新疆叶尔羌河流域斑重唇鱼寄生单式指环虫中国一新纪录
2020-11-26容梦婕喀迪尔丁艾尔肯张文润郝翠兰穆妮热喀迪尔田胜利
容梦婕,喀迪尔丁·艾尔肯,张文润,郝翠兰,穆妮热·喀迪尔,田胜利,岳 城
(新疆农业大学动物医学学院,乌鲁木齐 830052)
叶尔羌河位于新疆西南部,发源于喀喇昆仑山脉北麓,是我国最大的内陆河—塔里木河的重要源流之一。该河的发育形成、地理地貌特点以及上游水系的特殊自然环境,使得该河孕育有一些特殊的土著鱼类资源,包括裂腹鱼亚科(Schizothoracinae)及鳅科高原鳅属(Triplophysa)[1]鱼类。斑重唇鱼(Diptychusmaculates)隶属于鲤形目(Cypriniformes)鲤科(Cyprinidae)裂腹鱼亚科,在我国主要分布于新疆天山以南的塔里木河水系和天山以北的伊犁河流域各水系[2-3]。近年来,由于人为因素的干扰,斑重唇鱼的分布区域正在逐渐缩小,种群数量逐渐下降,现已被列为新疆维吾尔自治区II级水生野生保护动物[4]。为了保护斑重唇鱼类资源,牛建功等[5]研究了斑重唇鱼在人工条件下驯化养殖、繁育和增殖放流等技术,以达到物种保护、生态修复和恢复种群数量的效果。此外,相关学者对该流域斑重唇鱼的研究多集中于野外资源调查和种群进化等研究[6-9],对危害其种群健康的寄生虫病原报道较少。
指环虫(Dactylogyrus)是单殖吸虫种类中最为丰富的一个类群,其宿主种类多达28科194属,主要寄生于鲤科鱼类的鳃部[10],表现出极强的宿主特异性。目前,由于仍有许多鲤科鱼类尚未进行全面调查,有学者认为还存在大量的指环虫新种有待发现、命名和描述[11]。据报道,寄生于裂腹鱼亚科鱼类的指环虫全球约有16种[12-13],在我国被记录的有8种指环虫,其中寄生于斑重唇的指环虫仅有一种,为喀迪尔丁等[14]在新疆伊犁河流域采集的重唇鱼指环虫(Dactylogyrusdiptychus)。本课题组于2019年5月-2020年7月进行叶尔羌河鱼类寄生虫区系调查时,在斑重唇鱼鳃部采获1种指环虫,与伊犁河流域发现的重唇鱼指环虫存在较大形态差异。经资料检索该种在我国未见报道,本研究通过传统形态学和分子生物学方法鉴定,明确该指环虫种类,丰富叶尔羌河鱼类寄生虫病原体的种类组成,补充我国单殖吸虫名录,同时补充该指环虫的分子数据,为该虫在分子系统进化方面积累资料。
1 材料与方法
1.1 样品的来源与处理
2019年5月-2020年7月在叶尔羌河流域的塔什库尔干河段(37°46′ N,75°13′ E)共采集斑重唇鱼272尾。对新鲜的鱼类标本进行拍照、编号、称重和测量,并记录相关数据。鱼类解剖及寄生虫采集主要参照《鱼病调查手册》[15]。将宿主鳃片逐一取下后,滴加少量生理盐水置载玻片上,用解剖针轻轻刮取粘液,并在解剖镜下逐一检查,挑取虫体。发现虫体后将其吸取到盛有清水的小皿中,冲洗干净,置于1.5 mL的EP管中,滴加75%和95%浓度的酒精固定。
1.2 形态学鉴定
在解剖镜下挑取形态完整的12只虫体逐一放置在载玻片上,用4% PVL(聚乙烯醇-乳酸酚染液)固定封片。在光学显微镜(Nikon E2000)下观察,通过EZ-MET软件测量后吸器及交接器中的几丁质结构。利用t检验比较测量结果平均数的差异是否显著。测量拍照后在硫酸纸上绘制特征图,并依据文献[12,13]进行形态学鉴定。所制虫体标本保存在新疆农业大学动物医学学院寄生虫教研室。
1.3 DNA提取、扩增及测序
取95%乙醇固定的虫体样本置于TE Buffer(pH 8.0)中浸泡过夜后,将虫体分别装入1.5 mL的EP管中,用Trans Gen动物组织基因组DNA提取试剂盒提取虫体DNA(具体操作方法依照试剂盒说明书)。
选取18S-ITS1-5.8S rDNA序列作为目的序列,扩增引物分别参照imková等[16],上游引物S1:5′-ATTCCGATAACGAACGAGACT-3′,下游引物IR8:5′-GCTAGCTGCGTTCTTCATCGA-3′,由生工生物工程(上海)股份有限公司合成。PCR反应体系总体积为25 μL:2 × Easy Taq PCR SuperMix 12.5 μL,1 pmol/μL上下游引物各1 μL,10 ng/μL的模板2 μL,ddH2O 8.5 μL。同时,以双蒸水代替模板DNA作为阴性对照。优化选取的反应条件:95 ℃预变性4 min,92 ℃变性1 min,55 ℃退火1 min,72 ℃延伸90 s,35个循环,最后在72 ℃延伸10 min。
取5 μL PCR产物在1%的琼脂糖凝胶电泳检测,其余20 μL PCR产物跑胶后,切取目的条带,并用Trans Gen Biotech(全氏金)胶回收试剂盒进行纯化。纯化产物连接到pMD18-T载体上,再转入大肠杆菌(E.coli) DH5α感受态细胞中,LB平板培养,经PCR检测后每个样品选取2个阳性克隆送至上海生工进行测序。
1.4 序列比对及系统发育树构建
测序结果用 SeqMan 软件进行校对和编辑,在DNAMAN 7.0软件中进行序列比对,计算序列间的差异百分比。基于Kimura双参数模型,采用MEGA 7.0和BLAST计算所测序列之间的A、T、G、C的平均含量、碱基变异位点、简约信息位点等信息,并计算遗传距离。从GenBank数据库中选取与所得序列相似度较高的20种指环虫18S-ITS1-5.8S rDNA序列,以几丁嗜丽鱼虫(Cichlidogyrussclerosus)为外类群,一起导入 Phylosuite[17]软件中,进行贝叶斯法(Bayesian Inference,BI)建树和最大似然法(Maximum Likelihood,ML)建树。运用Model Finder进行模型计算,以AIC为标准筛选出ML树和BI树的最佳替换模型分别为TIM3+F+I+G4和GTR+F+I+G4。用 PhyloSuite 中自带的IQ tree和Mr Bayes软件建立ML和BI系统发育树。其中BI树共运算6 000 000代蒙特卡罗模拟(Markov chain Monte Crlo;MCMC),每100代取样1次,共进行两次重复,用后验概率来表示各节点的置信度。利用IQ tree 2版本[18]中的近似无偏检验(AU检验)ML和BI方法构建的系统发育树的拓扑构形差异是否具有统计显著性。
2 结果
对272尾斑重唇鱼进行剖检,共收集虫体2 995只。通过种类鉴定,结果发现斑重唇鱼仅感染了一种指环虫,总感染率为70.2%,平均感染强度为15.68 ± 1.88,最高感染强度为189。
2.1 虫体的形态学特征
基于12个封片标本,虫体呈柳叶状,为小型单殖吸虫,体长394(308 ~ 471) μm,体最大宽89(79 ~ 103) μm。眼点两对,咽近乎呈圆形,大小48(39 ~ 58) μm× 49(40 ~ 58) μm。后吸器位于体后端,类椭圆形,长69(64 ~ 76) μm,宽103(94 ~ 126) μm,包含1对中央大钩、1个联结片、7对边缘小钩。边缘小钩发育良好,可明显区分钩尖、钩柄和柄轴3部分,其中柄轴短而粗壮。7对几乎等大,全长23(19 ~ 24) μm,钩尖长5(4 ~ 6) μm,钩柄长8(7 ~ 9) μm,柄轴长9(7 ~ 10) μm。中央大钩基部较短,有长而弯曲的钩尖,全长61(56 ~ 68) μm,基部长40(37 ~ 45) μm,钩尖长25(22 ~ 28) μm,内外突明显分叶,且内突远远大于外突,长26(23 ~ 28) μm,外突长2(1 ~ 3) μm。联结片呈粗壮“一”字型,两端向上微翘且圆钝,中央略“凸”,大小4(3 ~ 5) μm× 43(40 ~ 48) μm。
交接器由交接管和支持器组成,交接管呈现出明显的中空管状结构,基部膨大,并被形似“烟斗”状的基质包裹,逐渐延伸膨大,末端形成喇叭状,管长42(38 ~ 45) μm。支持器与交接管基部相连,其结构如同俄文字母“Г”,末端呈指状弯曲并附着一个弧形突起,支持器长38(37 ~ 42) μm。阴道和卵未见。

图1 单式指环虫整虫(A);后吸器(B、D);交接器(C、E)Fig.1 Total view of Dactylogyrus simplex (A);Haptor (B and D);Copulatory organ (C and E) A、B、C:显微结构图;D、E:手绘图,比例尺=10 μm
2.2 虫体的分子鉴定及系统发育分析
扩增获得的指环虫rDNA片段进行校正后,最终长度为951 bp,包含18S和5.8S的部分序列(长度分别为514 bp和27 bp)以及ITS1区域的完整序列(长度为410 bp)。其中保守位点499个,变异位点515个,简约信息位点438个,以及73个自裔位点。A、T、G、C碱基的平均含量为23.5%、27.8%、26.5%、22.2%,A+T的含量高于G+C的含量。选取2个虫体的18S-ITS1-5.8S rDNA 序列(两序列的一致性为100%),上传至GenBank数据库,登录号为MT476980-MT476981。
ML法和BI法构建的系统发育树经AU检验后发现无显著性差异(P>0.05)。因而,选取BI树(同时标注ML分析的分支支持率)作为代表,用来说明系统发育关系(图2)。系统发育树分为两个大的进化支,其中斑重唇鱼上的重唇鱼指环虫与隐藏指环虫(D.cryptomeres)、具鳞指环虫(D.squameus)和页形指环虫(D.lamellatus)亲缘关系较近,聚为稳定的独立分支。而单式指环虫与鲫寄生的美丽指环虫(D.formosus)、锚钩指环虫(D.anchoratus)及弧形指环虫组成的分枝互为姊妹枝,且位于另一分支上。
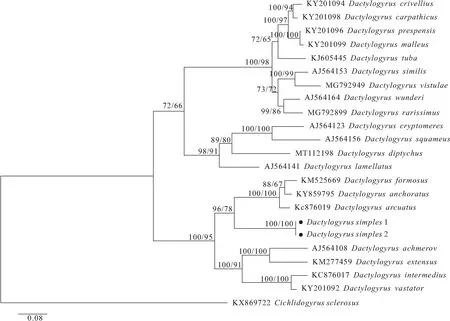
图2 基于 18S-ITS1-5.8S rDNA 序列构建的鲤科鱼类寄生指环虫贝叶斯法(BI)系统发育树Fig.2 Phylogenetic tree of Dactylogyrus based on 18S-ITS1-5.8S rDNA sequences based on the Bayesian Inferences (BI) method分支节点数值分别表示BI和ML的分支自展检验支持率
3 讨论
我国指环虫种类多样,约占世界的40%,这与指环虫强烈的宿主特异性密切相关[19]。通常情况下,指环虫可以根据宿主种类,进行初步的鉴定。经文献查阅,寄生于斑重唇鱼的指环虫有3种,分别为单式指环虫(D.simplex)、D.drjagini和重唇鱼指环虫(D.diptychus)[12,14],故将叶尔羌河斑重唇鱼鳃上检获的指环虫与这3种指环虫进行了形态及测量值的比较。
本种与D.drjagini和重唇鱼指环虫进行形态学特征比较时发现,不管是在交接器还是后吸器的形态上都明显存在不同(图3),主要体现在:本种中央大钩内外突明显分叶,且内突远远大于外突,钩尖较长,而重唇鱼指环虫中央大钩内外突分叶不明显,D.drjagini的虽明显分叶,但内外突几乎等大,且钩尖较短;本种仅有一联结片,而重唇鱼指环虫和D.drjagini含有背、腹两个联结片;交接器的差异也尤为明显。但本种与前苏联文献记载的单式指环虫[12]进行形态学特征比较后,发现两者形态非常相似,仅交接管基部的形态以及联结片中部有略微差别。两者交接管基部的“漏斗状”肌质的包裹状态不太一样,该形态的差异可能是由于虫体的观察角度不同[20]。本种的联结片中部略微“凸”出,而先前文献中单式指环虫的联结片呈“一”字,该结构的差异可能是因为发育和寄生虫对宿主或局部环境的适应而发生的变化[21]。此外,对特征形态的测量值对比发现(表1),两者的测量值差异不显著(P>0.05)。因此,从斑重唇鱼鳃部采集的指环虫为单式指环虫,该种在我国尚未报道,为我国一新纪录种。

图3 重唇鱼指环虫(A)、D.drjagini(B)、单式指环虫[前苏联(C)]、单式指环虫[本研究(D)]几丁质结构对比Fig.3 Comparison of the chitinous structure of Dactylogyrus diptychus (A),D.drjagini (B),D.simplex (C,former Soviet Union),D.simplex (D,the present study)a.中央大钩,b.边缘小钩,c.背联结片,d.腹联结片,e.交接器A、B、C图比例尺:20 μm;D图比例尺:10 μm

表1 新纪录种与斑重唇鱼寄生指环虫的形态特征比较Tab.1 Comparison of the morphological characteristics of the newly recorded species and the known Dactylogyrus of the D.maculates μm
传统形态学一直作为鉴定和研究指环虫的主要方法,但随着科技的发展,形态学的局限性逐渐被人们所意识到,因而有学者开始探索研究指环虫其它方法,以补充和完善传统形态学的不足及其局限性[22]。Cunningham等[23]是将分子生物学方法应用于单殖吸虫研究的首位学者。本研究基于18S-ITS1-5.8S rDNA序列构建的BI树显示,单式指环虫与鲫属鱼类寄生的美丽指环虫、锚钩指环虫及弧形指环虫聚为一支,其亲缘关系较近,而单式指环虫与同种宿主上寄生的重唇鱼指环虫分别位于不同的两支。该结果可能与地理隔离对宿主鱼类的影响相关[24-25]。
