向日葵油酸合成上游基因HaFAB2克隆与表达分析
2020-12-17周菲
周 菲
(1.黑龙江省农业科学院博士后科研工作站,哈尔滨 150086;2.黑龙江省农业科学院经济作物研究所,哈尔滨 150086)
向日葵(Helianthus annuusL.)是重要油料和经济作物[1]。我国向日葵产区主要分布在东北、西北和华北,冷凉气候条件下生产的葵花籽一般油酸含量相对较低,因此,提高向日葵种子油酸含量是我国向日葵遗传改良重要目标。
植物脂肪酸合成代谢通路主要包含脂肪酸脱饱和酶(FAD)、乙酰CoA羧化酶(ACCase)及脂肪酸合酶复合体(FAS)等。近年来,关于脂肪酸含量相关酶基因研究越来越多。至今在大豆中已分离得到脂肪酸合成酶相关基因包括ACC、FatB、KASⅠ、KASⅡ、KASⅢ、SACPD等[2]。将大豆中GmACP基因沉默后,大豆根中总脂肪酸含量降低22%[3]。将高山被孢霉(Mortierella alpina)中Δ6脱饱和酶的同工酶基因Δ6II转化到米曲菌(Aspergillus oryzae)中表达,导致γ-亚麻酸(GLA)占总脂肪酸37%,而在对照米曲菌(Aspergillus oryzae)中未检测到γ-亚麻酸[4]。将玻璃苣种子中分离的Δ6-脂肪酸去饱和酶基因转化到烟草中,对转基因烟草叶片脂质分析表明,γ-亚麻酸(GLA)和十八碳四烯酸(C18∶4 Δ6,9,12,15)分别积累到总脂肪酸含量13.2%和9.6%[5]。将拟南芥中Δ15脂肪酸去饱和酶转化到大豆,其α-亚麻酸(ALA)含量显著升高,而同时具有玻璃苣中Δ6-脂肪酸去饱和酶和拟南芥中Δ15脂肪酸去饱和酶两种去饱和酶的转基因大豆,其十八碳四烯酸(STA)相对含量明显提高,ω-3类脂肪酸相对含量达到60%以上[6]。在亚麻中通过体内体外试验发现,LuLACS8A和LuDGAT2-3共表达催化亚麻酸在三酰甘油(TAG)中富集[7]。研究发现植物中转录因子对油脂和脂肪酸合成也具有重要调控作用,这些转录因子主要包括AP2家族WRI1、B3家族FUS3、DOF家族DOF、HAP3/CBP家族LEC1和CHD3家族PKL等,利用代谢调控网络发挥调控脂肪酸功能[8]。
油酸(18∶1)第9位与第10位碳原子间含一个双键因此成为单不饱和脂肪酸,第12位碳原子脱饱和后形成亚油酸(18∶2)[9]。目前向日葵与油酸含量相关基因研究较少,Δ12脂肪酸脱氢酶基因位于油酸合成通路下游,控制油酸转化成亚油酸,是控制种子油酸含量关键基因[10]。油料作物如大豆、玉米、油菜、花生种子中,通过RNA干扰等技术抑制FAD2基因表达抑制油酸合成亚油酸,显著提高种子油酸含量[11]。FAB2(Stearoyl-ACP Desaturase 2)编码Δ9硬脂酰-ACP脱饱和酶,是植物体内调控油酸合成上游基因,在18碳脂肪酸长链第9和第10个碳原子之间插入一个双键,使硬脂酰-ACP脱氢生成油酰ACP,油酰-ACP转运至内质网后形成油酸,该过程直接影响油酸含量。FAB2在大部分高等植物不同组织部位中具有表达特异性,在发育种子中表达量最高[12]。FAB2是脂肪酸合成过程中饱和脂肪酸转化成不饱和脂肪酸关键分支点,是不饱和脂肪酸合成中必不可少的重要酶,FAB2可能是除FAD2外控制油酸含量另一个关键基因[9],此外,FAB2还参与植物膜脂质中不饱和脂肪酸生物合成,关系到细胞膜流动性与耐受性,具有抗非生物胁迫等重要功能[13]。但目前关于向日葵FAB2基因研究未见报道,其编码蛋白结构、表达模式及功能尚不明确。
向日葵脂肪酸代谢关键酶基因克隆和表达模式分析是基因功能研究重要前提,可为向日葵脂肪酸合成分子机理解析提供科学依据。本课题组前期利用高低油酸向日葵种子转录组测序结果,在脂肪酸代谢通路差异表达基因中挖掘出可能影响油酸含量的候选基因硬脂酰-ACP脱氢酶基因HaFAB2,以此为基础,本研究克隆向日葵Ha-FAB2基因,并作生物信息学分析,结合高低油酸向日葵种子中油酸积累变化趋势,分析HaFAB2基因在不同发育时期种子及其他组织部位表达模式,为该基因功能研究提供参考。
1 材料与方法
1.1 材料
1.1.1 植物材料
向日葵高油酸保持系“L-1-OL-1”和低油酸保持系“86-1”于2019年种植于黑龙江省农业科学院经济作物研究所试验基地。于同一时段对当日开花植株挂牌,标明开花日期,从开花7 d起,每5 d取5个花盘,取至开花后37 d,取花盘最外3圈种子,一部分样品风干后测定油酸含量;另一部分液氮速冻后冻存于-80℃冰箱,用于基因克隆和荧光定量PCR分析,基因克隆利用“86-1”开花7 d种子为试验材料。
1.1.2 菌株与载体
克隆载体pMD18-T(TaKaRa),大肠杆菌DH5α感受态细胞(博凌科为)。
1.1.3 主要试剂
所用试剂包括DNA marker DL2000(东盛)、2×KOD PCR MasterMix(艾德莱)、2×Taq PCR Master-Mix(艾德莱)、gel-red染料(莫纳)、氨苄青霉素(Amp)、琼脂糖DNA凝胶回收试剂盒(天根)、无水乙醇、β巯基乙醇、50×TAE。
1.1.4 培养基配制
液体LB培养基:称取10 g蛋白胨、5 g酵母提取物和10 g NaCl,溶解于1 L水中。若配制固体LB培养基每升再加12 g琼脂粉;IPTG溶液(24 mg·mL-1):称取0.24 g IPTG,溶解于10 mL去离子水,完全溶解后过滤并灭菌;X-Gal溶液(20 mg·mL-1):称取0.1 g X-Gal,溶解于1 mL二甲基甲酰胺(DMF)中,最后加DMF定容至5 mL。
1.2 方法
1.2.1 种子RNA提取及cDNA第一条链合成
利用植物总RNA提取试剂盒(天根)提取向日葵种子总RNA,取0.5 μg RNA利用ReverTra Ace qPCR RT Master Mix with gDNA Remover(TOYOBO)合成cDNA第一条链。
1.2.2 HaFAB2基因克隆
根据已公布向日葵基因组数据库信息,设计向日葵HaFAB2基因克隆引物,引物序列为F:AGAACAACATCAACAATGGCGATTC,R:AGCAC TACACGAAAGAAACGACAAT,使用高保真酶(艾德莱)扩增HaFAB2基因开放阅读框(ORF)序列,PCR总反应体系20 μL:cDNA 1 μL,Primer各1 μL,2×KOD PCR MasterMix 10 μL,ddH2O 7 μL。PCR扩增程序为:94℃3 min,94℃30s,58℃30 s,72℃90 s,共30个循环,最后72℃延伸7 min。
扩增产物经1%琼脂糖凝胶电泳分离,利用琼脂糖凝胶DNA回收试剂盒(天根)回收目的条带,并克隆至pMD18-T载体(TaKaRa),挑取阳性克隆,经菌液PCR验证送吉林省库美生物有限公司测序。
1.2.3 基因生物信息学分析
利用DNAMAN比对克隆基因序列和NCBI公布向日葵基因序列。在NCBI BLAST数据库中作序列同源性比对和相似性搜索。MEGA7软件构建系统进化树。利用ExPASy系统下蛋白质基本理化性质预测工具Protparam(https://web.expasy.org/protparam/)预测氨基酸组成、相对分子质量、等电点和原子组成等参数。利用SOPMA在线程序(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)分析蛋白二级结构。利用Protscale在线软件(http://web.expasy.org/protscale/)分析基因氨基酸序列亲/疏水性。
1.2.4 种子油酸含量检测
向日葵种子油酸含量委托中国农业科学院油料作物研究所测定,采用气相色谱法,仪器为Agilent7890A气相色谱仪。利用SPSS 13.0软件分析种子发育不同时期油酸含量变化差异显著性。
1.2.5 荧光定量PCR分析
采用 THUNDERBIRD SYBR qPCR Mix(TOYOBO)试剂,Roche LightCycler 480II荧光定量PCR仪,以向日葵β-actin基因(AF282624)为内参基因,Real-time PCR总反应体系为20 μL:cDNA 1.2 μL, SYBR Green 10 μL, Primer 各 0.6 μL,ddH2O 7.6 μL。Real-time PCR 反应程序如下:98 ℃ 2 min,98 ℃ 10 s,58 ℃ 10 s,68℃ 30 s,共45个循环,待PCR反应结束后分析溶解曲线,采用2-ΔΔCt法分析目标基因表达量。内参基因β-actin引物为F:GCAAAAAGCAGCTCGTCTGT,R:AGCAGCTTCCATTCCAATCA,HaFAB2荧光定量引物为 F:CTGGTCAAACAGTCAACATATGGGT,R:CA GCGGTTATGGTGAGGT。利用SPSS 13.0软件分析基因在“L-1-OL-1”和“86-1”种子中表达量差异显著性。
2 结果与分析
2.1 向日葵HaFAB2基因克隆
将提取的“86-1”开花7 d种子总RNA反转录合成cDNA,使用设计好的引物扩增HaFAB2开放阅读框(ORF)序列,PCR结果显示有符合目的片段长度的单一条带(见图1)。将胶回收后目的片段与T载体连接并转化大肠杆菌,菌液PCR检测结果表明扩增出阳性条带符合目的片段长度,将菌液送至公司测序,比对结果表明序列正确。向日葵Ha-FAB2基因ORF核苷酸序列长度为1 191 bp,编码396个氨基酸,通过比较克隆的HaFAB2和NCBI公布向日葵FAB2(LOC110878160)序列发现,核苷酸序列相似度为99.16%(见图2)。

图1 向日葵HaFAB2基因PCR电泳结果Fig.1 PCR electrophoresis result of sunflower HaFAB2 genes

图2 克隆HaFAB2与公布向日葵FAB2核苷酸序列比对结果Fig.2 Comparison of nucleotide sequences of HaFAB2 cloned with FAB2 published in sunflower
2.2 HaFAB2基因生物信息学分析
2.2.1 向日葵HaFAB2编码蛋白质基本理化性质预测
ExPASy软件预测结果表明,HaFAB2基因编码蛋白原子总数6 307个,分子式C2014H3131N551O594S17,蛋白分子质量45.11 ku,等电点理论值5.99,预测该蛋白为酸性蛋白,HaFAB2编码蛋白质氨基酸组成见表1。
2.2.2 向日葵HaFAB2基因系统进化分析
为分析向日葵HaFAB2基因进化关系,将其与拟南芥(Arabidopsis thalianaL.)、大豆(Glycine maxL.)、异果菊(Dimorphotheca sinuataDC.)、烟草(Nicotiana tabacumL.)和薇甘菊(Mikania micranthaKunth)等物种FAB2氨基酸序列作系统进化树构建与分析(见图3A)。
上述结果可知,向日葵HaFAB2与薇甘菊(Mikania micranthaKunth)FAB2聚在同一分支上,进化关系最近,序列相似性高达94%,可能是由于向日葵和薇甘菊同属菊科,具有物种同源性。将NCBI上检索到向日葵FAB家族3个成员HaFAB1(CAC80359.1)、HaFAB2(XP_021982111.1)、HaFAB3(KAF5783066.1)与拟南芥硬脂酰-ACP脱饱和酶(SAD)家族7个成员AtSSI2(At2g43710)、AtSAD1(At5g16240)、AtSAD2(At3g02610)、AtSAD3(At5g 16230)、AtSAD4(At3g02620)、AtSAD5(At3g02630)、AtSAD6(At1g43800)以及蓖麻RcSAD1(NP_00131 0659.1)作进化关系分析(见图3B)。其中,AtSSI2和RcSAD1对18∶0-ACP具有底物选择性,催化18∶0-ACP生成18∶1-ACP,结果发现HaFAB1、HaFAB2、HaFAB3序列相似性较高,并与拟南芥AtSSI2和蓖麻RcSAD1聚在同一个分支上,其中HaFAB2与AtSSI2、RcSAD1序列相似性分别高达80.55%和84.60%,表明亲缘关系较近。

表1 向日葵HaFAB2编码蛋白质氨基酸组成Table 1 Amino acid composition of protein encoded by HaFAB2 in sunflower
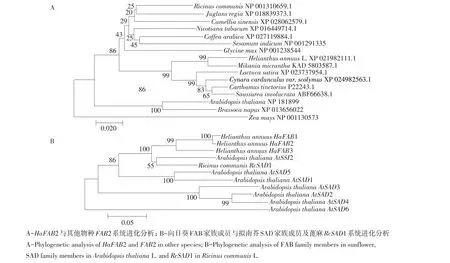
图3 向日葵HaFAB2基因系统进化树分析Fig.3 Phylogenetic tree analysis of HaFAB2 gene in sunflower
2.2.3 蛋白质二级结构预测
通过使用SOPMA在线软件预测分析HaFAB2编码蛋白质二级结构,结果表明,该蛋白质具有53.54%α螺旋,33.84%无规则卷曲,7.58%延伸链,5.05%β转角。预测结果表明,HaFAB2编码蛋白二级结构为α螺旋(见图4)。
图4 向日葵HaFAB2编码蛋白质二级结构预测Fig.4 Secondary structure prediction of protein encoded by HaFAB2 in sunflower
2.2.4 蛋白质亲疏水性分析
蛋白质亲疏水性一定程度上决定蛋白折叠方式,是蛋白质空间结构因素之一,在蛋白特定功能上发挥重要作用。使用ProtScale分析HaFAB2编码蛋白氨基酸序列亲疏水性。>0部分高于50%为疏水性蛋白,而<0部分高于50%为亲水性蛋白,可知HaFAB2编码蛋白为亲水性蛋白(见图5)。
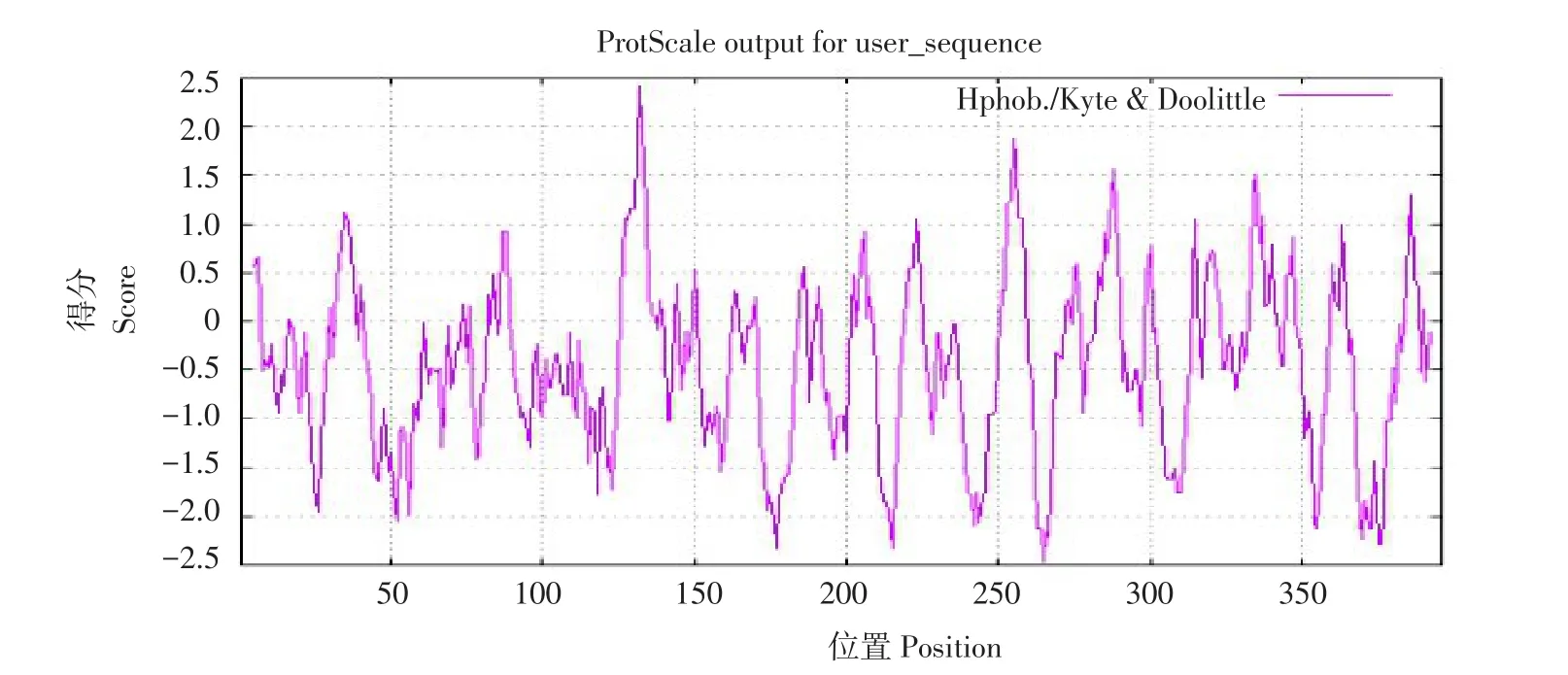
图5 向日葵HaFAB2编码蛋白质亲/疏水性分析Fig.5 Hydrophilic/hydrophobic analysis of protein encoded by HaFAB2 in sunflower
2.3 HaFAB2基因表达分析
测定向日葵种子不同发育时期油酸含量,高、低油酸含量的两份向日葵材料在种子发育过程中油酸积累变化趋势一致,但稍有不同,在向日葵种子发育初期至中期(开花后7 d到开花后22 d),随发育时间增加,油酸含量均迅速增长,“L-1-OL-1”在开花后22 d油酸含量有小幅增长,开花后37 d油酸含量达最高值。而“86-1”在22DAF油酸含量达最高,之后油酸含量下降,开花后27 d趋于稳定(见图6A)。
通过qRT-PCR检测向日葵HaFAB2基因在高低油酸向日葵种子不同发育时期以及“L-1-OL-1”在开花后7 d根、茎、叶、管状花、舌状花表达量。结果表明,HaFAB2在各部位均有不同程度表达,发育早期种子和成熟叶中表达较高。7DAF-17DAF两份材料种子中表达量均较高,且均在17DAF种子中表达量最高,此时“L-1-OL-1”明显高于“86-1”,油酸积累变化趋势(见图6A)结果表明,7DAF-17DAF种子油酸处于逐渐积累时期,可看出,HaFAB2基因表达量与油酸积累变化趋势一致。17DAF之后,HaFAB2基因在大部分发育时期种子中较低表达,但在27DAF“86-1”种子中较高表达量,此时明显高于“L-1-OL-1”(见图6B)。

图6 向日葵HaFAB2基因表达分析Fig.6 Expression analysis of HaFAB2 in sunflower
3 讨论与结论
Δ9硬脂酰-ACP脱氢酶基因FAB2使硬脂酸脱饱和后形成油酸。由于FAB2催化去饱和作用是植物不饱和脂肪酸生物合成第一步,在很大程度上决定植物中饱和脂肪酸与不饱和脂肪酸比例。在玉米种子中过表达FAB2同源基因ZmSAD1,不饱和脂肪酸与饱和脂肪酸比例提高20.40%~20.61%[14]。在棉花中利用RNA介导基因沉默技术干扰FAB2同源基因SAD表达,硬脂酸含量从20%提高到40%[15]。在大豆中,通过抑制FAB2基因,可增加硬脂酸含量[16]。异位表达花生FAB2提高酵母体内油酸含量[17]。Kachroo等研究拟南芥FAB2突变体,发现突变体油酸含量偏低[18]。本研究克隆向日葵HaFAB2基因,并分析基因生物信息学和研究表达模式,为进一步研究HaFAB2基因功能提供参考,为通过分子育种手段改良向日葵油品质建立基础。
在大部分高等植物中,FAB2在各器官中均表达,但表达模式不同。FAB2同源基因SAD在油樟种子、叶和花中表达量较高,但在根和茎中表达量低[19]。在花生克隆的3个FAB2同源基因中,FAB2-1与FAB2-2在种子中表达量较高,随种子发育两者表达水平持续升高;FAB2-3在花中表达量较高,仅在种子发育早期呈上调表达趋势[17]。而海滨锦葵SAD在叶中表达量最高,其次是种子、茎和花中表达量最低,且随种子发育表达量逐渐降低[20]。GhSAD2基因在棉花叶片中表达量高于茎和根;GhSAD2基因在棉花种子发育过程中表达量呈先升后降趋势,在花后25 d种子中表达量最高,在成熟种子中表达相对较弱[21],与本研究结果相近,本研究HaFAB2基因在各部位均表达,尤其是叶、种子中表达量较高,在7DAF-17DAF两份材料种子中表达量均较高,种子中油酸快速积累,且Ha-FAB2均在17DAF种子中表达量达到最高,此时高油酸材料中表达量明显高于低油酸材料,HaFAB2催化硬脂酸生成油酸,因此其表达越高油酸积累较多,可看出HaFAB2基因表达量变化与油酸积累变化趋势一致,推测HaFAB2基因是影响油酸含量的重要基因。目前已利用人工诱导的FAD2突变体选育出高油酸向日葵品种Pervenets,将葵花籽亚油酸含量从66%降低到30%(单基因突变体)和12%(双基因突变体),油酸含量从20%分别上升至60%和80%[22]。目前,各国高油酸向日葵品种多是以Pervenets为基础材料育成。FAD2突变显著提高种子油酸含量,若在向日葵fad2突变体中过表达Ha-FAB2,理论上可提高种子中总不饱和脂肪酸和油酸含量,基因编辑技术发展将带动培育出更多高油酸向日葵新品种。
FAB2还与多种生理生化反应以及抗胁迫相关。拟南芥FAB2突变体内硬脂酸(C18∶0)含量增加,油酸(C18∶1)含量减少,提高水杨酸与茉莉酸介导的抗病反应[23]。过表达小麦FAB2同源基因TaSSI2,转基因拟南芥植株对白粉病抗性降低[24]。低温下培养甘蓝型油菜FAB2的一个同功基因上调表达,导致硬脂酰ACP去饱和酶含量增加[25]。RNAi的GhSAD2转基因棉花植株相比对照,其抗寒能力减弱,耐热能力增强[21]。FAB2基因还与植物抗炭疽病菌和植物激素信号转导等密切相关[26-27]。本研究中在17DAF之后,HaFAB2基因在大部分发育时期种子中表达较低,但在27DAF“86-1”种子中表达量较高,可能与某种生理生化反应及胁迫相关,值得未来深入研究。本试验初步研究HaFAB2基因在高低油酸种子表达模式,推测其在调控向日葵脂肪酸合成尤其是油酸合成可能发挥功能,但需通过转基因等技术验证。
