动力性递增负荷运动中肌电与肌氧拐点及其联合评价无氧阈的方法比较
2020-11-26毕学翠郑晓鸿张英平
严 瀚,毕学翠,郑晓鸿,张英平,王 睿
无氧阈的研究一直是运动监控领域的研究热点之一,无氧阈的确定对于评价运动员有氧能力,制定适宜的有氧运动强度具有重要的意义。对于无氧阈的研究通常采用递增负荷实验,测试的指标主要包括气体代谢指标[1]、血乳酸指标[2]、心率指标[3]等。由这些指标而产生的通气阈、乳酸阈、心率阈来评价无氧阈。但是,在实践过程中也发现通气阈的测量需要佩戴面罩,影响受试者运动感觉,乳酸阈的采集有创,心率阈的稳定性较差,可检测率较低等缺点,研究者们一直试图寻找可靠、便携、无创的检测手段来评价无氧阈,其中表面肌电和肌氧技术成为新的热门监控指标。表面肌电技术作为一种无创,便携的可以量化肌肉活动的监控手段,很早就被尝试来成为评价无氧阈的新手段,肌电阈的概念由此产生,指用肌电设备收集运动过程中肌肉的放电信号,通过计算递增负荷中固定时间段肌电的变化,描绘出肌电的各项指标与负荷强度的一一对应曲线,观察当运动强度达到某一水平,该曲线会出现由平缓转向陡峭的“拐点”,此“拐点”就被称为肌电阈。主要反映肌肉由有氧代谢向无氧代谢转化的转折点[4]。肌电阈确定方法主要有2种[5],一种是以传统的以肌电图平缓变为陡峭的拐点为标准[6-7]。另一种是采用T.MATSUMOT[8]提出的强度—斜率曲线来求得。而肌氧技术主要通过近红外光谱术(near infrared spectroscopy,NIRS)[9]实时监测局部肌肉中的氧合血红蛋白(HbO2)浓度来反映局部肌肉的氧含量变化[10]。近年来在肌氧技术上的研究[11]也发现了局部肌肉肌氧变化过程也存在拐点现象,被称之为肌氧阈,并被证明可以用来反映局部肌肉的氧代谢能力[12],可以作为评价无氧阈的新指标。
肌氧阈和肌电阈已经被证实与乳酸阈、通气阈及最大摄氧量有显著相关性,而被提议用于无氧阈的确定以及有氧运动能力的预测。然而,也存在着很多争议[13]。首先在肌电阈值的确定上,杨继强[7]使用积分肌电(IEMG)来确定肌电阈,并发现与通气阈和乳酸阈存在高度相关。而T.V.CAMATA[14]研究认为,均方根振幅值(RMS)相对于IEMG能较好确定肌电阈。陈建宁[6]研究比较通气阈、乳酸阈和肌电阈,发现无氧阈出现时,IEMG阈的相关性最低。还有研究[15]发现肌电阈检出率并不高,不同肌肉之间的肌电阈值与无氧阈的关系存在差异。其次,在肌氧阈的确定上,争议主要出现在对于肌氧拐点的确定上,虽然多项研究都表明自己的方法确定的肌氧阈与无氧阈高度相关,但是由于受试者数量,受试者体脂,实验范式,评价标准等不同也使得肌氧阈的确定还存在着争议。所以说肌电肌氧这种无创便携的手段是否可以成为可靠的评价无氧阈的方法依旧不明朗。在无氧阈的确定上,血乳酸阈是应用最广也是最为长久经典的指标,本文以血乳酸阈为主要标准,检测主要的几种肌电、肌氧以及其联合评价无氧阈的手段(双线性拟合肌电拐点法[7]、双线性拟合肌氧拐点法[12]、肌氧15%差值法[16]、肌电与肌氧比值法[13]),试图寻找肌电、肌氧以及其联合评价无氧阈的可靠方法。
1 试验对象与方法
1.1 试验对象
本实验选取23 名首都体育学院体育教育专业本科生作为试验对象,体脂正常(体脂过高对受试者肌氧测量有影响[17]),无心、肺、肝、肾及内分泌和代谢等疾病史,下肢1年内未有重大伤病。受试者平均年龄(20.86±1.43)岁,平均身高(178.69±3.96)cm,平均体重(72.38±3.87)kg,平均体脂(18.2±3.72)%。试验前确定优势腿,确定方法为:运动员要求全力踢球,踢球腿为优势腿[18-19]。本研究中,所有受试者优势腿皆为右腿。
1.2 试验范式
采用功率自行车递增负荷试验范式,在英国产WATTBIKE功率自行车上做5 min 准备活动,期间帮助受试者调整座椅、把手到最合适位置。准备活动结束后休息3 min 开始试验,以80、110、140、170、200、230 w 每级递增30 w 做递增负荷试验直到力竭,每级骑行3 min。
1.3 表面肌电采集
肌电设备采用TRIGNO 肌电采集设备(DELSYS 公司生产制造,由1 个检测器和16 个传感器构成。仪器参数设定:采样频率为4 000 Hz EMG,148 Hz ACC/通道,加速度类型为内置三轴加速度)。参照以往文献以及肌电使用指南,根据自行车运动的主要发力肌肉位置,下肢肌电贴合部位分别是:左右股直肌,左右胫骨前肌,左右股二头肌,左右腓肠肌外侧头。传感器贴合前,对贴合部位进行去毛,酒精擦拭处理以降低电阻。肌电传感器贴合于肌腹位置,平行于肌纤维走向,利用双面胶布固定在皮肤,并用弹性绷带以适合松紧度包扎以防掉落。
1.4 NIRS肌氧数据采集
利用NIRS 技术采集肌肉内氧含量,主要的采集器材为Moxy 肌氧监测仪(厂商由美国Fortiori Design LLC 公司制造,规格尺寸为61 mm x 44 mm x 21 mm,波长630 、750 、850 nm,约重42 g),采集部位为左右股外侧肌[20],传感器贴合于肌腹位置,平行于肌纤维走向,利用双面胶固定在皮肤,由于肌氧主要为光学采集,最后需要利用深色弹性绷带以适合松紧度包扎以防掉落或者干扰数据。
1.5 血乳酸采集
血乳酸的采集使用德国产EKF台式血乳酸分析仪,试验前先在受试者耳垂处打预留孔,并用棉球包裹防止出血过多或者凝血,以保证试验过程中可以在不影响受试者蹬车的情况下直接采血。在每级负荷结束即刻以及最后结束后3、5、7、10 min采集耳血,检测血乳酸,用H.STEGMAN[4]的方法测定乳酸阈强度(见图1)。

图1 递增负荷试验流程图Figure1 Incremental Exercise Test Experimental Flow
1.6 拐点确定
1.6.1 双线性拟合肌电拐点 数据由肌电产商提供的专业软件进行处理,首先用巴特赫兹带通滤波器进行滤波(10~400 Hz),再对滤波后的结果进行绝对值处理,最后以30 s 为一单位,截取递增负荷运动时间内的IEMG 和RMS,分别将8块肌肉的IEMG 和RMS 与递增负荷强度建立曲线,利用计算机自动拟合功能[21]判断肌电拐点,当2 段线性拟合结果中出现的残余误差平方和最小时,相应的分界点作为肌电拐点。
1.6.2 双线性拟合肌氧拐点 Moxy 的数据分析设备已经对肌氧进行归一化处理,采集的主要数据为肌肉内氧合血红蛋白所占百分比。同样以每30 s 对肌氧数据进行平均,作为该时间段的肌氧值。参照文献[12]利用计算机对肌氧指标进行2 段线性拟合,当2段拟合结果中出现的残余误差平方和最小时,相应的分界点作为肌氧拐点。
1.6.3 肌氧下降15%拐点法 采集的主要数据以每30 s对肌氧数据进行平均,作为该时间段的肌氧值。以肌氧下降幅度超过上一等级数据15%定义为拐点[20]。
②指标组成不明确。表中的指标是综合指标,不清楚具体包括哪些项目及对应指标,易造成理解分歧。如挡水工程、泄洪工程中的排水孔、土(石)坝中的面(趾)板止水等,在项目划分表中是单独列项的,而细部结构指标又综合了 “排水工程”“止水工程”。从逻辑上理解,二者没有重叠。再如,设计上常提供厂房照明设施清单,而细部结构指标中又包括了“照明工程”,不知道厂房是否除外。
1.6.4 肌氧/肌电比值法 通过Moxy 分析软件数据先求得脱氧血红蛋白的变化数值,在以每30 s 对数据进行平均,作为该时间段的脱氧血红蛋白数值,将每个时间对应的脱氧血红蛋白数值与肌电IEMG、RMS 数值相除,得到肌氧/肌电的比值,将数据以时间为横坐标,以比值为纵坐标建立一个5 项式曲线拟合方程,比值随时间增长后出现峰值并下降,这个峰值点定义为拐点[13]。
1.7 统计分析
肌电和肌氧采集均同步进行,为了保证时间统一以及防止延迟造成数据混乱,于受试者开始试验前10 s 统一开启肌电和肌氧设备以及秒表记录数据,受试者开始运动时记录运动开始时间和最后的结束时间。截取递增负荷运动时间段的肌电和肌氧数据,并统一以30 s 为一时间段记录均值。除了特别强调以外,所有数据均用平均值±标准误的形式呈现。相关性分析利用皮尔逊相关,单因素方差分析检验不同方法以及不同肌肉评价之间的差异性(P<0.05),克朗巴赫指数与组内相关系数(ICC)检验测得的无氧阈与乳酸阈之间的一致性(P<0.01),所有统计分析均用SPSS 22.0软件实现。
2 研究结果
2.1 肌电、肌氧、血乳酸变化趋势
试验过程中,所有受试者都完成了至少6 级递增负荷,其中16 人完成6 级递增负荷,4 人完成7 级递增负荷。负荷总持续时间为(1 163.44±66.47)s。在递增负荷试验过程中,8 块肌肉的IEMG 和RMS 都随着负荷的增加而增加,左右股外侧肌肌肉内氧含量也随着负荷增加而下降,血乳酸值随着负荷的增加而增加。
2.2 双线性拟合肌电拐点
在递增负荷较低强度时,积分肌电与均方根值都有所变化。随着负荷的增加而增加,在较大负荷强度时,积分肌电与肌电均方根值都出现显著的非线性增加,表现在大负荷强度时积分肌电和均方根值的变化速率明显高于低强度时的变化速率(见图2)。测试肌肉之间的拐点位置存在强相关(R=0.687~0.908,P<0.01),测试所有肌肉的拐点位置与乳酸阈存在强相关(R=0.685~0.908,P<0.01)(见表1),测试肌肉之间的拐点位置都没有统计学意义(P>0.05),测试所有肌肉与乳酸阈都没有统计学意义(P>0.05)。测试出的所有肌肉的积分IEMG 阈和RMS阈都要早于乳酸阈。从相关系数和一致性系数来看,股直肌和腓肠肌外侧头测得无氧阈与乳酸阈最为接近。RMS 值比IEMG 值与乳酸阈更接近,右侧的数据比左侧的数据与乳酸阈更为接近(见表2)。
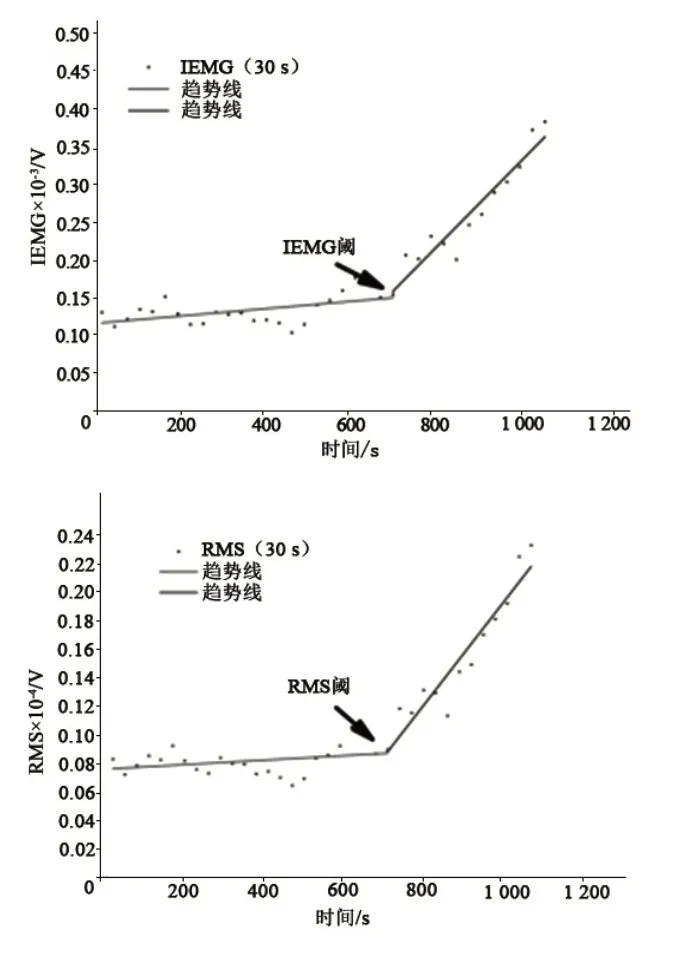
图2 受试者递增负荷中右侧股直肌肌电IEMG和RMS的拐点(典型被试)Figure2 Breakpoints of IEMG and RMS of the Right Rectus Femoris During Incremental Exercise Test(Typical Subjects)

表1 测试肌肉肌电IEMG与RMS拐点与乳酸阈的相关性以及一致性分析Table2 Correlation and Consistency Analysis Between IEMG and RMS Breakpoints and Lactate Threshold
2.3 双线性拟合肌氧拐点
在递增负荷强度较低时,肌氧下降幅度较为平缓(见图3)。随着负荷的增加而下降,在递增到一定负荷强度时,肌氧含量出现显著的非线性下降,表现在大负荷强度时肌氧的下降速率明显高于低强度时的下降速率,左右股外侧肌拐点位置之间存在强相关(R左=0.759,R右=0.809,P<0.01),左右股外侧肌的拐点位置与乳酸阈存在强相关(R左=0.762,R右=0.822,P<0.01)(见表3)。测试肌肉之间的拐点位置差异不具有统计学意义(P>0.05)。左右股外侧肌与乳酸阈都不具有统计学意义(P>0.05)。测试出的左右股外侧肌拐点位置都要早于乳酸阈。从相关系数和一致性系数来看,右侧的数据比左侧的数据与乳酸阈更为接近(见表4)。

表2 测试肌肉肌电IEMG与RMS拐点对应的功率与时间Table1 The Power and Time Estimated by IEMG and RMS Breakpoints
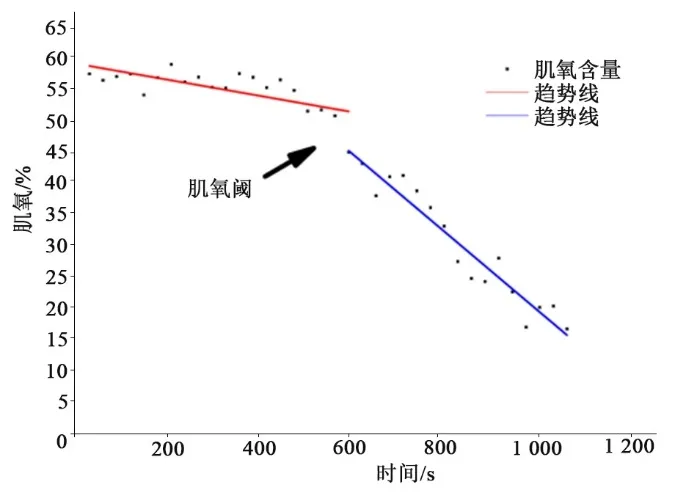
图3 受试者递增负荷中右侧股外侧肌肌氧的拐点(典型被试)Figure3 Breakpoints of Muscle Oxygenation Index of the Right Vastus Lateralis during Incremental Exercise Test(Typical Subjects)

表3 左右股外侧肌肌氧拐点与乳酸阈的相关性和一致性分析Table3 Correlation and Consistency Analysis between Muscle Oxygenation Breakpoints and Lactate Threshold

表4 左右股外侧肌肌氧拐点对应功率与时间Table 4 The Power and Time Estimated by Muscle Oxygenation Breakpoints of Tested Muscles
2.4 肌电与肌氧比值
肌电/肌氧比值法检验无氧阈,总体检出率61.1%(见图4)。通过比值法测定的左右股外侧肌肌氧和同侧测定肌肉肌电IEMG 和RMS 的比值所换算的对应无氧阈强度与乳酸阈强度不相关(P>0.1),检测出的无氧阈强度均值大于乳酸阈(见表5)。

图4 受试者递增负荷中右侧股外肌肌氧与右侧股直肌IEMG比值的拐点(典型被试)Figure4 Breakpoints of Muscle Oxygenation in the Right Vastus Lateralis/IEMG in Right Rectus Femoris during Incremental Exercise Test(Typical Subjects)

表5 左右股外侧肌肌氧与对应侧的测试肌肉肌电拐点对应功率与时间Table 5 The Power and Time Estimated by Muscle Oxygenation/EMG or RMS Breakpoints of Tested Muscles
2.5 15%肌氧拐点
15%肌氧拐点法的总体检出率43.47%,其中检测出的右侧股外侧肌与对于人数的乳酸阈强度不相关(R=0.607,P<0.05),检测出的左侧股外侧肌与对应人数的乳酸阈强度强相关(R=0.7,P<0.05),检测出的右侧股外侧肌肌氧拐点均值大于乳酸阈强度,左侧股外侧肌肌氧拐点均值小于乳酸阈强度(见表6)。

表6 左右股外侧肌肌氧拐点对应功率与时间Table 6 The Power and Time Estimated by Muscle Oxygenation Breakpoints of Tested Muscles
3 讨论与分析
3.1 双线性拟合肌电拐点
以功率为参数,以乳酸阈为标准来探讨肌电阈与无氧阈的关系时发现,测试所有肌肉的肌电阈强度与乳酸阈强度呈显著相关,分析结果认为,当运动过程中的负荷强度达到一定量时,糖酵解供能系统的加入使得肌乳酸迅速升高,肌乳酸进入血液需要2~8 min,之后使得肌乳酸和血乳酸达到平衡的状态[23],肌乳酸的升高明显快于血乳酸。肌乳酸的迅速增加使得参与工作的运动单位能力下降,为了维持正常的运动强度,便需要募集更多的快肌纤维参与运动,而此时肌细胞的内环境变化也进一步影响动作电位的传导,最终导致肌电幅值的变化。这表明肌乳酸对肌电的影响要快于血液中的乳酸堆积效应,这与本文研究中肌电阈与乳酸阈并非同时发生,各项肌电指标在各个肌肉范围内的阈值强度都要先于乳酸阈强度出现的结果一致,说明肌电阈与乳酸阈之间存在着密切关系。也有可能是肌电阈判断方法的问题,有待进一步研究。在肌肉的选择上,股直肌和腓肠肌外侧头表现出更好的与乳酸阈相关性,有文献[24]表明,自行车运动中骑行的主要发力部位在股四头肌,在蹬踏过程中,腓肠肌和胫骨前肌主要负责传递力,而股二头肌主要作用于蹬踏旋转过程中脚蹬的回收过程[25]。股直肌所测的拐点要小于腓肠肌外侧头,可能与肌肉之间的构造有关,有证据表明股四头肌处的慢肌纤维比例少于腓肠肌处的慢肌纤维比例[26],另外,股四头肌处的氧化酶活性低于腓肠肌处的氧化酶活性[27]。慢肌纤维的比例较少,以及较低的氧化酶活性,导致在递增负荷运动中股四头肌的快肌纤维将较早地被募集,从而促使肌电图谱发生变化产生拐点。
3.2 双线性拟合肌氧拐点
此方法是应用的较多的肌氧阈的确定方法,主要的生理机制是低强度运动时,局部肌肉中的氧解离速度增加,以满足运动需求的增长。但是由于运动时心血管活动增强,摄氧量增加以及局部温度升高,毛细血管开放程度增大,血流速度的加快为局部肌肉提供了足够的氧气,因此肌肉内氧含量下降不显著。但是当运动强度继续增加达到一定强度时,为了满足局部骨骼肌中的ATP 需求,肌肉氧耗急剧增加,更多的氧气从氧合血红蛋白中解离出来,PO2迅速下降从而导致了氧合血红蛋白解离加速,形成了“S”氧离曲线的第二段特征,导致肌氧含量的持续下降。
在以功率为参数,以乳酸阈为标准来探讨肌氧阈与无氧阈的关系时发现,测试的左右股外侧肌肌氧阈强度与乳酸阈强度呈显著相关,分析认为运动强度较大时,肌细胞内外各种酸性代谢产物增多,乳酸开始积累,乳酸在促进氧离中扮演着重要的角色,大强度运动过程中,局部内环境酸化,PCO2和H+浓度增高,促使O2与血红蛋白快速解离(波尔效应)[28],从而导致肌氧大幅度下降。而也如前文所述,肌乳酸的进入血液有延迟现象,而肌氧含量下降是瞬时的没有延迟,也导致测得的肌氧阈强度要先于乳酸阈。而肌电与肌氧都表现出右侧肢体拐点与乳酸阈更为接近的现象可能与实验对象的优势腿都为右腿有关。
3.3 肌电与肌氧比值法
此方法是由S.Y.KUZNETSOV[13]提出的新的联合肌电肌氧指标联合评价无氧阈强度的方法,优点是较双线性拟合要简单易行。生理学基础是随着负荷强度的上升,慢肌纤维已经不能维持运动强度,需要快肌纤维的补充。相对于有氧氧化功能来说,糖酵解供能系统开始发挥更大作用,因此之后随着负荷的增加肌肉氧耗不再剧烈增长,显示为肌氧含量到达平台期。同时肌电幅度会因为快肌纤维加入而产生急剧增加,此时建立的肌氧/肌电的比值含义是:在无氧阈之前的分子-脱氧血红蛋白含量上升,分母-肌电幅值小幅度上升,整体趋势是比值呈上升趋势。当运动到一定强度时,分子-脱氧血红蛋白含量上升出现平台,而分母-肌电幅值大幅度上升,比值不再上升而产生下降,整体的趋势线出现拐点并下降。
然而,此方法存在两点不足,首先是检出率较低,为61.1%;其次,检测出的各块肌肉的拐点强度与乳酸阈强度没有相关性。可能与很多受试者在未出现肌氧下降平台的时候已经力竭或者从始至终未出现肌氧平台有关。
3.4 15%肌氧下降法
此方法由A.C.SNYDER[16]使用的对于肌氧下降拐点的判断,优点是判定较为便利,生理学基础是肌氧到达一定负荷强度后会急剧下降的特点。然而在实验中我们发现此方法检出率低,仅为43.47%,且只有左腿肌氧拐点强度与乳酸阈强度相关。原因可能在于:首先,肌氧含量的变化容易波动出现部分极端值影响判断。其次是个体差异的存在导致很多受试者超过15%肌氧下降点并未出现,而较为严格的15%阈值的确定也使得类似下降14%这种临近值无法纳入。
4 小 结
无氧阈的检测是运动训练领域的重点研究方向之一,众多学者也在探寻可以便携,无创,简单的无氧阈检测方法。结果表明,以乳酸阈为标准,利用计算机双线性拟合技术建立的下肢各用力肌肉的肌电以及肌氧拐点与无氧阈存在着显著相关,其中肌电数据中RMS 值、肌肉选择中股直肌和腓肠肌表现出了更佳的相关性。而肌氧数据中,优势腿的股外侧肌肌氧表现出更佳的相关性。肌氧/肌电比值法和15%肌氧下降法虽然操作便利,判定简单,但是容易受到极端数据以及受试者个体差异性的影响,在无氧阈的判断上还是存在着检出率低以及不稳定的缺点。
