
1)波纹龙虾眼柄的切除有助于提高养殖存活率,促进其摄食和生长,在养殖生产上值得借鉴和推广。
2)触角的切除对其生长没有益处,不但提高了饵料系数,增加了养殖成本,还可能因病原入侵而导致死亡。因此,在养殖生产中应谨慎捕捉波纹龙虾,以保持其触角完整。
