UV-B辐射增强对粳稻营养生长时期叶片光合特性的影响
2020-12-04战莘晔吕晓金丹丹殷红
战莘晔,吕晓,金丹丹,殷红
(1.鞍山市气象局,辽宁 鞍山 114004;2.沈阳农业大学农学院,沈阳 110866;3.锦州市生态与农业气象中心,辽宁 锦州121000;4.辽宁省农业科学院植物营养与环境资源研究所,沈阳110161)
大气平流层当中臭氧(O3)衰减引起的地表UVB 辐射增强是全球气候变化问题之一。太阳紫外辐射到达地面需通过平流层中的O3层进行过滤,O3衰减乃至空洞的出现,致使到达地面的太阳紫外辐射增强。据北京监测数据显示,我国大气平流层的O3衰减情况也很严重,平流层O3总量呈降低趋势,我国各地平均降低5.1%[1-2]。研究表明,大气平流层O3每减少1%,到达地面的太阳紫外辐射增加2%[3-4]。UV-B辐射(280~320 nm)是可见光的天然成分,对植物具有重要的生物学作用[5-6]。Dehariya 等[7]认为,增强的UV-B 辐射可以改变植物的光合作用、水分的代谢以及碳从生长池分配到二级代谢的途径,且外观形态上与在UV-B 环境下生长的植物相比,在没有紫外辐射下的叶片生长速度更快、叶片更大。
有研究者认为当发生常规光合作用时,激发态的光子传递到光系统Ⅱ(PSⅡ)后使光反应中心P680发生电荷分离,产生光合电子流,且光合电子流越大,表明其光合能力越强[8-10],但当受到外界UV-B 胁迫的时间和强度增加时,光合反应中心的开放程度以及光合效率则会降低[11]。在研究水稻等C3植物过程中,曾由Farquhar 等[12-13]提出的FvCB 模型因能描述稳态的C 同化过程且具有明确的生物学意义而被广泛应用于光合作用研究[14]。用该模型可描述植物光合作用中如最大羧化速率、最大电子传递速率等几个重要参数[15-16],但叶子飘等[17]在估算植物叶片电子最大传递速率以及计算具体的C 同化和光呼吸所分配的光合电子流时,估算值高于FvCB 生化模型的测算值,因此本文采用叶子飘等[17]的研究方法进行估算。
近些年,研究者们常通过研究叶绿素a 快速荧光诱导动力学参数的变化进而研究外界胁迫对植物光合作用的影响[18]。叶子飘等[19]利用快速荧光诱导动力学技术,发现UV-B 辐射增加使绿豆幼苗的光合作用效率降低,捕光复合体和电子传递系统被损坏,电子传递速率下降。王伟等[20]在实验中增设UV-B 辐射后,紫花苜蓿的各项荧光参数均降低,表明:一是紫花苜蓿叶片的光合器官活性受UV-B 胁迫后降低,进而对C 的同化和固定减少,导致光能过剩;二是植物叶片光系统PSⅡ为维持自身运转,通过减少光合电子传递方式来调节对C 的代谢力,这与王锦旗等[21]的研究结果相一致,且有研究发现当PSⅡ反应中心开放程度变大时,光化猝灭也会增大[22]。而韩文等[23]研究表明,短时间的UV-B 辐射对拟南芥叶绿素荧光特性有影响,原初光能转化效率、光化学猝灭系数等指标降低,NPQ 值则呈上升趋势。研究认为,只有经过长时间的UV-B辐射PSⅡ反应中心才会受创,从而影响光合电子传递,而短时间辐射对PSⅡ反应中心影响较小。
已有研究表明,作物对UV-B 辐射增强表现敏感,株高、叶面积、干物质量等均有降低表现[24],水稻产量与品质以及有效穗数和结实率显著下降,叶绿素含量与光合速率也有所降低[25-27]。但针对UV-B 辐射对北方粳稻光合特性的研究还相对较少,尤其是针对营养生长时期,因此本文以东北粳稻为受试材料,探究了营养生长时期UV-B 辐射增强对粳稻光化学活性和光合电子流传递分配等方面的影响机制。
1 材料与方法
1.1 试验材料与方法
以粳稻品种‘沈农265’为试验材料,于2014 年5—10 月在沈阳农业大学水稻研究所试验田进行,试验布局如图1 所示。采用盆栽方式种植,并选用南地大田水稻土作为生长土壤,土壤有机质含量33.8 g·kg-1,速效钾含量155.7 mg·kg-1、速效磷含量138.2 mg·kg-1、速效氮含量187.8 mg·kg-1。设置对照组CK(自然光照射)、处理组T1(自然光照射+UV-B 辐射1.05 W·m-2)和T2(自然光照射+UV-B 辐射2.1 W·m-2)。夏季自然光照实际日均UV-B 辐射为21 W·m-2数据来源于沈阳气象局,即T1 和T2 处理分别增加了实际辐射的5%和10%,各组均种植42 盆,每日8:00起连续照射8 h。移栽后每隔20 d对叶片进行一次活体测量和取样,每处理重复3次。
图1 试验布局图Figure 1 Experimental design layout
1.2 测定项目与方法
选取水稻所培育的三叶期幼苗移栽于无孔试验盆中,试验盆上直径0.33 m、下直径0.20 m、盆深0.29 m,插秧时3 穴·桶-1,1 株·穴-1,对照组CK 和处理组T1、T2 各42 桶,并进行常规水肥管理。架设北京电光源研究所生产的紫外灯管于幼苗上方,每个灯架配有防雨灯罩,且呈东西方向摆放,以减少遮阴效果,灯架高度可调整,始终与植株保持0.3~0.4 m 高度,以保证植株受到相同剂量的UV-B辐射,每日8:00-16:00开启紫外灯照(阴雨天除外)。
在营养生长3 个测定日期即移栽后第20、40、60 d(孕穗期前),在CK和处理组中各随机选出6株植株做标记。选择晴朗午前进行活体测量,利用暗适应夹进行30 min暗适应后,测定荧光诱导动力学曲线及其参数;用Li-6400 型光合测定仪测定净光合速率(Pn)和暗呼吸速率(Rd),选择人工红蓝光源,光强设定为1 000 μmol·m-2·s-1;利用FMS Ⅱ型便携式荧光测定系统(Hansatech,UK)测定同期荧光参数,暗反应参数则需叶片暗适应30 min后测定,测量时设定光照强度为1 000 μmol·m-2·s-1。每处理重复3次。
1.3 模型应用与数据处理
1.3.1 模型简介
计算分配到碳同化和光呼吸光合电子流的模型[28]。
(1)光合电子传递活性:通过PSⅡ的非环式光合总电子传递速率:JF=ΦPSⅡ×I×a×f=rETR;
参与光呼吸的电子流:Jo=2/3[JF-4(Pn+Rd)];参与碳还原的电子流:Jc=1/3[JF+8(Pn+Rd)];PSⅡ实际光化学量子效率:ΦPSⅡ=(Fm′-Fs)/Fm′。
(2)Rubisco 的氧化速率(Vo)和羧化速率(Vc):JF=4Vc+6Vo,Vc=Pn+Rd+0.5Vo。
其中ΦPSⅡ为实际光化学量子效率;I 为入射叶面光强;a 为叶片的光吸收系数;f 为光能在光系统Ⅱ中所占比;rETR 为光电子传递相对速率;Pn为净光合速率;Rd为暗呼吸速率;Fm′为光下最大荧光;Fs为光下稳态荧光。
1.3.2 荧光诱导动力学参数的计算
荧光诱导动力学参数及计算方法见表1。

表1 叶绿素荧光动力学参数Table 1 Parameters of fluorescence transient
1.3.3 统计分析
数据采用SPSS 17.0 进行方差分析,采用LSD 法对数据进行多重比较,显著性检验在0.05的水平下进行,并运用Office 2003制图。
2 结果与分析
2.1 UV-B辐射增强对光合电子传递活性的影响
在营养生长期,参与光呼吸的非环式电子流传递速率Jo 在UV-B 处理下,T2 对Jo 的抑制虽然大于T1,但降低趋势不显著。参与碳还原的非环式电子流传递速率Jc,在T1 处理下只有移栽后60 d 表现出显著下降趋势,较CK下降了9.44%,但在T2处理下移栽后20 d 降低13.64%,移栽后40 d 下降了10.54%,60 d 时降低了18.3%,整个营养生长期都呈现出高剂量UV-B辐射下的Jc明显小于CK的趋势,说明UV-B增强抑制了光合电子向光化学方向的传递。参与光呼吸的电子流比例Jo/JF,在T1 处理下移栽后60 d 较CK增加了3.52%,在T2 处理下移栽后40 d 和60 d 较CK分别增加了3.41%和7.16%,说明营养生长期随着UV-B 辐射的增强,总电子流参与光呼吸的比例也增大。光呼吸和碳还原方向的光合电子分配比例Jo/Jc在营养生长期随UV-B辐射增强呈增加的趋势,在T1处理下移栽后60 d增加了8.42%,在T2处理下均呈增加趋势,且在移栽后60 d 增加了19.78%,增幅最大,说明UV-B 辐射增强随着辐射时间增加,光合电子的光呼吸消耗增加,碳还原过程被减弱。
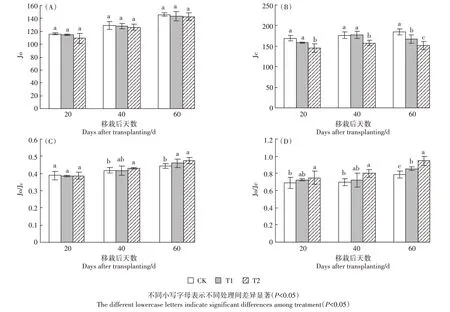
图2 UV-B辐射增强对粳稻Jo、Jc、Jo/JF、Jo/Jc的影响Figure 2 Effects of enhanced UV-B radiation on Jo,Jc,Jo/JF,Jo/Jc in Japonica rice
2.2 UV-B 辐射增强对Rubisco 氧化速率和羧化速率的影响
Rubisco氧化速率Vo在营养生长期UV-B处理下变化趋势不显著。Rubisco羧化速率Vc,在T1处理下移栽后60 d 较CK 显著降低了19.49%,T2 与CK 在第20、40、60 d 均达到显著差异(P<0.05),分别下降了10.74%、19.55%和31.17%,说明随着UV-B 辐射的增强,光合电子进入Rubisco羧化反应受到了抑制,羧化传递速率显著降低。专一因子用Vo/Vc 表示,即叶片Rubisco 氧化速率/羧化速率,在营养生长期,T1 处理下移栽后60 d 较CK 显著增加了20.3%,T2 处理下移栽后40 d 和60 d 较CK 分别显著增加了34.21%和48.02%。
2.3 UV-B辐射增强对荧光诱导动力学曲线的影响
OJIP 曲线一般分为O、J、I、P 四相特征点位,分别表示10~50 μs、2 ms、30 ms、0.3~2 s时的荧光。由图4可以看出:移栽后20 d(图4A),CK 和处理组相比在整个阶段均表现出一些差异,而在移栽后40 d(图4B)和60 d(图4C)影响较小;移栽20 d 后J-I 阶段表现出明显的CK>T1>T2,说明高剂量UV-B 辐射降低了快还原型PQ 库数量,其他阶段没有表现出明显特征;在I-P 段,移栽后20 d 和60 d 表现出明显的CK 大于处理组,说明UV-B 辐射抑制了慢还原型PQ 库的还原,而P点(0.3~2 s间的峰值)呈现最明显的变化出现在移栽后60 d,此时CK明显大于处理组,说明即使在短暂的UV-B 辐射下P 点没有明显变化,但是随着时间推移,最终会降低荧光产量。
2.4 UV-B辐射增强对荧光动力学参数的影响
随UV-B 辐射的增强,Fv/Fm 在移栽期后的40、60 d均呈减小趋势,且CK大于处理组,移栽后20 d变化不显著;移栽后40 d和60 d均显著降低(图5A),说明UV-B 辐射的增强能够降低粳稻原初光能转换效率。随着UV-B 辐射增强,qP 在T2 处理下移栽20 d后显著降低了8.98%(图5B),但在移栽后40 和60 d没有显著降低。
随UV-B 辐射的增强,在T2 处理下移栽后40 d和60 d NPQ 极显著增加了56.13%和48.8%(图5C),说明UV-B 辐射的增强,抑制了粳稻光化猝灭,促进了非光化猝灭作用。随UV-B 辐射的增强,T2 处理下3 个测定时期的rETR 均显著降低(图5D),呈现出CK>T1>T2 的趋势。移栽后20 d,T1、T2 分别降低了3.77%和4.53%;移栽后40 d T2处理降低了8.76%;移栽后60 d,两处理分别降低了4.32%和9.4%,说明UV-B 辐射越强,光合电子传递的相对速率越低,T2对于rETR的抑制作用大于T1。
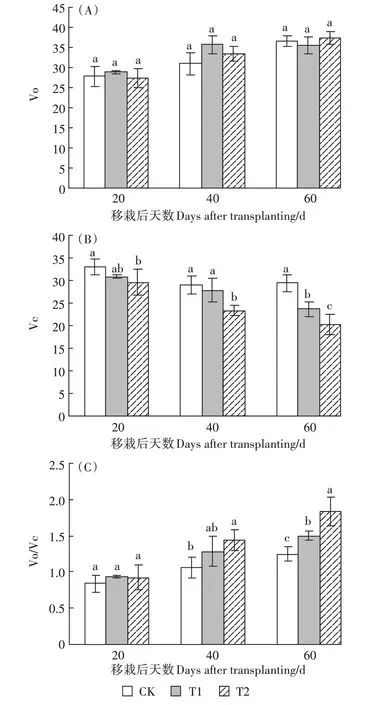
图3 UV-B辐射增强对粳稻Vo、Vc、Vo/Vc的影响Figure 3 Effects of enhanced UV-B radiation on Vo,Vc,Vo/Vc in Japonica rice
3 讨论
光合电子流是C 同化的动力来源,也是光合能力的基础[28]。本试验采用的粳稻为C3植物,在C3植物中,光呼吸是非环式光合电子流分配的主要途径,同时也是重要的C 代谢途径之一[29],光合电子流分配是植物光合系统的重要部分[30],提高C3植物的光合效率,减少光合产物消耗的主要影响因素是光呼吸活性,通过对各类环境中各种植物的光合电子流传递分配规律的探索有助于诠释光合机理与传递模式[31]。因此,在本实验中,通过增设UV-B 辐射,探讨分析了粳稻营养期叶片的光合电子流分配规律。杜清洁等[32]在试验中发现,番茄叶片在受干旱与亚低温共同胁迫下PSⅡ总电子流及流向各库的电子流均下降,包括光合碳同化的电子流占总电子流比例,此外其他部位电子流比例均上升,且分配给光呼吸的电子流比例增幅最大,表明受到胁迫时光呼吸可作为主要的电子流库,耗散过剩光能。这与本试验得出的结论一致,在本试验中,Jc 降低,但Jo/Jc 增加,表明UV-B 抑制了营养时期非环式光合电子流参与碳还原的比例,而增加了光呼吸电子流占比,同时Vc降低,专一因子Vo/Vc 却有所增长,表明对向羧化方向传递的光合电子有抑制作用。
叶子飘等[33]和薛娴等[34]在植物光合原初反应过程中发现,光合电子流是经捕光色素分子采集光能后,在PSI和PSⅡ之间传递所产生的电荷形成的电子汇集而成。这些电子经由电子受体流转,最终被传递到NADPH 和ATP 中,此时在为碳同化和光呼吸提供能量的同时完成了光能到化学能的转换。余小龙等[35]认为细叶青冈的叶绿体PSⅡ反应中心如果受到损伤,从PSⅡ反应中心到QA、QB和PQ 的电子转移会受到一定的限制,进而影响反应中心色素的激发能转移。本试验中,在UV-B处理20 d后降低了Pheo将电子传给QA,QA-传递给QB的过程,即快还原型PQ 库的数量。李晓科等[36]认为光合速率下降的原因可能是高强度的UV-B 辐射导致大麦幼苗叶片叶绿素遭到破坏致使含量降低,并且光合酶如PEP 羧化酶、RuBP羧化酶活性下降,光系统受到影响,特别是捕光色素,最终使叶绿体吸收的光能降低。
张玉红等[37]和李雪莹等[38]在研究中发现,被测作物原初光能捕获效率及光合电子传递效率光化猝灭系数等受UV-B 胁迫后显著降低,这与本试验中3 个测定时期Fv/Fm、rETR、qP 均与UV-B 强度的增大而呈负相关结论一致。祁虹等[39]在研究棉花时发现,随着UV-B 辐射增强,叶片中NPQ 与之呈正相关增强,这与本研究结论一致,在本试验中3 个测定时期NPQ都随UV-B辐射的增强而增少,表明PSⅡ反应中心受损,光化学效率降低,用于光合作用的激发能比例降低,NPQ的提高反映出一种自我保护机制。
4 结论
在营养生长期UV-B 辐射增加总体上会增加叶片的光呼吸耗能,降低叶片对光的利用能力,进而影响植株粳稻光合性能。
(1)在营养生长期UV-B 增强总体上对光合电子的Rubisco 氧化速率起到了推进作用,光合电子进入Rubisco 羧化反应受到抑制,同时专一因子Vo/Vc 也呈上升趋势,说明UV-B 辐射的增强迫使Rubisco 氧化反应消耗更多的光电子,从而没有更多光电子用于羧化反应。
(2)UV-B 短期处理会影响OJIP 整个阶段,随着辐射时间增加,处理间差距缩小,最后测量阶段荧光产量的明显降低说明UV-B 辐射最终会影响营养生长时期叶片的相对荧光产量。
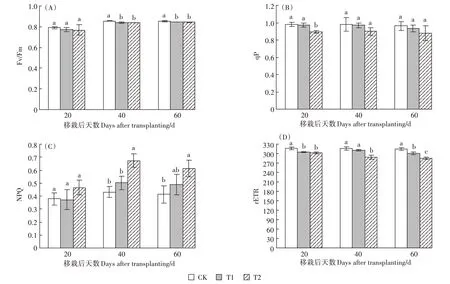
图5 移栽后荧光动力学参数的变化Figure 5 The changes of basic parameters on fluorescence after transplanting in Japonica rice
(3)UV-B 辐射的增强会使移栽期后3 个测定时期的Fv/Fm、qP、rETR 减小,但NPQ 升高,说明UV-B辐射的增强,抑制了营养生长期叶片的光化猝灭速度,却促进了非光化猝灭速度,同时光电子传递速率也随之越低。
