KDM2A在结直肠癌中的表达及其与结直肠癌转移的关系研究
2020-12-08李晓伟崔立红
李晓伟,崔立红
解放军总医院第六医学中心消化内科,北京 100048
结直肠癌(colorectal cancer, CRC)是全世界最为常见的恶性肿瘤之一,根据2018年世界卫生组织最新的研究结果,CRC发病率在全世界所有恶性肿瘤中排名第3位,死亡率排名第2位[1-2];其发病率在我国排名第2位,死亡率排名第5位[3]。虽然CRC的发病率及死亡率在美国、日本、法国等发达国家呈下降趋势,但在我国均呈上升趋势[4]。转移是肿瘤患者死亡的主要原因,但因为CRC转移的机制目前不清,其疗效欠佳[5-6],因此阐明其转移的机制迫在眉睫。
表观遗传学参与了人体从胚胎发生到衰老的所有生物过程[7-14]。在核小体中,DNA包绕在两组核心组蛋白H2A、H2B、H3和H4周围,组蛋白氨基酸残基的多种不同翻译后修饰(例如甲基化、糖基化、乙酰化或磷酸化等)均可影响细胞的功能[15-16]。甲基化是组蛋白重要的修饰方式,可参与调节基因转录和DNA的损伤修复等多种过程,并与肿瘤发生、发展密切相关[17-19]。通常情况下,组蛋白H3赖氨酸K9、K27及K20等位点的甲基化与转录抑制相关,而组蛋白H3赖氨酸K4、K36及K79等位点的甲基化与转录激活相关[20]。在正常细胞中,组蛋白的甲基化一般处于稳定状态;在肿瘤细胞中,组蛋白甲基化失调并造成癌基因的转录激活及抑癌基因的转录抑制[21]。
KDM2A又名JHDM1A、FBXL11、Ndy2,是一种去甲基化酶,可使H3K36位点的单/双甲基化(K36 me1/2)去甲基化[20,22]。研究表明,KDM2A与多种肿瘤的增殖及转移相关,KDM2A在肺癌中高表达并可促进肺癌的增殖及转移[23];在肝癌中,KDM2A促进肝癌的起始及对索拉非尼的耐药[24];KDM2A可通过激活PI3K/AKT/mTOR/Vimentin通路促进卵巢癌的上皮间质转化(epithelial mesenchymal transition, EMT)及转移[25];KDM2A在胃癌中高表达并可促进胃癌的增殖及转移[26-27];KDM2A可促进CRC的增殖[28],但其与CRC患者预后的相关性及其是否参与调控CRC转移目前尚不清楚。
1 材料与方法
1.1 数据库介绍Oncomine数据库是一个汇集多个已发表文章的数据库,可用于研究目的基因在不同肿瘤组织及正常组织中的表达水平,并可研究目的基因的共表达基因。UCSC Xena是一个公共网站,可用于研究不同基因表达与肿瘤患者生存期的相关性,该网站包含了多个数据库的数据,本文我们利用TCGA数据库分析KDM2A表达与CRC患者生存期的相关性。C-bioportal网站包含了多个公开发表的数据库,可用于研究不同基因在多种肿瘤中的突变情况及该基因在不同肿瘤中与其他基因的共表达情况,本文我们利用TCGA数据库分析KDM2A与EMT相关调节蛋白在CRC中的表达相关性。CCLE是一个公共数据库,包含了多个系统肿瘤细胞系的测序数据,可用于分析靶基因在相应细胞系中的表达水平。
1.2 Real-time PCR采用Takara试剂盒提取相应细胞总RNA,对总RNA进行反转录及Real-time PCR实验,具体实验操作方法参考课题组既往已发表文章[29-31]。其中GAPDH为内参基因,相关基因mRNA表达量按2-ΔΔCt法计算。KDM2A及内参引物序列如下:KDM2A-F:5′-ATGGCTCGTCAATAGGCT-3′,KDM2A-R:5′-GGACTGATCTGTAAGGTGGC-3′;GAPDH-F:5′-CG-AACCTCTCTGCTCCTCCTGTTCG-3′,GAPDH-R:5′-CATGGTGTCTGAGCGATGTGG-3′。siKDM2A序列:siKDM2A-1:5′-AACAAGGAGAGUGUGGUGUUU-3′,siKDM2A-2:5′-GAACCCGAAGAAGAAAGGAUUCG-3′。
1.3 Transwell实验用胰蛋白酶分散SW620细胞,用含1% BSA的DMEM培养基重悬细胞。对于迁移实验,取1×105个细胞于200 μl含1% BSA的DMEM中,将该细胞悬液移至Transwell上室中,下室中加入含20% FBS的DMEM。对于侵袭实验,提前取PBS稀释后的Matrigel铺于上室中,实验前在上室中加入含1% BSA的DMEM培养基润洗Matrigel凝胶;上室中加入2×105个细胞(总体积200 μl),下室中加入含20% FBS的DMEM培养基。24 h后,弃培养基,擦去上层细胞,PBS清洗细胞,多聚甲醛固定后,结晶紫细胞染色,PBS清洗杂质,细胞风干后拍照,并进行统计分析。
1.4 统计学分析采用SPSS 20.0软件进行统计学分析,采用非配对t检验对计量资料进行比较,P<0.05为差异有统计学意义。
2 结果
2.1 KDM2A在CRC中的表达为研究KDM2A在CRC转移中的作用,我们首先用Oncomine数据库(GSE20916、GSE9348、GSE5206)分析KDM2A(探针:208987_s_at)在CRC组织及正常结肠黏膜组织中的表达水平,发现KDM2A在结直肠腺瘤及结直肠腺癌组织中的表达水平均高于正常结肠黏膜组织(P<0.05,见图1)。综上,本课题研究表明KDM2A在CRC组织中高表达。
2.2 KDM2A高表达与CRC患者生存期的关系KDM2A高表达的CRC患者总体生存期(P=0.064)、疾病特异性生存期(P=0.1053)、无进展生存期(P=0.013)、无疾病生存期(P=0.015)等均低于KDM2A低表达患者(见图2),提示KDM2A高表达患者预后差。
2.3 KDM2A促进CRC细胞的侵袭和迁移为研究KDM2A是否与CRC的侵袭及迁移相关,我们首先用CCLE数据库研究KDM2A在72种肠道肿瘤细胞系中的表达水平,结果表明,KDM2A在高转移潜能的SW620及LoVo细胞中的表达水平高于低转移潜能的HCT116及SW480细胞(见表1)。我们利用siKDM2A在SW620细胞中下调KDM2A的表达(见图3A),Transwell实验结果表明,KDM2A下调后SW620细胞的侵袭和迁移能力均受到抑制(见图3B~3C),提示KDM2A可促进CRC细胞的转移。
2.4 KDM2A诱导CRC细胞的EMT我们课题组前期研究结果表明,EMT对CRC转移至关重要[14],另有研究表明,KDM2A可通过激活PI3K/AKT/mTOR通路促进卵巢癌的EMT及转移[25]。我们利用TCGA数据库(n=526)分析了KDM2A与PI3K/AKT/mTOR等的表达相关性,发现KDM2A与mTOR(r=0.42,P=3.01E-24,见图4A)、N-cadherin(r=0.54,P=2.7E-40,见图4B)及Vimentin(r=0.27,P=4.9E-10,见图4C)在CRC中的表达呈正相关。提示KDM2A可能通过上调mTOR的表达促进CRC的EMT。
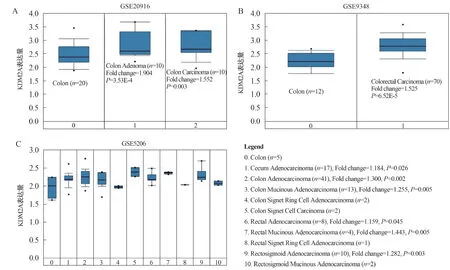
注:在GSE20916数据库(A)、GSE9348数据库(B)及GSE5206数据库(C)中,KDM2A(探针:208987_s_at)在结直肠腺瘤、结直肠腺癌及正常结肠黏膜组织中的表达。图1 KDM2A在CRC及正常结肠黏膜组织中的表达Fig 1 The expressions of KDM2A in CRC and normal colon mucosal tissues
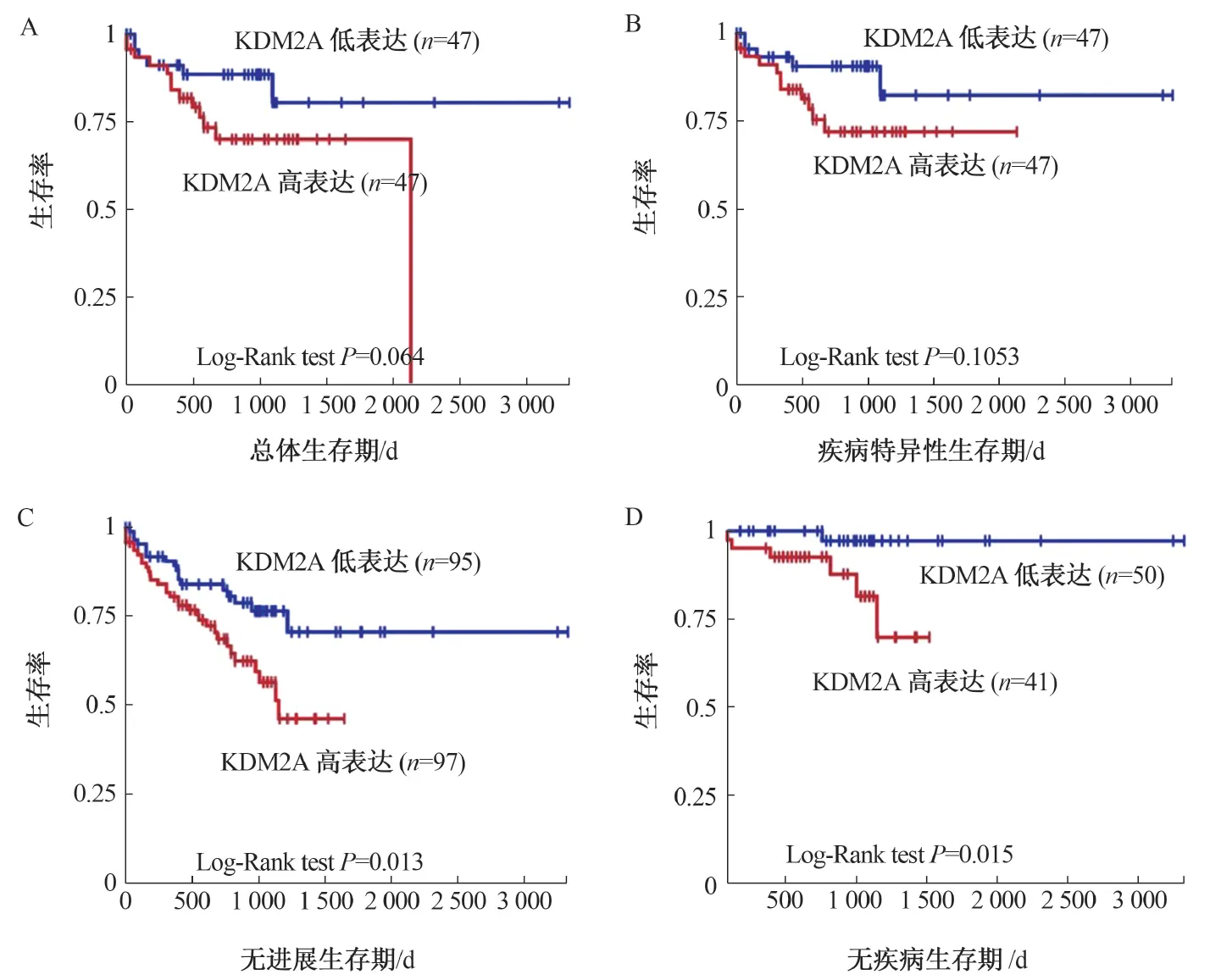
注:在TCGA数据库中,KDM2A表达与CRC患者总体生存期(A)、疾病特异性生存期(B)、无进展生存期(C)、无疾病生存期(D)的相关性。图2 KDM2A表达与CRC患者预后的相关性Fig 2 The correlation between KDM2A expression and the prognosis of the CRC patients

表1 KDM2A在CCLE数据库中72种肠道肿瘤细胞系中的表达水平Tab 1 The expression of KDM2A in 72 intestinal tumor cell lines in the CCLE database
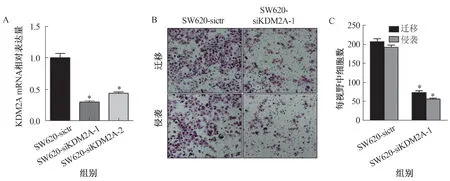
注:A:Real-time PCR实验用于比较KDM2A mRNA在相应细胞中的表达,与SW620-sictr组细胞相比,SW620-siKDM2A-1组及SW620-siKDM2A-2组细胞KDM2A表达量明显降低,*P<0.05;B:Transwell实验用于检测相应细胞的侵袭和迁移能力;C:Transwell实验的统计学分析,与SW620-sictr组相比,SW620-siKDM2A-1组细胞侵袭及迁移能力明显下调,*P<0.05。图3 KDM2A促进CRC细胞的侵袭和迁移Fig 3 KDM2A promoted the migration and invasion of CRC cells
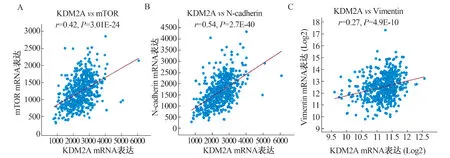
注:在CRC组织中KDM2A与mTOR(A)、N-cadherin(B)及Vimentin(C)的表达呈正相关(TCGA,n=526)。图4 KDM2A在CRC细胞中与EMT标志物的表达相关性Fig 4 The expression correlation between KDM2A and EMT markers in CRC cells
3 讨论
CRC是全世界最常见的恶性肿瘤之一。区别于西方发达国家,CRC发病率和死亡率在我国均呈上升趋势。CRC治疗主要有手术、化疗、放疗、靶向治疗、免疫治疗等,但因其发病机制不清,缺乏特效的靶向治疗。转移是CRC患者死亡的主要原因[5-6],阐明其转移的机制将对CRC的治疗至关重要。
我们课题组前期研究表明,BMI1-MEL18通路与CRC的发生相关[32],近年来研究表明,赖氨酸去甲基化酶KDM2B可促进靶基因对BMI1及MEL18招募[33],且可通过调节BMI1的表达促进CRC的EMT[34]。我们课题组前期研究结果表明,SIBO阳性患者结肠息肉及CRC发病率高于SIBO阴性患者,且SIBO阳性患者结肠黏膜KDM2A表达量高于SIBO阴性患者,KDM2A是KDM2B同家族蛋白,均可使组蛋白H3赖氨酸K36位点的单/双甲基化(me1/2)去甲基化[20,22],既往研究表明,KDM2A可促进胃癌[27]、肺癌[23,35]、骨肉瘤[36]、宫颈癌[37]、胶质瘤[38]、乳腺癌[39]、卵巢癌[25]等肿瘤的转移。在CRC中,KDM2A可促进肿瘤细胞增殖[28],但KDM2A是否与CRC转移相关目前尚不清楚。本课题发现KDM2A mRNA在CRC组织中高表达,其高表达提示预后差;通过体外细胞功能学实验我们发现,KDM2A表达下调可抑制SW620细胞的运动能力,提示KDM2A在CRC中高表达并能促进CRC转移。
我们课题组前期研究结果表明,EMT与CRC转移密切相关[14],且多项研究表明,KDM2B可通过诱导细胞的EMT促进多种肿瘤的转移[34,40],KDM2A可通过激活PI3K/AKT/mTOR通路促进卵巢癌的EMT及转移[25],但KDM2A促进CRC转移是否与EMT相关尚不清楚。本课题发现KDM2A与mTOR、Vimentin及N-cadherin在CRC中的表达正相关,提示KDM2A通过调节CRC的EMT促进其转移。但KDM2A是否通过上调mTOR的表达促进CRC转移需要更多实验验证。
在肝癌中,KDM2A可通过使NANOG、SOX4及OCT4启动子区域H3K36位点发生去甲基化,维持肝癌肿瘤干细胞的干性及对索拉非尼的耐药[24]。在骨肉瘤中,KDM2A可通过促进p21及puma启动子区域H3K36me2的去甲基化,促进骨肉瘤细胞的增殖。但KDM2A促进CRC的转移是否通过其去甲基化功能需要更多实验验证。
总之,本研究发现KDM2A在CRC高表达并提示预后差,KDM2A通过诱导CRC发生EMT进而促进CRC的转移,靶向KDM2A通路可能作为CRC转移的新思路。
