乳酸菌粘附肠道上皮细胞的研究进展
2020-12-01李慧臻焦文姝王成风霍贵成李柏良
占 萌 李 春 李慧臻 李 娜 焦文姝 王成风 霍贵成 李柏良
(东北农业大学 乳品科学教育部重点实验室 哈尔滨150030)
在人类和其它哺乳动物的胃肠道中大约居住着1014细菌,共1 000 多种,它们在生长和形成不同微生物菌群的过程中会不断与宿主相互作用。这些细菌产生大量的代谢物被宿主视为刺激因子。微生物菌群的行为显著影响宿主的健康。近年来,使用全基因组分析技术对细菌进行全基因组测序和微生物群的宏基因组分析,揭示了宿主与人体复杂生态系统内细菌共生关系的结构。
乳酸菌(lactic acid bacteria,LAB) 是人体肠道中的优势菌群,尤其在十二指肠到回肠末端的区域含量丰富。乳酸菌是可以发酵碳水化合物产生大量乳酸的细菌统称。研究表明LAB 具有促进营养成分的分解、吸收,增强机体免疫力,治疗腹泻[1],改善胃肠道功能和降低胆固醇及抗氧化[2-3]等作用。胃肠道中定殖的乳酸菌数量和种类与机体的健康功能密切相关。乳酸菌对胃肠道的粘附是其定植的关键先决条件之一,也是发挥益生功能的首要条件。
粘附是长期历史进化过程中产生的生态特性,其特点是特异性和生理性。产生这种特性的根本原因是“生命与环境的同一性”。粘附(adhesion)通常指微生物与宿主的上皮细胞接触后,通过生物化学作用使微生物与宿主细胞上皮表面相连接。乳酸菌对肠道上皮细胞的特异性粘附作用有助于其在肠道定植,增强乳酸菌与肠道细胞之间的信号交流,抑制病原菌在肠道的定植和提高机体的免疫力。乳酸菌在肠道中定植的前提是其必须粘附于宿主细胞表面[4-5]。目前普遍将乳酸菌粘附性作为评价其能否成为益生菌的首要标准[6-8]。
1 乳酸菌粘附的生物学效应
1.1 维持肠道菌群稳态
乳酸菌粘附于肠道黏膜有助于维持肠黏膜形态与功能的完整性,对保证肠道正常菌群稳态具有重要作用。乳酸菌主要通过空间位阻效应来维护肠道菌群的正常结构[9]。病原菌使宿主致病的第1 步是对肠道上皮细胞的粘附。当高黏附性的LAB 先于病原菌定植在肠道时,占据了致病菌的粘附位点,阻断其与肠黏膜的接触,在肠道表面形成一层物理屏障,从而抑制定植和转位,维护肠黏膜的完整性和肠道内稳态环境[10]。Singh 等[11]研究表明受试的8 株活罗伊氏乳杆菌(L.reuteri)均能抑制大肠杆菌ATCC25922 (Escherichia coli ATCC25922)、伤寒杆菌NCDC113(Salmonella typhi NCDC113)、单核细胞增多性李斯特氏菌ATCC53135(Listeria monocytogenes ATCC53135)和粪肠球菌NCDC115 (Enterococcus faecalis NCDC115)对Caco-2 细胞的粘附作用。
1.2 调控黏蛋白表达
覆盖在胃肠道表面的黏液层是微生物菌群与宿主之间的第1 个接触点,它为微生物群提供了栖息地。黏液由肠道表面内的杯状细胞(goblet cell)分泌,黏蛋白(mucins)是其主要组分[12]。目前比较完整的乳酸菌遗传学工具和分析技术的进步显著加快了对乳酸菌菌株识别和粘附黏蛋白的分子机制研究。这些机制涉及细菌表面相关的多种粘附素与黏蛋白链,即碳水化合物-蛋白质的相互作用。Hymes 等[14]研究发现,相对于亲本菌株,敲除纤连结合蛋白基因(fbpB)的突变菌株嗜酸乳杆菌NCFM (L.acidophilus NCFM)(ΔfbpB mutant)在体外对黏蛋白的黏附性发生显著降低。ΔfbpB突变体对人血浆纤连蛋白的黏附性降低了72%,对猪胃III 型黏蛋白的黏附性降低了47%。表明NCFM 菌体表面的纤连结合蛋白具有与黏蛋白结合的能力。LAB 与黏蛋白的相互作用,不仅有利于自身的长期稳定定植,而且能够调控黏蛋白的表达与分泌,增加黏液层厚度,增强黏膜免疫力,进而稳固黏膜屏障。李晶[15]通过体外黏蛋白模型从哺乳仔兔小肠黏膜中分离到的菌株中筛选出高粘附菌株植物乳杆菌2(L.plantarum 2),并且发现单独灌服植物乳杆菌2 仔兔小肠中有益菌比例增加,回肠绒毛生长旺盛和空肠、回肠黏蛋白MUC1,MUC2 的mRNA 表达水平提高,黏膜屏障功能增强。
1.3 增强细胞间的信号交流
当LAB 的粘附素与宿主细胞表面的特异性受体结合时,就会启动一系列信号传递过程。Konstantinov 等[16]研究了嗜酸乳杆菌NCFM 及其细胞表面化合物与树突状细胞 (dendritic cells,DCs)之间的相互作用,发现NCFM 通过与树突状细胞表面的特异性细胞间粘附分子C 型凝结素(DC-specific ICAM-3-grabbing nonintegrin,DCSIGN)结合,诱导IL-10 和IL-12p7 浓度依赖性产生。益生乳酸菌代谢物也能通过调节细胞信号途径来保护肠道屏障。由鼠李糖乳杆菌GG(L.rhamnosus GG,LGG)分泌的p40 和p70 蛋白可阻止细胞素引起的细胞调亡,进而增强肠屏障功能,主要有两种途径可能通过磷酸肌醇-3’-激酶(PI3K)的依赖性途径激活抗细胞调亡蛋白激酶B(Akt/PKB)或通过抑制TNF-α,IFN-γ 或IL-1β 对促凋亡p38 丝裂原活化蛋白激酶 (p38/MAPK)活化[17-18]。
1.4 粘附的特异性
乳酸菌的粘附具有菌株特异性和宿主特异性。不同菌种对同种宿主的粘附能力也存在差异,即使菌种相同,由于在菌株水平上存在差异,其黏附性也不尽相同。Tuo 等[8]检测了15 株植物乳杆菌、5 株干酪乳杆菌和2 株鼠李糖乳杆菌对Caco-2 细胞的粘附能力,发现同种乳酸菌的不同菌株的粘附能力差异较大。Argyri 等[19]从食品中分离出37 株戊糖乳杆菌,同样发现不同菌株的粘附能力差异较大,粘附能力最高达60.78%,最低仅33.72%。同样,对于同一乳酸菌菌株,粘附不同的宿主,最后的结果也可能不同,表现出明显的宿主特异性。分离自同一群体的乳酸菌对本群体肠道的粘附能力较本群外的动物更强,甚至不在本群外的动物肠道中粘附[20]。
2 粘附机制
20世纪90年代初,1 株乳杆菌菌株用蛋白酶处理后,对HT29-MTX 细胞分泌的黏液的黏附量减少[21]。第1 次研究表明了乳杆菌中存在与病原菌相似的粘附过程,即:菌体细胞表面的某个成分与肠上皮细胞受体的相互作用。近些年来,关于乳酸菌粘附特性的研究十分广泛,包括从分子水平和基因水平探索乳酸菌与肠道上皮细胞或者胞外基质的粘附机制。目前普遍认为LAB 的粘附是乳酸菌表面特异性识别分子和宿主靶细胞表面的相应受体,在空间结构上相互匹配并结合,也就是黏附素-受体学说。
2.1 黏附素
黏附素分为蛋白类黏附素和非蛋白类黏附素。蛋白类黏附素又包括表层蛋白 (S-层蛋白,SLP)和转肽酶依赖蛋白(包括黏液结合蛋白和菌毛蛋白)[21-22]。这些蛋白黏附素也是目前研究较深入和广泛的黏附素。表1中列出目前报道的乳酸菌的黏附素。
2.1.1 表层蛋白 乳酸菌表层蛋白是目前研究最多,也是乳酸菌中比较普遍的一类黏附素。S-层蛋白几乎存在于所有的革兰氏阳性菌及革兰氏阴性菌中,由单分子晶体排列的单一蛋白质亚单位或糖蛋白以对称的排列形式构成,通过非共价键连接于多数细菌的细胞壁[23]。乳杆菌的表层蛋白是已知的表层蛋白中相对分子质量最小的一类,介于25~71 ku 之间,占细胞蛋白总量的10%~15%,具有高度稳定的三级结构[15]。与其它菌种相比,乳杆菌的SLP 等电点范围在9.35~10.40 之间。多数菌种SLP 的等电点较低,而乳杆菌SLP 等电点偏高成为其独有的特征之一[24]。目前已从多种乳酸菌中分离鉴定出表层蛋白。如嗜酸乳杆菌、瑞士乳杆菌(L.helveticus)、短乳杆菌(L.brevis)、卷曲乳杆菌(L.crispatus)、开菲尔乳杆菌(L.kefir)、布氏乳杆菌 (L.buchneri)、类高加索酸奶乳杆菌(L.parakefir)和格氏乳杆菌(L.gasseri)等,大部分乳酸菌的SLP 为非糖基化结构,也有少数乳酸菌(如布氏乳杆菌和开菲尔乳杆菌)具有糖基化的SLP,并且大部分乳酸菌都有编码SLP 的基因[15]。其中,嗜酸乳杆菌ATCC 4356 和NCFM、卷曲乳杆菌JCM 5810 均含有两个SLP 编码基因,短乳杆菌ATCC 14869 和嗜酸乳杆菌KLDS1.0901 各含有3个编码SLP 的基因。具有亲缘性的菌株的SLP 编码基因序列具有相似性[25]。
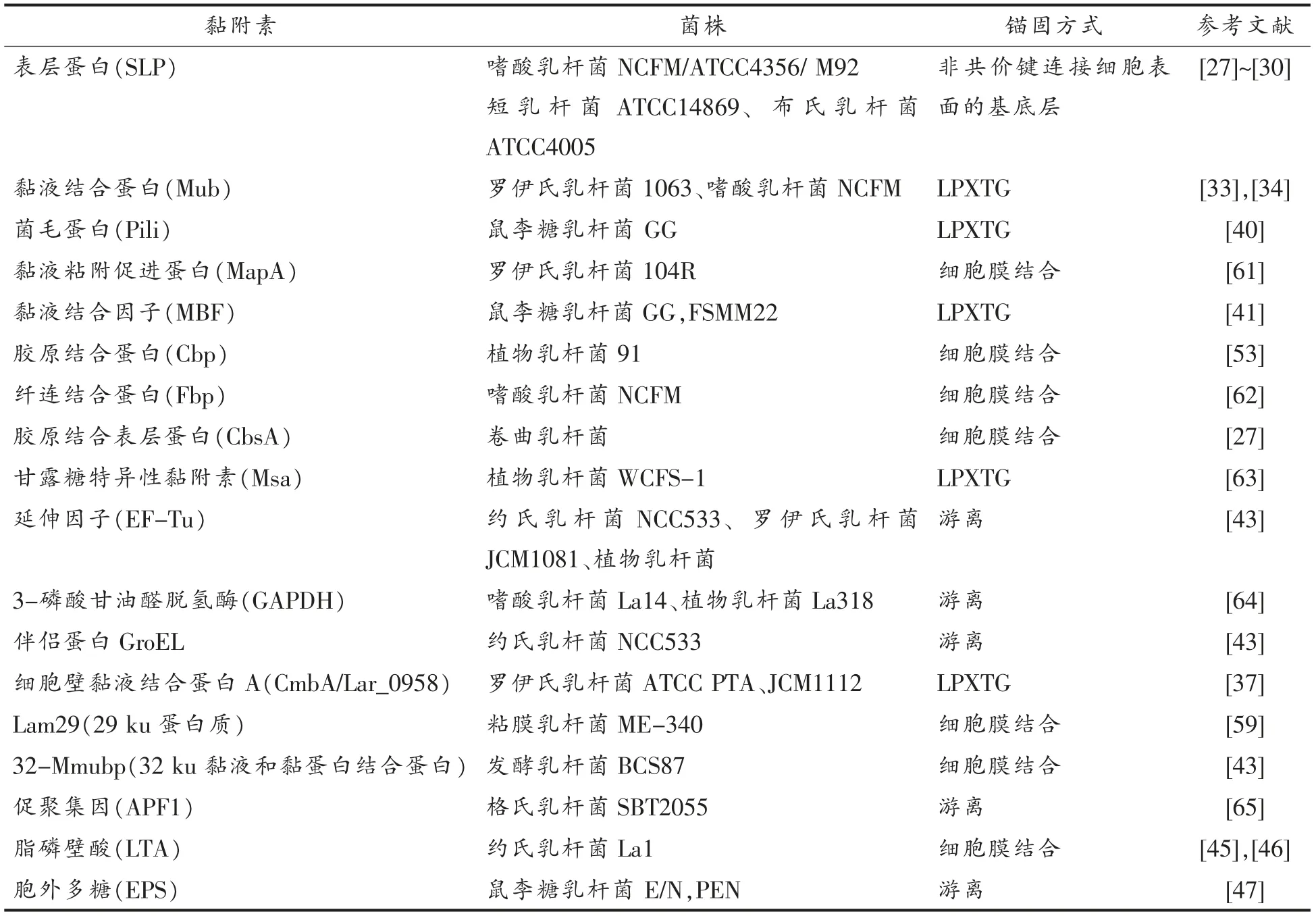
表1 乳酸菌相关的黏附素Table 1 Adhesion factors in Lactobacillus species
关于SLP 结合到细胞外基质(ECM)蛋白并粘附到肠上皮细胞的证据己从嗜酸乳杆菌NCFM 的SlpA 蛋白[26]、卷曲乳杆菌JCM5810 的CbsA 蛋白[27]、卷曲乳杆菌ZJ001 中的SlpB 蛋白[28]和短乳杆菌ATCC8187 的SlpA[29]蛋白中得到证实。对于SLP 具体哪一段氨基酸序列参与ECM 组分的粘附,有研究表明卷曲乳杆菌JCM5810 CbsA 蛋白N 端位置31~274 处的氨基酸残基是与鸡结肠胶原蛋白结合所必需的结构域[30]。短乳杆菌ATCC 8287 的S-层蛋白SlpA 介导了菌体细胞与人结肠癌上皮细胞系和纤连蛋白的结合,并且SlpA N 末端的81 个氨基酸足以与上皮细胞结合[31]。
近年来陆续有研究发现,表层蛋白对乳酸菌发挥益生功能有着卓著的贡献。SLP 除了作为黏附因子介导乳酸菌粘附定殖于肠道上细胞或ECM 蛋白以延长其在肠道中发挥益生作用的时间之外,还有助于乳杆菌抑制病原菌对肠道组织的感染,减少致病菌对肠道的细胞伤害,以及调节免疫的功能[26]。有研究报道嗜酸乳杆菌ATCC 4356 的SLP 可以结合特异性抗原DC-SIGN,调节免疫反应,抑制胡宁病毒引起的细胞损伤[32]。
2.1.2 黏液结合蛋白 在目前发现的乳酸菌黏蛋白粘附因子中,黏液结合蛋白家族可能是研究最广泛的。最初在罗伊氏乳杆菌1063 和嗜酸乳杆菌NCFM 中发现,MUB 蛋白是一种大分子蛋白质(分子质量≥300 ku),其N-端具有一个特征性YSIRK 分泌信号序列,以及C-末端存在一个转肽酶依赖性LPXTG 锚定基序[33-34]。MUB 蛋白包含两个特征保守的重复区域 (如图2所示),分别为Mub1 和Mub2,每个重复序列由183~206 个氨基酸残基组成。Mackenzie 等[35]通过使用MUB 蛋白抗体的抑制试验证实MUB 蛋白在罗伊氏乳杆菌ATCC 53068 中作为黏蛋白粘附因子的作用。研究报道发现MUB 的重复序列与单核增生李斯特菌报道的黏液蛋白结合蛋白 (MucBP,PF06458)域和氨基酸序列高度相似[36],这促使对其它菌种中MucBP 同源蛋白的进一步研究。目前为止已在很多乳酸菌中发现和MucBP 结构域相似的氨基酸序列的表面蛋白,然而很少对它们的真正作用做深入研究。最近从罗伊氏乳杆菌中提取纯化的MUB 蛋白的组织学分析表明,MUB 蛋白能识别黏蛋白链末端的唾液酸残基,因此MUB 蛋白粘附于黏蛋白的具体作用方式正在逐渐被发现[37]。此外,基于MucBP 相关结构域 (MUBAD) 化学合成的MUB70,不同于罗伊氏乳杆菌mub 编码的MucBP结构域,其特征是能与Muc2 型黏蛋白的硫酸碳水化合物结合。而在结肠黏液癌发病过程中,Muc2 型黏蛋白分泌过多,因此基于MUB70的特异性结合亲和力可以开发出黏膜癌的潜在标记物。随着生物信息分析技术的发展,通过菌体基因组分析序列中存在的YSIRK 信号、LPXTG 序列或MucBP 结构域推定假定的黏附因子。比如,鼠李糖乳杆菌中发现的黏膜结合因子(MBF)能与层黏连蛋白、纤连蛋白、IV 型胶原等黏蛋白和细胞外基质(ECM)蛋白结合[38]。
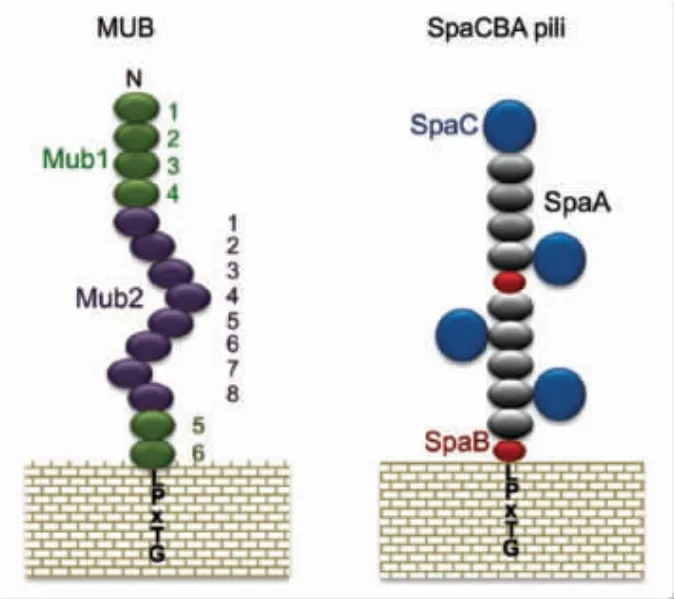
图1 MUB 蛋白和SpaCBA 菌毛蛋白结构示意图[43]Fig.1 Schematic model of MUB and SpaCBA pili[43]
2.1.3 菌毛蛋白 菌毛(fimbriae)长期以来被认为是致病菌特有的特征。然而,对约氏乳杆菌NCC533 进行全基因组测序后,在乳杆菌属物种中首次发现编码菌毛的基因簇[39]。随后,通过免疫电镜在鼠李糖乳杆菌GG 细胞表面鉴定出菌毛。鼠李糖乳杆菌菌毛是由3 个亚基SpaA,SpaB 和SpaC(称为SpaCBA)聚合形成[40](如图2所示)。其中SpaC 分布于菌毛整个长度,具有凝集素样特征,能够特异性结合位于黏蛋白非还原末端的半乳糖基[41]。除了SpaCBA 操纵子外,在鼠李糖乳杆菌GG 基因组中还发现第2 类菌毛基因 (称为SpaFED)[40]。研究发现乳酸乳球菌(Lactococcus lactis) 中也存在SpaFED 菌毛蛋白,位于顶端的SpaF 菌毛蛋白是介导菌体细胞与黏蛋白、胶原蛋白和纤连蛋白以及Caco-2 和HT-29 肠细胞系相互作用的重要决定因素[42]。
2.1.4 其它黏附素 乳酸菌粘附到胃肠道的过程中除这些常见的蛋白因子参与外,还有一些兼职蛋白,除了行使主要功能外,也参与乳酸菌的粘附作用。如EF-Tu,GAPDH,伴侣蛋白GroEL,ATP 结合体(ABC)转运子等[43]。这些蛋白不含有典型的保守细胞表面锚定基序,其在细胞表面的定位受环境因素的影响,这些蛋白质可作为次要的黏附因子发挥作用,然而它们与碳水化合物链表现出特定的相互作用。最近研究[44]发现乳杆菌表面的丙酮酸激酶(PK),葡萄糖-6-磷酸异构酶(PGI)、磷酸甘油酸激酶(PGK)、伴侣蛋白GroEL 和EF-Tu能与宿主表面的肌动蛋白粘附。此外,PGK-肌动蛋白结合域分析显示,PGK 中127 号氨基酸位置的赖氨酸可能在介导细菌粘附肌动蛋白的过程中起关键作用。
一些非蛋白因子在乳酸菌粘附宿主的过程中也发挥着重要作用。早在1985年,Camp 等[44]在研究双歧杆菌的粘附时发现,LTA 通过增加细胞壁的电负性及疏水性影响乳杆菌粘附肠上皮细胞的能力。目前已发现乳酸杆菌细胞表面的EPS、脂多糖(LPS)以及肽聚糖(WPG)等成分在乳酸菌的粘附过程中也起到一定的作用[45-46]。研究发现,嗜酸乳杆菌中不同类型的EPS 影响细菌粘附到ECM组分的过程[47]。在嗜酸乳杆菌BG2F04 中,细菌表面的碳水化合物部分介导了菌株粘附到Caco-2细胞及由HT-29-MTX 分泌的黏液层的过程[48]。
2.2 黏附受体
黏附素受体是乳酸菌与宿主细胞粘附的关键成分之一。乳酸菌表面含有一些黏附素,只有宿主细胞有针对这些黏附素的特异性受体才能完成粘附过程,进而传递胞外信号,启动一系列细胞生物化学反应。菌株的黏附素受体种类显著影响粘附方式,也可以认为黏附过程具有菌株特异性。值得注意的是黏附受体的成分会随着生物体的发育分化和营养物质的变化而改变,相应的配体也会随之变化,这就解释了为什么幼小畜禽和婴儿在不同的生命阶段肠道菌群有演替变化的过程。例如:婴儿在刚出生时肠道菌群以大肠杆菌和链球菌为主。在母乳喂养期,肠道中双歧杆菌数量急剧增加,同时大肠杆菌和链球菌减少。断奶后,链球菌和大肠杆菌的数量进一步减少,出生后的第2年会形成成人型的肠道菌群[49]。
目前鲜有研究报道乳酸菌的黏附素受体。张博[47]将HT-29 细胞用高碘酸钠处理后,与乳酸乳球菌NZ900 共孵育,做黏附试验,结果表明:NZ900 对处理的细胞黏附量与处理前无差异,推测糖类不是菌体表面CwaA 蛋白的主要黏附受体。通过糖抑制试验证实CwaA 蛋白的黏附受体中不含有C-C 单键,即不是糖或糖蛋白。再通过蛋白酶处理试验表明CwaA 的黏附受体可能是蛋白类物质,因此随蛋白酶的处理浓度的提高,对HT-29 细胞的粘附能力逐渐下降。Fujiwara 等[50]研究发现,长双歧杆菌(Bifidobacterium longum)培养液中存在的一些分泌蛋白可以抑制产肠毒素型大肠杆菌(ETEC)的特异性受体GA1 的粘附,抑制效果随着加入分泌蛋白浓度的增加而增强,表明肠道上皮细胞表面的GA1 位点也有可能是长双歧杆菌黏附素的受体。长双歧杆菌通过竞争性地与GA1 位点结合,有效排斥病原菌与肠上皮细胞的粘附。对罗伊氏乳杆菌的研究表明,该菌株能够特异性地与肠上皮细胞外侧的糖脂和硫苷脂结合,这两种成分可能是罗伊氏乳杆菌黏附素受体[51]。目前对乳酸菌黏附素受体的报道很少,大部分研究多是推测黏附素受体可能的类型与性质,很少在分子水平对黏附素受体进行鉴定,以及分析其与黏附素的作用机制。关于乳酸菌黏附素特异性受体的鉴定及相互作用需更深入研究和发掘。
3 影响乳酸菌粘附的因素
乳酸菌的粘附效应不仅与黏附素和黏附受体的特征相关,也与外界的环境,如温度、酸碱度、离子强度等因素有关,以及菌体生长阶段、菌液浓度、菌体表面理化性等自身因素也影响乳酸菌的黏附性。
有研究报道,温度对植物乳杆菌的黏附性是破坏性的,因为温度可能会改变乳酸菌表面黏附素的结构,从而影响菌体与肠道上皮细胞表面受体的特异性结合,使乳酸菌的黏附能力下降。也有研究表明,温度对双歧杆菌的黏附性没有显著影响,即使双歧杆菌在较高的温度下预处理一段时间,其粘附能力与活菌非常相似[52]。可能温度对黏附性的影响存在菌株差异性。Yadav 等[53]研究了pH 值对植物乳杆菌Lp5276,Lp91 与胶原蛋白相互作用的影响,结果表明在pH 6 时,两株植物乳杆菌对胶原蛋白的粘附性较高。
pH 值对乳酸菌粘附能力的影响具有菌株差异性。总体而言,在偏酸性或中性条件下有利于乳酸菌的粘附。离子浓度也显著影响乳酸菌的黏附性,尤其是Ca2+。王斌等[54]研究了Ca2+浓度对植物乳杆菌L2 粘附Caco-2 和IEC-6 细胞能力的影响,结果表明,L2 对两种细胞的黏附量分别达到595 CFU/100 细胞和538 CFU/100 细胞。当培养液中增加浓度为0.9 mol/L Ca2+时,L2 菌粘附IEC-6 细胞的数量高达11 700 CFU/100 细胞,表明Ca2+能显著增强L2 菌的粘附能力。Ca2+促进粘附效果可能是因为激活了Ca2+介导的信号通路,从而促进细菌与细胞表面受体间的特异性结合。
乳酸菌在不同的生长阶段,其形态结构、菌体活性、代谢产物等都有一定的差异。大量研究表明,大多数乳酸菌生长到稳定期初期的黏附性大于其它时期。可能在稳定期初期,菌体细胞形态趋于完整,且表达较高的黏附素。一般情况下,随着菌悬液浓度的增大,乳酸菌黏附能力也随之增大,当达到一定浓度时,乳酸菌黏附量趋于稳定。研究报道[55]当植物乳杆菌菌体浓度在2×108CFU/mL 以上时,其对Caco-2 细胞黏附性没有显著差异,说明菌体达到饱和占位状态。乳酸菌的表面疏水性也对其黏附能力产生一定影响。最近几年将其作为评价乳酸菌黏附性的重要指标。乳酸菌表面疏水性是由其表面疏水性的蛋白质、糖类及其它化合物所赋与的一种表面特性。Ehrmann 等[56]研究表明,乳杆菌的疏水性与其黏附能力高度相关,疏水性越高的细菌其黏附力越强,反之,细菌的黏附力较低。
4 乳酸菌黏附的评价方法
目前已建立多种检测方法来评价乳酸菌的黏附性能,主要分为直接测定法和间接测定法。直接测定法一般采用体外细胞模型模拟人体肠道上皮细胞,通常用Caco-2、HT-29 和Hela 细胞等模型与乳酸菌共同孵育一段时间后,通过粘附计数来评价乳酸菌的粘附效果。通过革兰氏染色后显微镜观察和平板计数的方法检测黏附量。这些方法存在一定缺陷,主要是与体内试验结果存在差异,忽略了人体肠道上皮细胞表面覆盖着一层厚厚的黏液层,通常情况下乳酸菌无法通过黏液层直接接触上皮细胞。乳酸菌若要与上皮细胞发生作用引起特异应答,首先要粘附于黏液层。也有学者直接采用黏蛋白模型来评价乳酸菌的粘附能力。Buntin 等[57]利用体外黏蛋白模型评价了18 株植物乳杆菌的粘附特性。
还有研究报道用荧光染料标记乳酸菌,对菌体进行追踪指示,进而观察乳酸菌对肠黏膜的粘附能力。细胞增殖示踪荧光探针(cFDA-SE)和异硫氯酸标记(FITC)是两种比较常用的标记染料。其中cFDA-SE 对乳酸菌进行标记具有安全性高、方法很简单、省时、标记率高、成本低等优点,对最终收集到的乳酸菌用流式细胞仪等仪器检测标记细胞的数量,便可计算乳酸菌的黏附率。谢琼[55]用cFDA-SE 标记植物乳杆菌ZDY2013 后,灌胃给小鼠,探究植物乳杆菌ZDY2013 在体内的粘附情况是否与体外试验一致。流式细胞仪检测标记的乳酸菌在小鼠肠道中的定植结果表明,ZDY2013能很好地附在小鼠的不同肠段,与体外试验结果有较好的一致性。目前也有一些研究者基于酶联免疫(ELISA)的方法检测乳酸菌的粘附性能[58]。虽然ELISA 的方法可以同时处理多个样本,准确性高,但是需要特殊的抗体,试验成本较高,尤其在处理少量样品时,不适合用此方法。
间接评价法是指通过测定乳酸菌细胞表面的理化性质来间接反映其粘附能力的强弱。比如疏水性、自聚集能力和表面电荷量等,其中最常用的表面性质是疏水性。任大勇[12]检测了9 株乳酸菌的自凝聚能力、疏水性和对Caco-2 细胞的粘附能力,通过相关性分析表明,乳酸菌的疏水性与菌株的粘附能力存在一定的正相关性。研究乳酸菌表面疏水性与其黏附性的相关性,一方面可以方便、快速地筛选高黏附性菌株,另一方面,有助于弄清乳酸菌与宿主粘附的作用机制。然而,也有研究不认为两者之间具有相关性[59]。疏水性只能作为筛选高黏附性乳酸菌的一个参考指标。
近些年来,计算机在科研中得到广泛的发展和应用。一些新的评价细菌粘附能力的方法出现。如计算机分子模拟法,即通过计算机软件以原子水平的模型来模拟研究对象的结构和行为,从而进一步模拟物质的各种理化性质。Das 等[60]通过蛋白质-蛋白质对接软件(Hex ver 6.0),分析4 种乳杆菌细胞表面的黏蛋白与肠黏蛋白的三维空间结构对接情况,筛选出对肠黏蛋白具有最高粘附能力菌株。
5 展望
目前,众多研究报道了乳酸菌的粘附机理,主要集中在乳酸菌黏附素上,对肠道上皮细胞和黏液层受体的研究还应深入探索。表层蛋白是目前研究较全面和深入的黏附素。近些年来对乳酸菌进行全基因组测序和基因组学分析对深入了解乳酸菌粘附机制具有深远意义。在国家生物技术信息中心(NCBI)网站上可查多株乳酸菌表层蛋白的基因序列,可以通过基因组比对分析,得到全基因组测序菌株是否具有分泌SLP 的能力。对于具有益生功能但粘附能力很差的乳酸菌,将已知高黏附性乳酸菌的粘附相关蛋白基因序列重组到这些黏附性差的益生菌基因组中,使其在宿主体内粘附定殖并更好地发挥益生功能,这是一个非常值得深入研究的新思路。
