西藏色季拉山森林群落沿海拔梯度变化格局
2020-12-01刘丽娟方江平
高 辉, 刘丽娟, 方江平
(1. 西藏农牧学院 高原生态研究所, 西藏 林芝 860000; 2. 西藏高原森林生态教育部重点实验室(西藏农牧学院),西藏 林芝 860000; 3. 西藏林芝高山森林生态系统国家野外科学观测研究站(西藏农牧学院), 西藏 林芝 860000;4. 西藏自治区高原森林生态实验室(西藏农牧学院), 西藏 林芝 860000; 5. 西藏自治区生态安全联合实验室(西藏农牧学院),西藏 林芝 860000; 6. 西华师范大学 环境科学与工程学院, 四川 南充 637002)
随着全球生物多样性丧失的不断加剧, 保护生物多样性成为生态学领域研究的热点内容[1]。生物多样性形成和维持很大程度上依赖于群落构建:生物多样性在一定程度上可以反映群落组成和结构沿着环境梯度的变化,如果单纯考虑物种多样性的不足,不同物种在群落中的生态适应性以及它对生态系统功能的影响就很难体现出来,所以在探讨生态系统功能和生物多样性之间的联系时人们往往会从更宽的维度范围来考虑[2]。通常情况下,生态位理论认为共存物种间的生态位分化等相对较为确定的因素主导群落的构建过程[3]。但从生态位的角度来考虑,我们认为具有相似的生理生态性状的物种生态位发生了重叠;而中性理论认为不同物种具有同等的生态学特性。生境条件相同,随着物种空间距离的增大,距离衰减效应则占据主导因素,这主要是由于遗传交流的传播受到一定的限制,这种特点给生态位理论的研究提供了全新的思路[4-5]。群落的构建机制只有通过整合生态位理论和中性理论共同探讨才具有全面性[6],目前关于群落构建机制的探究大多从物种维度展开[7-8]。物种维度(物种多样性、系统发育多样性和功能多样性)研究是基于物种是独立的进化和功能单元前提条件建立的,然而生态群落中的不同物种在进化史上具有紧密联系,现有生态圈的和谐共存是生态过程和协同进化过程的共同作用来构建起来的。Webb等[9]提出可以通过分析群落内物种的亲缘关系进一步推断群落构建的成因;也有学者提出生态位保守理论[10],并提出全新的观点——系统生态过程和发育结构的逻辑框架结构。生态位保守理论认为,种间的系统发育相似性可以用来代替种间的生态相似性,功能性状的相似程度与亲缘关系的相近性具有密切关系[10]。由于物种功能性状随着物种不同其保守程度一般情况下也不尽相同,物种间的功能性状高度分化是主要的决定因素,所以通常情况下不能仅仅依靠物种间的系统发育关系作为物种生态相似性的代表因素[11-12]。
群落系统发育α及β多样性对探究生态位和中性过程在群落构建中的作用具有重要影响[13-16],群落系统发育和功能性状的α及β多样性与研究尺度同样具有密切的关系[17]。现有研究表明,物种间的扩散限制和相互作用一般情况下发生在小群落尺度层面[18],环境过滤作用则发生在大群落尺度层面。通常较大的乔木更易发生竞争排斥作用,主要是由于其对更多生境资源的需求,所以群落系统发育的α多样性呈现出发散格局[19-20]。具有较为明显的尺度依赖性是β多样性格局的显著特点。β多样性的形成机制研究现在还不成熟,存在多种理解角度。基于中性理论的扩散限制作用和基于生态位理论的生境过滤作用是目前比较热门的研究方向。物种多样性研究通常会涉及群落系统发育,而群落成分的空间变化研究却较为全面[21-23]。例如:文献[24]从生物群落的系统发育角度来推测物种多样性沿海拔变化的驱动因素;Bryant等[25]发现北美落基山脉沿海拔梯度的变化格局表现为聚集类型,而植物群落表现为发散,同时发散程度随着海拔升高越来越显著。这种发现揭示了物种间存在较为明显的竞争作用[26],说明非随机过程对群落构建起着非常重要的作用,这也为探讨生物多样性格局沿海拔梯度的群落构建、物种的维持机制与生态环境适应性提供了全新的研究视角。
色季拉山位于西藏东南部原始林区的核心林区,地处雅鲁藏布江中游,山体较大,海拔梯度明显,是高山森林生态系统研究首选的典型地区[27]。本文以对环境变化极为敏感的色季拉山森林群落为研究对象,开展森林群落系统发育研究,通过分析不同植被类型的森林群落系统发育结构和沿海拔梯度的变化,试图探究:1)色季拉山沿海拔梯度不同生活型(乔木、灌木和草本)植物的系统发育α及β多样性如何变化?2)空间变量和环境变量如何影响系统发育的α及β多样性?从而推测西藏色季拉山群落构建的生态学过程和气候变化对物种共存的影响机制。
1 材料与方法
1.1 研究区概况
色季拉山(93°12′—95°35′E,29°10′—30°15′N)位于西藏林芝市雅鲁藏布江西北侧,念唐古山与喜马拉雅山脉接合处,处于西藏东南湿润区和半湿润区的过渡核心地带。
1.2 研究方法
1.2.1 样方设置和物种收集
根据研究地区地形地势以及色季拉山群落结构特征,样地的设置采用梯度样带网格法[28],即在海拔3 000—4 700 m研究区域内按垂直高度100 m分阳坡和阴坡设置典型样地,每个海拔梯度设置2个典型样地,样地大小为30 m×30 m,共设置阴坡36块样地,阳坡36块样地,合计72块样地。
将30 m×30 m的样地平分为9个10 m×10 m的网格样方,每个样方选取一个固定位置设置5 m×5 m灌木样方和1 m×1 m草本样方。对样地内乔木进行每木检尺和树木定位调查,测定其胸径、株高、株数,并记录物种名称;对灌木进行多样性指标统计,对草本进行多度统计和盖度估算;对整个样方的坡度、坡向、干扰程度、郁闭度和环境因子进行测量并记录。
1.2.2 群落系统发育多样性分析
用系统发育α多样性Faith’s PD指数和系统发育β多样性PhyloSor指数分析色季拉山群落的系统发育多样性,并在R软件进行计算。
Faith’s PD指数是样地中出现的所有分类群对应的最小生成路径支长的和,反映了该区域物种组成的系统进化特征多样性。同时采用随机系统发育关系的零模型同PD指数值进行参考比较,其结果可以表明群落的系统发育多样性的类型。
PhyloSor指数是2个群落共有物种的枝长与2个群落所有物种的枝长和的比值。比值较低,说明涉及到的两群落间的相似性就越小;反之,相似性越高。其计算公式为
(1)
式(1)中:LBLi为群落i内所有物种系统发育结构的枝长和;LBLj为分群落j内所有物种系统发育结构的枝长和;LBLi,j为2个群落内共有物种系发育结构的枝长和。
(2)
式(2)中:C是研究区域所有分类群构成的系统进化树;c是C上的一个分支(即2个节点间的片段);Lc是c的支长。
再采用最近邻体距离(nearest neighbor distance,D′nn)和平均成对距离(mean pairwise distance,D′pw)指数进一步衡量群落间随海拔变化的系统发育非相似性,计算公式为:
(3)
(4)
式中:min(δjk1)是指群落k2中物种j和群落k1中所有物种的最近邻体距离;min(δik2)是指群落k1中物种i和群落k2中所有物种的最近邻体距离;mean(δjk1)是指群落k2中物种j和群落k1中所有物种的平均成对距离;mean(δik2)是指群落k1中物种i和群落k2中所有物种的平均成对距离;fi是指群落k1中物种i的相对多度;fj是指群落k1中物种j的相对多度。标准化的S.E.S. D′nn和S.E.S. D′pw值利用零模型产生的D′nn 指数以及D′pw指数与实际观测到的D′nn 指数以及D′pw指数对比产生,以便用于对群落间的系统发育的非相似性是否为随机构成作出观测分析。其中零模型的确定方法是将样地中各个物种出现的频率设定为常数,其中物种是否会同时出现使用独立置换法被随机化99次后建立起零模型。
1.2.3 群落系统发育结构
通常情况下,在进行相关群落内物种间的亲缘关系测定时, 一般用系统发育关系最近的物种之间的距离(mean nearest taxanodal distance,MNTD)结合物种间的平均系统发育距离(mean pairwise nodal distance,MPD)来体现,但往往计算时会将MNTD和MPD换成最近种间亲缘关系指数(INT)和近亲缘关系指数(INR)。INT和INR的计算公式为:
(5)
(6)
式中:DMNTDO和DMPDO为实际观察的MNTD值和MPD值;DMNTDR和DMPDR为随机产生的999个零群落的MNTD值和MPD值;mean(DMNTDR)和mean(DMPDR) 为零群落MNTD值和 MPD 值的平均值;sd(DMNTDR)和 sd(DMPDR)为零群落MNTD值和MPD值的标准差。
使用Studentt进行检验,如果实际观测值与期望值间的差为正,说明该群落的系统发育情况为发育聚集;如果差为负,则该群落的系统发育情况为发散。
1.2.4 群落系统发育结构的冗余分析
采用R软件进行冗余分析。冗余分析及显著性的解释部分通过vegan软件包来实现,PCNM变量通过SpaceMaker软件包实现,前向选择通过packfor软件包实现。分析过程中采用前向选择法(n=9 999)筛选出空间和环境变量中的显著因子作为解释变量,同时对系统发育结构中INT和INR进行变差分解的分析探究。
2 结果与分析
2.1 系统发育α多样性沿海拔梯度的变化格局
由图1结果可知,色季拉山群落系统发育α多样性指数与物种丰富度变化呈现出典型的正相关关系。从观测值与零模型比较结果看:乔木和灌木未表现出显著差异;草本观测值与零模型预期Faith’s PD指数间的差异随着物种丰富度的增大越来越显著,整个变化呈现非随机的变化过程。综上可知,不同物种类型在色季拉山森林群落中的构建机制不同。
2.2 系统发育β多样性沿海拔梯度的变化
由图2分析可知,乔木、灌木和草本的PhyloSor指数均随着海拔变化形成的空间距离的增大而呈现下降趋势,2个变量之间表现出典型的负相关关系,表明色季拉山群落间系统发育相似性随着空间距离增加逐渐降低。这种现象主要是由于生境过滤或者竞争排斥引起的。综上表明,随海拔差异的增大,色季拉山群落系统发育β多样性会降低。
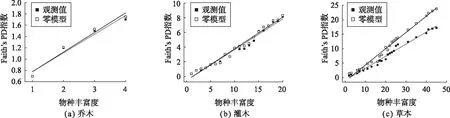
图1 色季拉山系统发育α多样性Faith’s PD指数(观测值vs零模型)沿物种丰富度的变化Fig. 1 Variation of phylogenetic diversity Faith’s PD index (observed value vs zero model) along species richness in Sejila Mountain, Tibet
由图3分析可知,色季拉山系统发育的平均成对距离(S.E.S.D′pw)和最近邻体距离(S.E.S.D′nn):对草本而言,沿海拔距离差的系统发育β多样性表现出一定程度的相似性,在海拔跨度较小的范围区间内比预期系统发育β多样性转变得慢一些;随着海拔跨度的逐渐增大,群落的系统发育相对比预期明显加快。乔木在海拔距离差为400、600、800 m时,灌木在海拔距离差为400、600 m时,草本在海拔距离差为200、400、600 m时,系统发育β多样性的标准化值接近零,表明群落在此海拔跨度呈现的是随机分布格局。
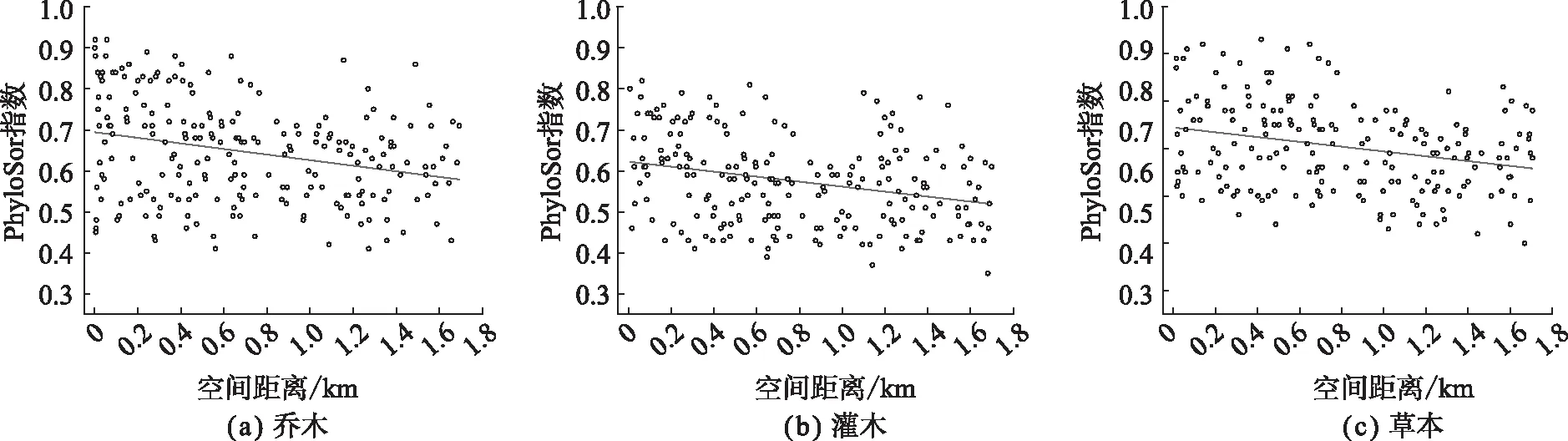
图2 色季拉山系统发育多样性PhyloSor指数沿海拔距离差的变化趋势Fig. 2 PhyloSor index of phylogenetic diversity along the difference of altitude distance in Sejila Mountain, Tibet

图3 色季拉山标准化的群落系统发育平均成对距离(S.E.S.D′pw)和最近邻体距离(S.E.S.D′nn)沿海拔距离差的变化Fig. 3 Variation of mean pair distance (S.E.S.D′pw) and nearest neighbor distance (S.E.S.D′nn) of standardized community phylogeny along altitude distance in Sejila Mountain, Tibet
2.3 群落系统发育结构与环境空间变量的关系
结合表1和图4可知,影响色季拉山乔木沿海拔梯度系统发育的近亲缘(NRI)的主要环境因子是土壤中的有效氮含量和坡度因素;坡向是影响灌木沿海拔梯度系统发育的近亲缘(NRI)的主要环境因子;土壤中有机碳含量是影响草本植物沿海拔梯度系统发育NRI的主要环境因子。

表1 前向选择法筛选影响色季拉山海拔梯度群落系统发育结构(NRI)的环境变量和空间变量

a表示乔木,b表示灌木,c表示草本。图4 影响不同生活型色季拉山植物群落系统发育结构(NRI和NTI)的环境变量和空间变量筛选结果Fig. 4 Screening results of environmental and spatial variables affecting phylogenetic structure (NRI and NTI) of plant communities of different life types in Sejila Mountain, Tibet
表2为前向选择法筛选结果,对于环境变量来说,空间变量同样显著影响着乔木、灌木以及草本的系统发育结构。最近亲缘关系指数(INT)的R软件冗余分析结果表明,空间变量在色季拉山乔木、灌木和草本的群落系统发育结构均表现出显著的影响结果,而环境变量方面没有筛选出显著的影响因子。可见,不同群落系统发育结构指数对于空间变量和环境变量的响应是不一样的。

表2 前向选择法筛选影响色季拉山海拔梯度群落系统发育结构(NTI)的环境变量和空间变量
3 讨论
生态位的分化过程常常用竞争排斥和生境过滤来解释:群落系统发育结构表现出离散结构是由竞争排斥作用起主导因素,而聚集结构则是由生境过滤作用所主导。本文研究结果表明:西藏色季拉山森林中,乔木群落的系统发育结构在高海拔和低海拔地区表现出聚集状态,中间海拔地区呈现离散结构,生态位过程中的生境过滤和竞争排斥对色季拉山森林群落物种多样性的维持起到了重要作用。低海拔地区主要存在强烈的人为干预因素,这种环境过滤促使色季拉山群落系统发育呈现出聚集的特点。在色季拉山中海拔地区,生境的异质性占据了主导因素,物种间的竞争变得愈发激烈,这些都是使群落呈现离散结构的重要因素。随着海拔的进一步升高,生长环境越来越恶劣,环境过滤作用的优势就凸显出来,群落系统发育则呈现出聚集状态。本研究中,色季拉山森林群落在高海拔地区呈现出聚集的发育结构,充分印证了生态位保守假说。与乔木层不同,色季拉山群落中灌木层和草本层在中低海拔地区系统发育呈现出离散状态,主要原因在于物种之间的竞争关系,地下部分主要是对水分和养分的需求,地上部分主要是对光生态位的竞争需求,所以选择更为合理的物种库验证群落系统发育随着海拔梯度变化格局就显得至关重要。乔灌草的群落系统发育结构受到环境变量和空间变量的影响较为严重,所以森林群落构建会受到环境过滤作用和空间变量的制约。色季拉山沿海拔梯度的群落系统发育β多样性与海拔距离差具有密切联系,相同海拔内乔木层、灌木层以及草本层的转变比预期要小,说明在相同海拔内环境的异质性相对也较小。随海拔距离差增大,随机过程呈现出主要作用,这主要与种子在较小的海拔跨度范围内不会发生散布限制等因素有关,随机的扩散过程对植物群落系统发育β多样性的形成就显得尤为关键。同时随着海拔距离差的增大,环境的异质性也增加,色季拉山森林群落中的系统发育转换要显著高于预期猜想,出现这一特点的主要原因是环境变量和空间变量的急剧变化。
零模型和物种库的选择对群落系统发育生态学的研究有着深层意义[29]。在色季拉山不同尺度和地形生境设置零模型,可以比较群落的系统发育结构以及物种β多样性与随机群落的差异程度。物种库通常是指一个地区可进入某一特定群落的潜在物种数目[30],但是在具体实验操作过程中往往很难通过实验数据来证实实际的物种库特点,所以本研究选取色季拉山森林群落内所有物种作为小尺度物种库,这也是当今世界生态学研究方法中相对合理的一种选择方法。Swenson等[31]研究发现,系统发育聚集的可能性与物种库的空间尺度呈现正相关关系。然而,目前研究比较热门的零模型也存在一定的局限性,在进行群落生态学探究的时候有必要全面考虑群落中的物种丰富度、多度以及生态学特征。
4 结论
本研究揭示了色季拉山森林群落构建随海拔梯度的变化规律, 研究结果表明在青藏高原典型森林群落中生态位过程对群落的构建发挥着更为重要的作用。生境过滤是低海拔群落构建的主导作用因素, 同时人为干扰也是影响低海拔植物群落系统发育聚集的主要原因之一。到了高海拔地区,竞争排斥因素则加入进来,竞争排斥和生境过滤2种主要因素对森林群落构建同等重要。但高海拔群落的系统组配机制及其驱动因素目前研究尚不明朗,还需要进一步探究。一般情况下,采用群落系统发育方法来解释和探讨群落组配机制, 这种方法在生态学界也得到了一致认可,对全球生物多样性的保护有着重要意义,但考虑到本研究还存在不足之处, 如受客观条件的限制,在色季拉山高海拔地区选取的群落样方数量较少、缺乏群落的功能性状特征数据、受小生境的环境因子等因素限制……,接下来,需要综合开展多维度研究来进一步深入探讨色季拉山森林群落构建的机制和青藏高原地区物种多样性维持机制及其驱动因素,并结合植物功能性状进行综合分析,研究其生理机能。
