翘嘴鳜·斑鳜·杂交鳜不同组织3种同工酶特征研究
2020-12-11梁家僖薛源史杨白
梁家僖 薛源 史杨白
摘要 揭示翘嘴鳜、斑鳜和杂交鳜F1 [翘嘴鳜(♀)×斑鳜(♂)]的同工酶生化遗传特性。采用非变性聚丙烯酰胺凝胶电泳(Native-PAGE)检测技术,分别进行了斑鳜、翘嘴鳜3个组织(肌肉、肝脏、肾脏)以及杂交鳜F1代5个组织(肌肉、肝脏、心脏、肾脏、脑)的3种同工酶(LDH 1.1.1.27、MDH1.1.1.37、EST3.1.1.1)酶谱特征的研究。3种同工酶均存在较稳定的物种和组织特异性表达的特征;杂交鳜F1代同工酶酶谱的组织表达与双亲(斑鳜、翘嘴鳜)之间存在明显的亲缘关系和特异性,除了斑鳜和翘嘴鳜肾脏中的MDH同工酶谱以外,3个不同鳜种间的酶谱特征均存在较明显的种属特异性,肝脏、肾脏中的LDH和EST同工酶谱以及肌肉中的EST同工酶谱均存在显著且稳定种属特异性差异表达,可作为鉴别3种鳜鱼种质特性的重要遗传生化参考指标。该研究结果为鳜鱼的种质鉴定和杂交育种提供了一定的理论依据,并具有较好的应用价值。
关键词 翘嘴鳜;斑鳜;杂交鳜;同工酶
中图分类号 S917.4 文献标识码 A 文章编号 0517-6611(2020)21-0109-06
Abstract In order to reveal the isozyme genetic characteristics of Siniperca chuatsi, Siniperca scherzeri and hybrid mandarin fish [Siniperca chuatsi (♀) × Siniperca scherzeri (♂)], using native polyacrylamide gel vertical electrophoresis technology, three isozymes (LDH 1.1.1.27, MDH1.1.1.37, EST3.1.1.1) of five tissues (muscle, liver, heart, kidney, brain) of hybridization F1 and three tissues (muscle, liver, kidney) of Siniperca chuatsi, Siniperca scherzeri were analyzed. The results showed that there were stable species and tissue specific expression characteristics in three isozymes. The tissue expression of isozyme of hybridization F1 had obvious relationship and specificity with its parents [Siniperca chuatsi (♀) × Siniperca scherzeri (♂)]. Except for the MDH isozyme zymogram of kidney of Siniperca scherzeri and Siniperca chuatsi, the isozyme characteristics of three species of mandarin fish showed significant species specificity. The LDH and EST isozyme zymogram of kidney and liver and EST isozyme zymogram of muscle had remarkable and stable species differential expression,which could be used as a key biochemical indicator to distinguish the germplasm characteristics of three species of mandarin fish. Therefore, the results of this study provided a theoretical basis for germplasm identification and hybrid breeding of these three species of mandarin fish, and had a good application value.
Key words Siniperca chuatsi;Siniperca scherzeri;Hybrid mandarin fish;Isozyme
基金項目 苏州大学校级大学生创新创业训练计划(2019xj069);江苏省重点研发计划(现代农业)(BE2017311);江苏省渔业科技类项目(Y2017-38);江苏省重点研发计划(现代农业)(BE2018398)。
作者简介 梁家僖(1996—),女,广西梧州人,从事鱼类生物学研究。*通信作者,副教授,博士,从事水生生物学研究。
收稿日期 2020-02-29
斑鳜(Siniperca scherzeri)和翘嘴鳜(Siniperca chuatsi)同属鲈形目、鲈亚目、鮨科、鳜亚科、鳜属,均为肉食性底栖凶猛鱼类,营掠性摄食方式,肉质细嫩,味道鲜美,是名贵的经济鱼类。斑鳜的运动和捕食能力较翘嘴鳜强[1],肌肉蛋白质含量显著高于翘嘴鳜(P<0.05)[2],但翘嘴鳜的生长速度快且体型较大,具有更高的商业养殖价值。利用父本斑鳜体型、体色、优良品质和母本翘嘴鳜生长速度快的双亲遗传特性开展杂交育种获得的杂交鳜可显现出明显的优良性状,具有较好的商业开发前景[3]。目前,对翘嘴鳜、斑鳜及其杂交种的研究主要集中于亲本与杂交鳜的生物学特征分析[4]、杂交鳜的胚胎发育研究[5-9]、养殖技术研究[10-14]、营养学特征分析[2,15]、遗传特征的微卫星分析等方面[16-17],但对其同工酶的研究鲜见报道。
2.1.3 酯酶(EST 3.1.1.1)。
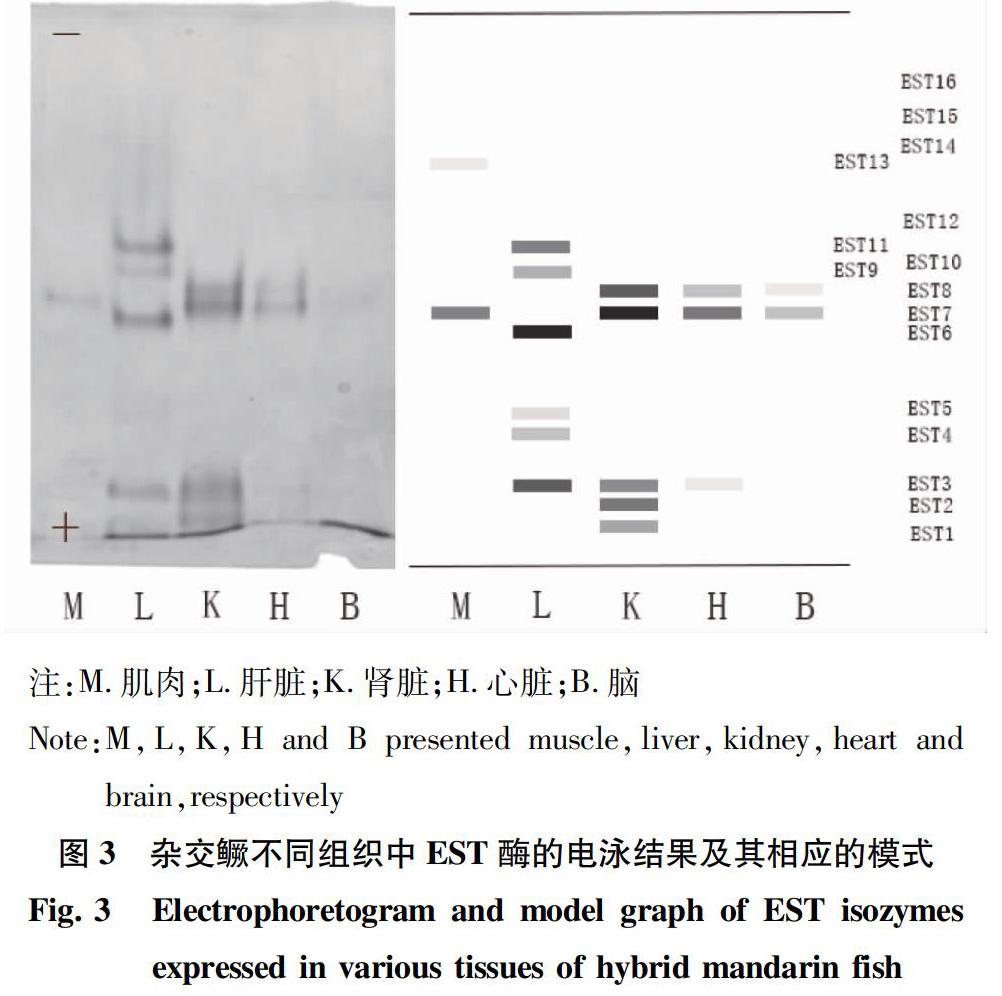
酯酶可在水分子的参与下,经由水解作用,将酯类切割成酸类与醇类。鱼类的酯酶一般是单体酶或二聚体酶,由多个基因座位控制。试验结果显示:在杂交鳜F1代的5种组织中共检测到15条酶带(图3),不同组织的 EST同工酶存在着基因座位的多态性,各组织的酶谱带型分别为M7(13)、L63(11)945、K78231、H783、B78,带型码完全不同,呈现出较强的组织特异性。EST同工酶在肝脏、肾脏组织中的活性很高,且酶带数很多;但在肌肉、脑组织中活性很低,且酶带数最少。具体而言,EST4、EST5、EST6、EST9、EST11仅在肝脏中有特异表达,其中,EST6、EST11的活性较强;EST1、EST2仅在肾脏中表达,且EST2、EST3、EST7、EST8在肾脏中优势表达;EST13仅在肌肉中表达。
2.2 杂交鳜F1[翘嘴鳜(♀)×斑鳜(♂)]与亲本翘嘴鳜、斑鳜的3种组织同工酶谱比较
2.2.1 3种鳜鱼的LDH酶谱比较。
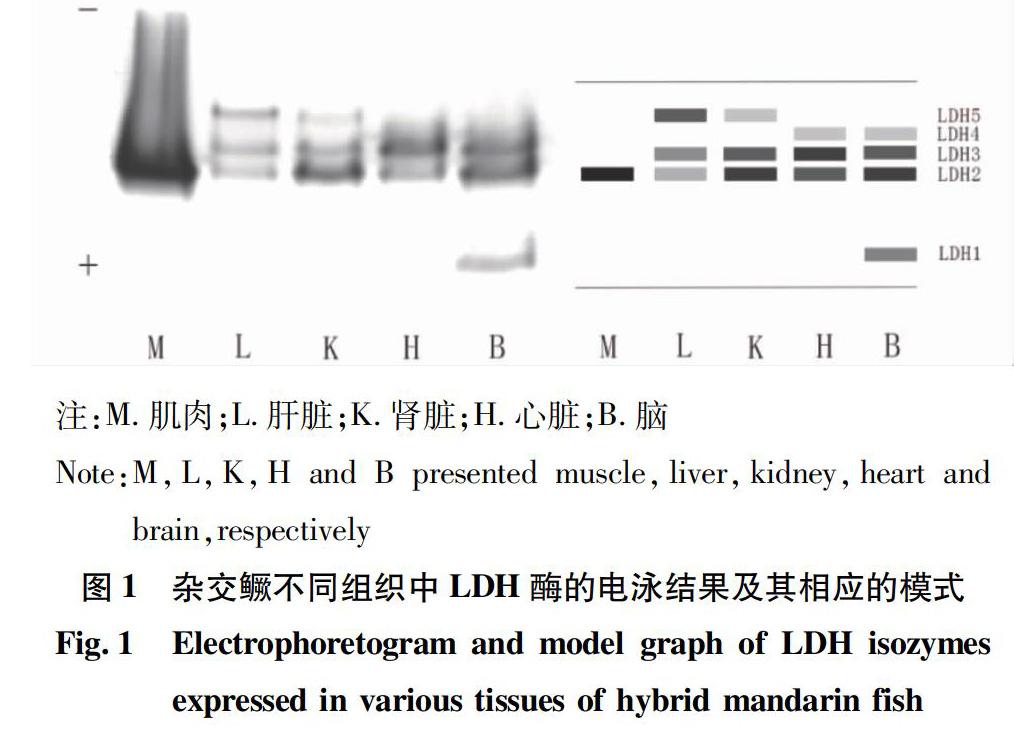
对杂交鳜F1代和亲本(斑鳜、翘嘴鳜)各自的3种组织(肌肉、肝脏、肾脏)同时进行了同工酶表达谱的比较试验。结果表明:杂交鳜F1代与亲本斑鳜、翘嘴鳜之间LDH同工酶谱存在明显差异(图4)。肌肉组织中均有LDH2条带表达,活性较强,但翘嘴鳜还显示出LDH4条带;肝脏组织中LDH酶谱带型差异显著:杂交鳜LZ532,斑鳜LB35,翘嘴鳜LQ342,杂交鳜F1代相比斑鳜多了1条LDH2条带,而相比翘嘴鳜少了1条LDH4酶带、多了1条LDH5酶带,表明杂交子代形成了杂合子,可作为鉴别3种鳜属鱼的生化标志;在肾脏中,3种鳜鱼的LDH酶谱带型分别为杂交鳜KZ235,斑鳜KB2345,翘嘴鳜KQ234,杂交鳜F1代缺少双亲的LDH4条带,显示出表达差异,而在3种鳜鱼的肾脏中均呈现出LDH23酶谱带型。
2.2.2 3种鳜鱼的MDH酶谱比较。
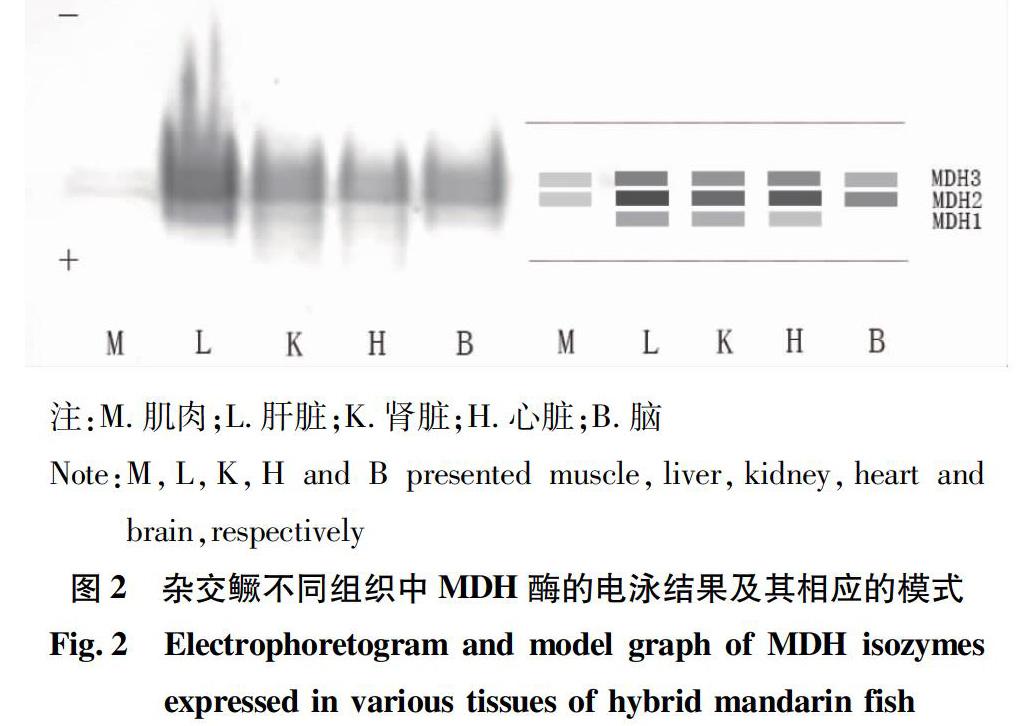
从图5可以看出,杂交鳜F1代的MDH表达在肌肉、肝脏中没有表现出明显差异,三者的肌肉MDH带型码均为M23,且表达水平普遍很低;肝脏MDH酶谱带型均为L231,且条带的相对活性一致;在肾脏中MDH酶谱带型分别为杂交鳜KZ231,斑鳜KB23,翘嘴鳜KQ23,其中,杂交鳜的表达活性较高,而且相比双亲多了1个MDH1条带,这些酶谱带型可作为鉴定杂交鳜的依据之一。
2.2.3 3种鳜鱼的EST酶谱比较。
从图6可以看出,EST酶在3种鳜鱼肌肉组织中的表达水平偏低,在肝脏、肾脏组织中表达水平较高。在肌肉组织中,杂交鳜F1代表达出双亲没有的EST7条带,具有与斑鳜相同的EST13酶带,而翘嘴鳜没有此酶带;在肝脏组织中EST酶的酶谱带型分别为杂交鳜LZ6(11)3945,斑鳜LB62(11)(14)(15)94(16)5,翘嘴鳜LQ27(12)(14)4(10)5,显示出不同的谱型特征;杂交鳜F1代EST酶谱条带少于双亲,但酶谱活性较高;在肾脏组织中EST酶的酶谱带型也有差异,分别为杂交鳜KZ78231、斑鳜KB72、翘嘴鳜KQ732,杂交鳜F1代增加了双亲不具有的EST8和EST1酶带,表达的相对活性与翘嘴鳜相近。
3 讨论
3.1 影响同工酶试验结果的几个因素
聚丙烯酰胺凝胶电泳法包括制胶、制样、加样、电泳、染色等主要步骤,每一个步骤对测定结果都至关重要,若操作不当,则会出现酶带分离效果不佳、有拖尾现象、酶失活导致无法染色等问题。笔者认为影响试验结果的因素主要包括以下方面:①样品中的脂肪影响。葛彦龙等[42]指出,酶带出现拖尾现象极有可能是由样品中的脂类造成的,可用氯仿脱脂以防止谱带出现拖尾现象。②上样缓冲液中蔗糖或甘油的影响。该部分试验中,上样缓冲液中添加了蔗糖,作用是保护同工酶。③上样时样品浓度的影响。点样液中同工酶的浓度过低,则酶带染色很浅,肉眼无法观察到以致于被误认为无条带,故应根据具体的鱼种类或组织调整点样液中酶的浓度。④电压大小的影响。应提高电泳电压,防止扩散,同时缩短时间,但电压过高、电流过大会产生热量过大,造成酶的失活。⑤胶的浓度。应根据检测的酶分子量大小选择合适的胶浓度,浓度过高,最终酶的条带过短,不利于观察;浓度过低,则分离效果不佳。葛彦龙等[42]在采用更“剧烈的”试验条件时,发现酶并没有失活,这么操作实际上是使酶的活性降到最低,避免样品中的酶会因为细胞破裂而相遇、互相降解。这与普遍认为的在样品研磨处理时应该保持低温、在电泳过程中也尽量保持温和的条件不同。综合以上几个方面因素以及试验操作,笔者认为在酶液提取时应动作迅速,脱脂后再进行电泳,且要避免将制好的样品反复冷冻解冻;电泳时应选择220 V电压。分离胶浓度为8%时对EST同工酶分离效果较好,LDH、MDH同工酶的最适分离胶浓度则有待探索。至于在上样缓冲液中添加蔗糖或甘油是否对酶有保护作用或是否会造成拖尾现象也应做进一步的对比分析研究。
3.2 斑鳜、翘嘴鳜同工酶图谱
该研究中得出的斑鳜、翘嘴鳜同工酶图谱与已有的研究报道存在某些差异。例如吴兴兵等[49]报道太湖水域野生翘嘴鳜亲鱼繁殖后代肌肉中的LDH酶带数为2,与该试验结果一致。但杨军峰等[50]报道武汉水域的翘嘴鳜肌肉中的EST酶带数比斑鳜多1条,与该研究结果(斑鳜肌肉中的EST酶带数比翘嘴鳜多1条)相反;另外,翘嘴鳜的肾脏EST酶谱结果显示1条酶带,而该研究结果显示有3条酶带;斑鳜的肝脏组织中LDH酶带仅检测到1条,此次试验检测出2条。究其原因,可能是种群之间有一定程度的遗传分化。例如长江、珠江和黑龙江三水系的草鱼(Ctenopharynodon idellus)种群之间存在明显的生化遗传差异[51]。表明同一种鱼类的不同种群间既具有共同的酶谱特征,又具有一定的种群特异性,而且同一类群内不同个体间亲缘关系的远近也会影响基因座位的多态性,进而影响同工酶的表达。另一方面,即使同一种鱼也会随着生态环境的改变发生生理生化方面的改变。例如,香鱼(Plecoglossus altivelis)因生活环境发生变化而形成了洄游型和陆封型2类,使两者的LDH同工酶存在差异[52]。蓝绿鳞鳃太阳鱼(Lepomis cyanellus)体内乳酸脱氢酶和苹果酸脱氢酶的活性水平会受到环境温度变化响应而发生显著变化[53]。可吸气式呼吸的翠鳢(Channa punctata)LDH同工酶的表达与是否接触空气有关[54]。通过该试验结果与已有的研究报道[49-50,55-56]對比分析发现,酶谱特征比较稳定是翘嘴鳜肌肉LDH同工酶酶带数为2,斑鳜肾脏LDH同工酶酶带数为4,这一酶谱特征能可靠地用于种质鉴定。
3.3 杂交鳜F1[翘嘴鳜(♀)×斑鳜(♂)]同工酶图谱
该研究首次报道了杂交鳜F1代5种组织的3种同工酶电泳图谱,结果显示,同工酶LDH和EST在各组织中的表达存在显著差异,而同工酶MDH的表达差异不大。LDH5酶带在杂交鳜F1代的肝脏中具有相对高的活性,而在其他组织中的相对活性低或不表达。LDH1酶带仅在杂交鳜F1代的脑组织有特异性的表达,而在其他组织中未有表达。EST在杂交鳜F1代5种组织中的酶谱特征都不相同,存在明显的组织表达特异性,表明该酶会根据不同组织的不同生理生化需要进行选择性表达。而MDH在除了杂交鳜F1代肌肉组织以外的4种组织中的表达水平都较高,酶谱的相似度也较高,且带型都比较简单。
3.4 杂交鳜F1[翘嘴鳜(♀)×斑鱖(♂)]与亲本翘嘴鳜、斑鳜的同工酶比较
同工酶表达的种质和组织特异性,与不同物种和器官组织的生理功能以及基因的调控有关。不同物种或种群之间存在着基因座位多态性,是划分天然种群和亚种群的重要手段。通过同工酶分析可以了解不同物种基因表达与调控情况。对试验结果分析后发现,杂交鳜F1代与亲本斑鳜、翘嘴鳜的同工酶酶谱间既有联系又存在差异,表明杂交鳜获得了双亲的遗传物质和生理生化性状。例如,编码乳酸脱氢酶亚基的3个基因通过分子杂交、2个位点的等位基因变体等方式显示在鱼体中[57]。将这3种鳜鱼的同工酶酶谱表达特征用带型码描述汇总(表1),这些具有鱼类种质特性的遗传生化标志可用来作为这3种鱼种质检测的重要参考指标。表1中除了肝脏的EST同工酶带型码较复杂外,其余的同工酶带型简单明了,条带清晰,种间差异明显,是较为实用的遗传性状标志。
4 小结
该研究采用非变性聚丙烯酰胺凝胶电泳检测技术,首次开展了杂交鳜5个组织3种同工酶酶谱特征的研究,并对斑鳜、翘嘴鳜3个组织3种同工酶酶谱特征进行了比较研究与分析。结果表明:不同地理种群斑鳜、翘嘴鳜的个别同工酶图谱存在一定的表达差异。杂交鳜F1代的3种同工酶具有不同程度的组织特异性,其中LDH同工酶、EST同工酶酶谱在5种组织中的表达特异性程度较高。杂交鳜F1代同工酶酶谱与双亲之间存在明显的亲缘关系和特异性,除了斑鳜和翘嘴鳜肾脏中的MDH同工酶谱以外,3个不同鳜种群间的酶谱特征均存在较明显的种属特异性,肝脏、肾脏中的LDH同工酶谱和EST同工酶谱、肌肉中的EST同工酶谱均存在显著且稳定种属差异表达。其中肝脏中EST同工酶谱较为复杂,其余的同工酶谱均简单明了,条带清晰稳定,种属差异显著,是较为实用的种质鉴定参考指标。通过分析斑鳜、翘嘴鳜及杂交鳜F1代的同工酶遗传特性,为种质鉴定工作提供了具有较高价值的生化遗传学资料,并为进一步开展斑鳜、翘嘴鳜的杂交育种工作提供了一定的理论支撑。
参考文献
[1]曾可为,王青云,高银爱,等.斑鳜的生物学及繁殖生物学的研究[J].当代水产,2005,30(2):21-23.
[2]宓国强,陈建明,练青平,等.杂交鳜与鳜鱼、斑鳜肌肉营养成分和氨基酸含量比较[J].水产养殖,2009,30(4):35-37.
[3]古丽.长珠杂交鳜[J].农村百事通,2018(3):29.
[4]许淼洋,郭金涛,赵金良,等.斑鳜(♀)×鳜鱼(♂)杂交子一代、子二代的形态特征分析[J].水产科学,2013,32(11):636-640.
[5]宓国强,练青平,王雨辰,等.杂交鳜胚胎发育观察[J].浙江海洋学院学报(自然科学版),2009,28(3):264-269.
[6]刘毅辉,陈永乐,朱新平,等.翘嘴鳜、斑鳜及其杂交后代的胚胎和胚后发育比较[J].大连海洋大学学报,2012,27(1):6-11.
[7]李燕,施顺昌,史建华,等.杂交鳜的胚胎发育形态观察[J].湖州师范学院学报,2014,36(8):19-24.
[8]袁勇超,梁旭方,田昌绪,等.翘嘴鳜、斑鳜杂交子代F1及其自交子代F2胚胎发育的研究及鉴定[J].湖北农业科学,2014,53(20):4920-4923.
[9]许淼洋.斑鳜(♀)×鳜(♂)杂交一代的胚胎发育、形态特征及肌肉生长相关基因表达研究[D].上海:上海海洋大学,2014.
[10]李燕,史建华,施顺昌,等.不同饲料对杂交鳜生长、成活率及体成分影响的比较[J].水产科技情报,2014,41(3):127-130.
[11]李传阳.不同种鳜鱼幼鱼摄食、消化能力和生长的比较研究[D].上海:上海海洋大学,2015.
[12]卢薛,古勇明,胥鹏.翘嘴鳜与斑鳜杂交的人工繁育技术[J].海洋与渔业,2015(8):60-61.
[13]高晓霞.“全雌翘嘴鳜已经培育成功,下一步攻克全雌杂交鳜”——访中山大学生命科学学院教授李桂峰[J].海洋与渔业,2017(9):46-48.
[14]陈俊豪,丁文岭,陈庆,等.翘嘴鳜“华康1号”苗种培育和成鱼养殖试验[J].科学养鱼,2018(1):40-41.
[15]李燕,史建华,李永强,等.翘嘴鳜、斑鳜和杂交鳜鱼体营养成分和氨基酸、脂肪酸组成的比较分析[J].水产科技情报,2015,42(5):246-250.
[16]宓国强,赵金良,贾永义,等.鳜(♀)×斑鳜(♂)杂种F1的形态特征与微卫星分析[J].上海海洋大学学报,2010,19(2):145-150.
[17]李传阳,许淼洋,赵金良,等.斑鳜(♀)×鳜鱼(♂)杂交后代遗传特征的微卫星分析[J].水产科学,2014,33(2):97-102.
[18]戴凤田,苏锦祥.鲿科八种鱼类同工酶和骨骼特征分析及系统演化的探讨(鲇形目:鲿科)[J].动物分类学报,1998,23(4):432-439.
[19]俞纯方,刘大翠,刘中全,等.鲿科四种鱼LDH同工酶谱的研究[J].四川职业技术学院学报,2003,13(1):86-88.
[20]方华华.江鳕和大头鳕形态学和遗传学的研究[D].青岛:中国海洋大学,2005.
[21]姬广磊,高天翔,柳本卓.黄海和日本海黄鮟鱇的形态和同工酶差异[J].海洋水产研究,2007,28(3):73-79.
[22]张媛.长江口刀鲚种群遗传结构分析[D].上海:上海水产大学,2005.
[23]李超,孙中武,王炳谦,等.芬兰虹鳟鱼同工酶分析[J].水产学杂志,2007,20(2):35-39.
[24]吴力钊,王祖熊.草鱼同工酶发育遗传学研究——Ⅰ.不同组织器官的同工酶分析[J].遗传学报,1987,14(4):278-286.
[25]庞秋香,庞书香,赵博生,等.泰山赤鳞鱼苹果酸酶同工酶表型差异的研究[J].安徽农业科学,2010,38(1):60-62.
[26]馬波,陈金平,董崇智.绥芬河三块鱼和珠星三块鱼种群的生化遗传变异及亲缘关系[J].中国水产科学,2005,12(6):688-693.
[27]葛学亮.唇(鲬骨)(Hemibarbus labeo)种群生化遗传学研究[D].哈尔滨:东北林业大学,2007.
[28]张林,周剑光,张涛,等.长江水系细鳞斜颌鲴形态特征及生化遗传特性分析[J].中国渔业质量与标准,2018,8(2):29-35.
[29]王小虎,叶玉珍,吴清江.鲤鲫人工多倍体谱系中同工酶和蛋白的基因表达[J].水生生物学报,2002,26(5):425-432.
[30]余波.虹鳟杂交及多倍体诱导的研究[D].天津:天津师范大学,2008.
[31]吴力钊,王祖熊.草鱼同工酶发育遗传学研究——Ⅱ.早期发育过程中的同工酶分析[J].遗传学报,1987,14(5):387-394.
[32]张庆朝,王慧,秦孜娟,等.泰山赤鳞鱼同工酶的研究[J].动物学研究,1994,15(2):62-67.
[33]付予昌.团头鲂胚胎发育过程及成体组织中八种同工酶系统的研究[D].武汉:中国科学院水生生物研究所,1985.
[34]方廖琼,何成明,刘宁.草鱼细菌性败血症乳酸脱氢同工酶的研究[J].水产养殖,1997(6):11-13.
[35]李政,王国良,金珊.患白云病加洲鲈的同工酶分析[J].水产科学,2005,24(4):21-23.
[36]王金秋,石椿.松江鲈鱼(Trachidermus fasciatus)不同组织同工酶的研究[J].复旦学报(自然科学版),2001,40(5):465-470.
[37]赵田田,蒲宗旺,岳兴建.花斑副沙鳅七种同工酶的组织特异性表达[J].湖北农业科学,2017,56(14):2736-2743.
[38]张涛,周剑光,吴金平,等.达氏鲟幼鱼形态特征及其同工酶电泳分析[J].中国渔业质量与标准,2018,8(5):9-17.
[39]段辛斌,陈大庆.淡水鱼类种质资源信息系统的研制[J].长江流域资源与环境,1999,8(1):57-62.
[40]张祖兴,李明云.大黄鱼种质资源研究进展[J].水产科学,2006,25(7):376-378.
[41]贾超峰,刘海林,许津,等.大黄鱼种质遗传多样性研究进展[J].海洋通报,2017,36(1):12-18.
[42]葛彦龙,尹洪滨,石连玉,等.鱼类同工酶电泳方法的改进[J].水产学杂志,2010,23(2):51-52.
[43]HJELMELAND K,RAA J.Characteristics of two trypsin type isozymes isolated from the arctic fish capelin(Mallotus villosus)[J].Comparative biochemistry & physiology part B:Comparative biochemistry,1982,71(4):557-562.
[44]吴鹤龄,林锦湖.遗传学实验方法和技术[M].北京:高等教育出版社,1983.
[45]刘志刚,卢迈新,曹建萌,等.罗非鱼“粤闽1号”及其繁育群体的遗传多样性和遗传关系分析[J].渔业科学进展,2018,39(6):31-41.
[46]李思发,蔡完其.团头鲂乳酸脱氢酶与苹果酸脱氢酶组织特异性的激光扫描分析[J].上海水产大学学报,1992,1(Z1):38-47.
[47]余来宁,夏小平,杨东,等.草鱼♀×鳡♂杂交F1代同工酶和蛋白质的电泳分析[J].安徽农业科学,2014,42(30):10573-10575.
[48]傅予昌,王祖熊.团头鲂的胚胎及成体组织中八种同工酶系统的研究[J].水生生物学报,1988,12(3):219-229.
[49]吴兴兵,许璞,戴卫平,等.江苏水域7种重要养殖鱼类的同工酶分析[J].南京师大学报(自然科学版),2007,30(1):96-101.
[50]杨军峰,周乔,邵雪玲,等.斑鳜和翘嘴鳜不同组织中过氧化物酶、酯酶和乳酸脱氢酶的比较研究[J].水利渔业,2007,27(6):7-9.
[51]李思发,王强,陈永乐.长江、珠江、黑龙江三水系的鲢、鳙、草鱼原种种群的生化遗传结构与变异[J].水产学报,1986,10(4):351-372.
[52]王丹,于伟君,唐作鹏.洄游型和陆封型香鱼同工酶的初步研究[J].水利渔业,1999,19(3):10.
[53]SHAKLEE J B,CHRISTIANSEN J A,SIDELL B D,et al.Molecular aspects of temperature acclimation in fish:Contributions of changes in enzyme activities and isozyme patterns to metabolic reorganization in the green sunfish[J].Journal of experimental zoology,1977,201(1):1-20.
[54]AHMAD R,HASNAIN A U.Ontogenetic changes and developmental adjustments in lactate dehydrogenase isozymes of an obligate airbreathing fish Channa punctatus during deprivation of air access[J].Comparative biochemistry & physiology part B,2005,140(2):271-278.
[55]李达,杨春,张力,等.鄱阳湖鳜鱼不同组织中乳酸脱氢酶同工酶的比较研究[J].淡水渔业,2002,32(6):41-43.
[56]MARKERT C L,FAULHABER I.Lactate dehydrogenase isozyme patterns of fish[J].Journal of experimental zoology,1965,159(3):319-332.
[57]WHITT G S.Development genetics of the lactate dehydrogenase isozymes of fish[J].Journal of experimental zoology,2010,175(1):1-35.
