双硫仑及其代谢产物的抗感染药理活性研究进展
2020-11-20游雪甫杨信怡
孙 宇, 游雪甫, 杨信怡
双硫仑(disulfiram)是二乙基二硫代氨基甲酸酯(diethyldithiocarbamate,DDC)的二聚体,起初通过临床试验证明双硫仑通过抑制醛脱氢酶,阻碍乙醇在体内的代谢,致使饮酒者出现乙醛中毒样症状,并将这种症状表现命名为“双硫仑样反应”。基于这一作用,双硫仑被用作乙醇增敏药,促使嗜酒者对乙醇产生厌恶感,从而用于戒酒治疗[1-2]。当前,双硫仑作为戒酒药已被包括美国药典、日本药典、欧洲药典在内的多国药典收录。双硫仑安全性良好,口服用药量可高达500 mg/d[3]。双硫仑用于临床后,其代谢产物DDC通过调节抗氧化酶活性对抗体内过多自由基而产生的抗白内障作用也被发现,并被认为是颇具潜力的抗白内障药物[4]。近年来,研究还发现双硫仑可通过抑制肿瘤干细胞生物标志物醛脱氢酶的作用抑制肿瘤干细胞以及调节NF-κB等信号传导通路发挥抗肿瘤作用,其抗肿瘤机制的有效阐明[5-6],再次引起人们对该药的广泛关注。鉴于双硫仑曾长期作为除螨剂使用[7],因此很多研究者开始关注其在抗微生物感染领域的药用价值,进而关注其代谢产物的活性。本文针对双硫仑及其主要代谢产物在抗感染研究领域的最新研究进展进行综述,为相关领域研究者提供参考。
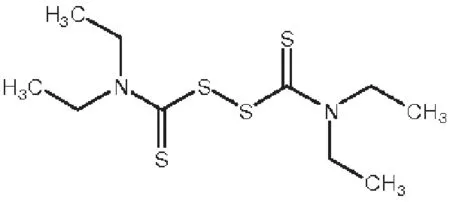
图1 双硫仑化学结构式
1 双硫仑的药动学特性
双硫仑口服后,经胃肠黏膜吸收入血,在内源性硫醇和红细胞中谷胱甘肽还原酶系统作用下迅速还原为单体DDC。后者并不稳定,可进一步降解为二乙胺(DEA)和二硫化碳(CS2)。同时,DDC也是Ⅱ相代谢酶系统的底物,在S-甲基转移酶等Ⅱ相代谢酶催化下形成二乙基二硫代氨基甲酸甲酯(Me-DDC)和葡萄糖醛酸化的DDC。Me-DDC再经氧化脱硫转化为二乙基硫代氨基甲酸甲酯(Me-DTC),Me-DTC则可被进一步氧化成其对应的砜和亚砜代谢物。吸收后,双硫仑及其代谢产物均匀分布于全身各种组织中。口服或腹腔给予大鼠或小鼠35S-双硫仑,在整个胃肠道以及血液、肝脏、肾脏、心脏、肾上腺、甲状腺、胰腺、睾丸、脾脏、骨髓和肌肉中均可检测到双硫仑和DDC,而在红细胞中未检测到放射性。然而每种代谢物在各器官中的分布差异很大,这可能与不同组织中具有的生物转化酶类型有关。双硫仑和DDC在体外和体内均可与不同蛋白质的游离巯基结合形成复合二硫化物,在血液中,双硫仑和Me-DTC主要与白蛋白高度结合。双硫仑的代谢终产物主要通过肾、粪便和肺排出。双硫仑口服后,约20.0%以原药形式经粪便排出,大多数代谢产物包括DDC的葡萄糖醛酸苷或有机硫酸盐通过肾脏消除,而CS2主要经肺排出[8]。
Faiman等[9]利用高效液相色谱(HPLC)技术在15名男性健康酗酒者中开展研究,获得了双硫仑单次和多次给药后药动学特性的详细报告。在受试者服药前和口服250 mg双硫仑片剂后,在不同时间点分别收集其血液、尿液和呼吸样本,获得单次给药样本。72 h后,受试者需每天口服250 mg双硫仑至第15天以获得多次给药样本。受试者服用单剂量或多剂量的双硫仑后,双硫仑及其代谢产物的药动学参数见表1。双硫仑、DDC、Me-DDC、DEA、CS2的消除半衰期分别为7.3 h、15.5 h、22.1 h、13.9 h、8.9 h。给药剂量为250 mg时,双硫仑及其代谢产物平均达峰时间为8~10 h。但是双硫仑及其代谢产物的血浆浓度存在明显的个体间差异,这种差异可能由于双硫仑的强脂溶性及其在脂肪组织中的分布、与血浆蛋白的结合力各不相同导致。药物一旦在脂肪组织达到“饱和”,将产生趋于一致的血浆药物水平。在对双硫仑及其代谢产物DDC的细胞和实验动物毒性研究中,未发现致癌、致畸或致突变作用[10]。
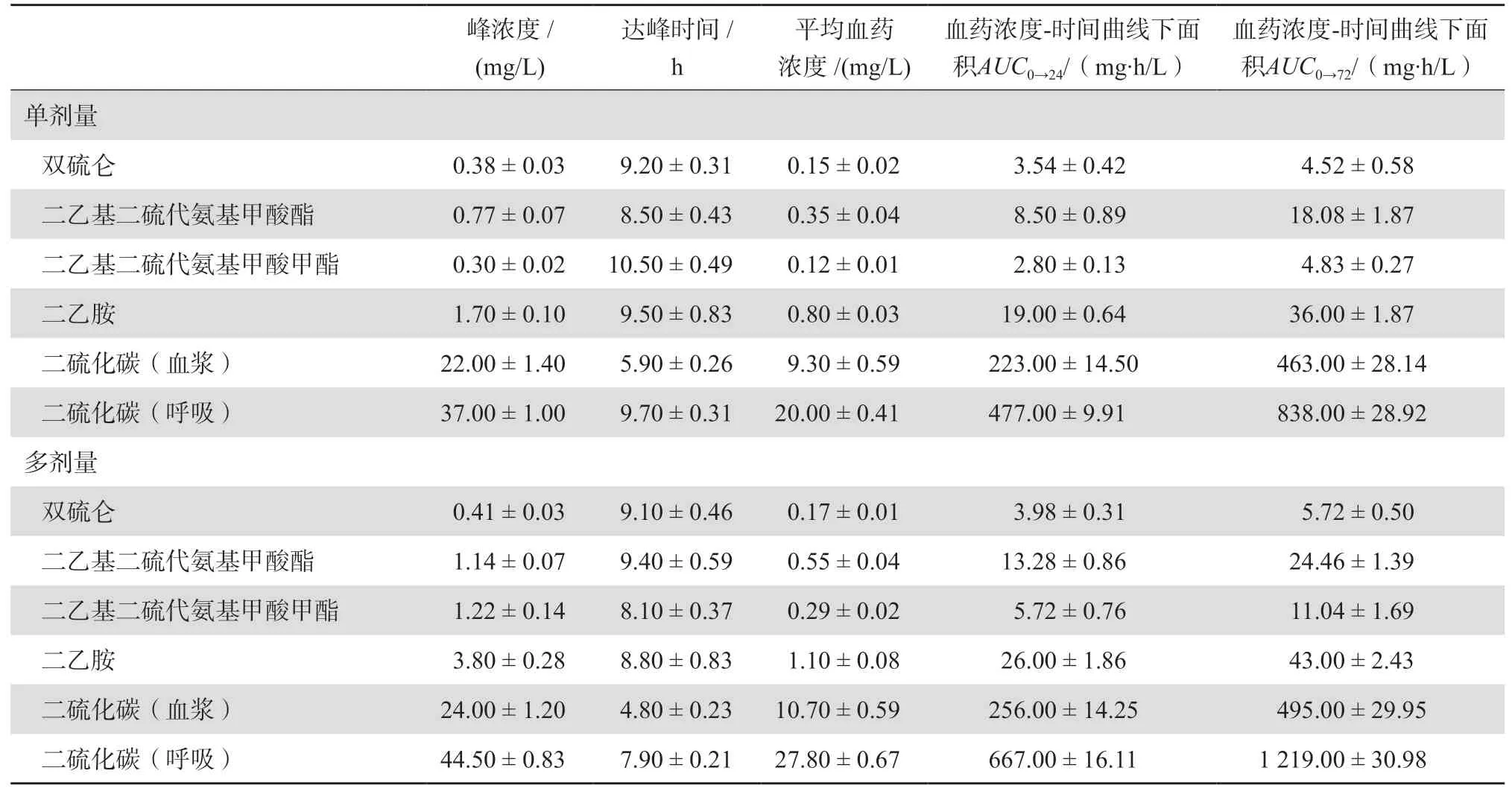
表1 双硫仑及其代谢产物的药动学参数
2 双硫仑及其主要代谢产物的抗感染药理活性
2.1 抗细菌活性
Frazier等[11]测定了双硫仑及其主要代谢产物对30种革兰阳性和阴性菌的抗菌活性。发现双硫仑对除单核细胞增生李斯特菌和链球菌之外的大多数革兰阳性菌均表现出抑菌活性。其中葡萄球菌(包括万古霉素不敏感菌株)对双硫仑的敏感性较高,最低抑菌浓度(MIC)为1~16 mg/L,而炭疽杆菌对双硫仑及代谢产物DDC均表现出很高敏感性,MIC为0.5~4 mg/L,但其余几种代谢产物未表现出明显的抗菌活性。见表2。
耐甲氧西林金黄色葡萄球菌(MRSA)是引发临床各类感染的重要病原,由于其耐多药特性,往往致使病情迁延或抗感染治疗失败。Phillips等[12]发现体外条件下,双硫仑对MRSA显示较强的抑菌活性,MIC为1.33 mg/L。之后的研究发现双硫仑除对耐多药MRSA有较好体外抑菌活性,与万古霉素联用还对万古霉素耐药金黄色葡萄球菌(VRSA)具有协同抗菌效应,在研究中测得双硫仑对临床分离的MRSA和万古霉素耐药肠球菌(VRE)的MIC范围为2~16 mg/L,对包括万古
霉素中介金黄色葡萄球菌(VISA)和VRSA菌株在内的30株受试MRSA菌株,双硫仑抑制90%菌株生长的MIC(MIC90)为16 mg/L,但革兰阴性菌对双硫仑敏感性低(MIC≥32 mg/L)[13]。棋盘法检测万古霉素和双硫仑联用,对万古霉素A型VRSA菌株具有明显的协同抗菌作用,对3株临床分离VRSA菌株,双硫仑(0.5~1 mg/L)将万古霉素的MIC从>128 mg/L降至4~16 mg/L,抑菌浓度(FIC)指数 <0.5。这些研究提示双硫仑或可作为万古霉素增效剂或联用药物,用于糖肽类药物治疗无效的慢性和复发性MRSA感染,其在体内的联用效果值得进一步关注。
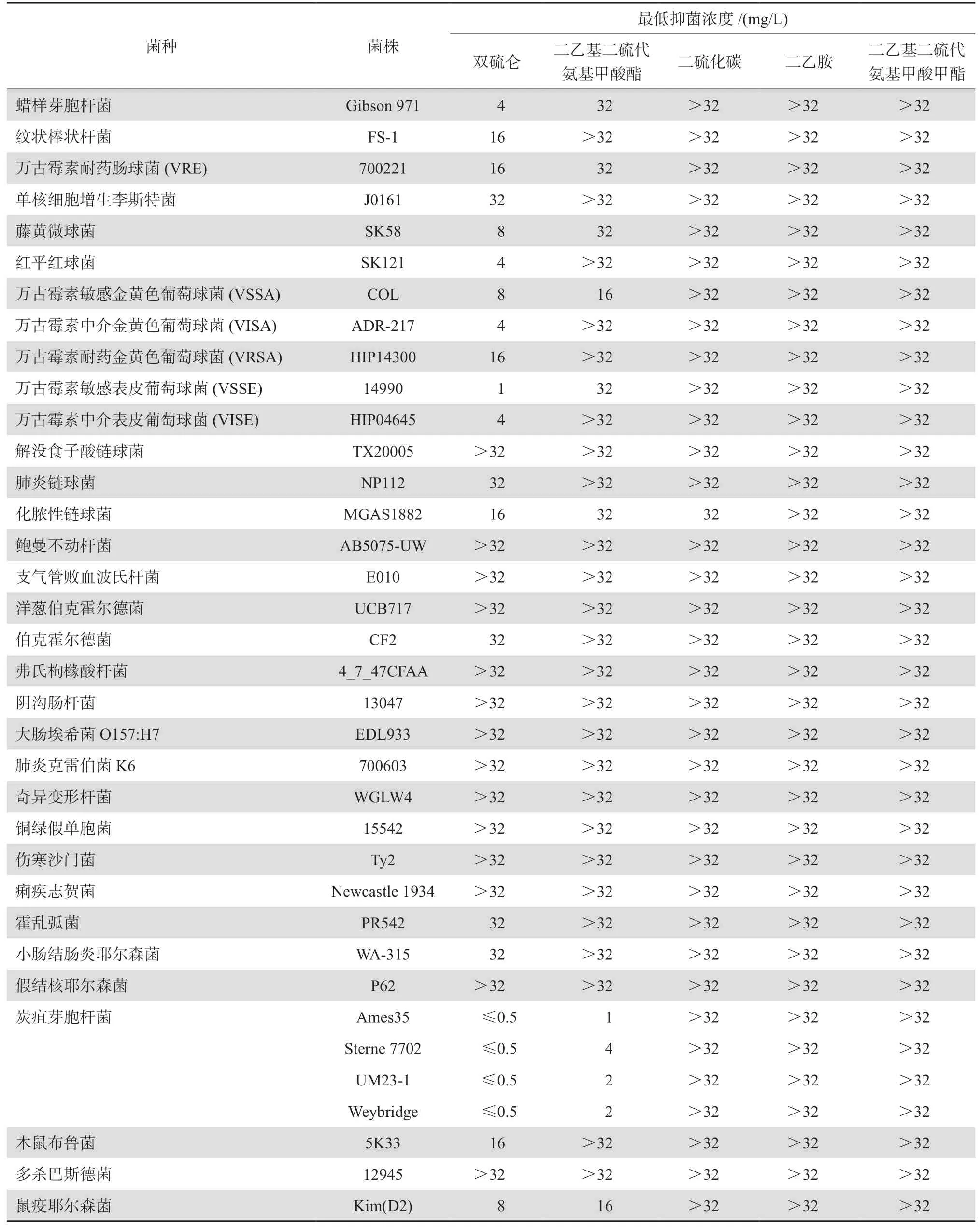
表2 双硫仑及其代谢产物的抗菌活性
通过检测双硫仑及代谢产物在动物体内的分布,证实双硫仑和DDC优先分布于肺部,这为双硫仑治疗某些感染性肺病如肺结核提供了依据[14]。早先,Jeney等[15]首次报道了双硫仑和DDC在豚鼠结核分枝杆菌(Mycobacterium tuberculosis,Mtb)感染模型中的抗菌活性。Hubner等[16]进一步观察到DDC在体外和体内条件下均可增强单核细胞的抗结核活性。在评价双硫仑和DDC对耐多药Mtb(MDR-Mtb)和泛耐药Mtb(XDR-Mtb)菌株的抗菌活性中,对包括MDRMtb和XDR-Mtb菌株在内的42株临床分离Mtb菌株,双硫仑和DDC均显示良好抗Mtb活性,MIC90分别为1.56 mg/L和3.13 mg/L,且与喹诺酮类药物(左氧氟沙星、司帕沙星、环丙沙星)无交叉耐药性[17]。随后,研究者利用人单核细胞系THP-1检测双硫仑和DDC细胞内杀Mtb的活性,发现二者以剂量依赖方式分别在6~30 mg/ L和10~30 mg/ L对细胞内Mtb呈现明显杀菌活性。在慢性结核病(TB)小鼠模型中,双硫仑亦显示较明显的体内杀菌活性。双硫仑治疗组在80~160 mg/kg剂量时,小鼠肺和脾中菌落形成单位计数值(CFU)从105CFU/脏器降低至104CFU/脏器。尽管目前双硫仑抗Mtb的作用机制仍不清楚,但有研究显示双硫仑结构类似物二硫代氨基甲酸盐类可能通过抑制β碳酸酐酶(β-CAs)活性抑制Mtb的生长,该作用机制对于寻找双硫仑及DDC的抗Mtb靶标具有一定启示意义。总之,双硫仑及其代谢物DDC可能是一类潜在的治疗MDR / XDR-TB的新型药物,其抗Mtb作用机制值得进一步关注。
2.2 抗真菌活性
免疫功能低下患者如艾滋病、癌症晚期、重症监护患者易发生真菌感染,目前常用的抗真菌药往往因真菌的跨膜外排机制[如P-糖蛋白(P-gp)介导的药物外排]而活性降低[18]。研究显示,双硫仑对真菌细胞膜表面的P-gp有抑制调节作用。Sauna等[19]对双硫仑作为多药耐药调节剂的分子基础进行探索,发现双硫仑能与多种ATP结合转运蛋白的药物-底物结合位点结合并抑制耐药性相关的ATP水解。以抗深部真菌的三唑类药物为例,真菌对其产生耐药性主要由P-gp介导。而双硫仑作为一种外排泵调节剂,可通过抑制这种药物外排克服真菌对三唑类药物(主要是氟康唑耐药白念珠菌)的耐药性,从而可作为一种潜在的联用药物,辅助治疗耐药性真菌引起的感染。
受此启发,Khan等[20]采用氟康唑联合双硫仑对白念珠菌和光滑念珠菌进行抗真菌测试,然而并未观察到两药存在任何协同或相加作用,他们推测可能是由于14-α-脱甲基酶的突变所致。之后,他们又对双硫仑的抗一系列酵母和丝状真菌活性进行了检测。研究显示,双硫仑对61株受试氟康唑敏感或耐药的白念珠菌、非白念珠菌、丝状真菌分离株均显示良好杀真菌活性,MIC50、MIC90分别为4 mg/L、8 mg/L。双硫仑对试验中13株曲霉也呈现明显的杀真菌活性,MIC50、MIC90分别为2 mg/L、8 mg/L。尽管目前人们已观察到双硫仑具有较显著的杀真菌活性,但其作用机制仍待阐明。
2.3 抗寄生虫活性
早在1942年,Gordon等[7]就提出四乙基结构类似物应具有杀螨作用,进而利用背肛螨属螨虫的感染大鼠模型研究了双硫仑的杀螨作用,并与2种疥疮治疗药物苯甲酸苄酯和二甲基噻吩(灭疥)进行了疗效比较。结果显示,双硫仑的杀螨作用较2种对照药更为高效、迅速,应是一种有效的疥疮治疗药物。这一发现为双硫仑用于疥疮治疗提供了依据,并启发人们进一步探索其治疗其他寄生虫感染的可能。双硫仑上市后,人们对其抗寄生虫活性的研究继续深入,后续研究显示其对利什曼虫、蓝氏贾第鞭毛虫(贾第虫)、恶性疟原虫、克氏锥虫、阴道毛滴虫等具有广谱杀虫活性。
利什曼虫感染可导致内脏或皮肤利什曼虫病,且一旦引起内脏利什曼虫病,如未及时接受治疗往往危及生命。Osorio等[21]采用荧光素酶转染的杜氏利什曼原虫,对仓鼠的离体脾脏进行感染制成外植体培养系统,筛选抗虫剂。最终从4 000多个化合物中筛得双硫仑,发现其在纳摩尔浓度具有显著杀利什曼原虫活性。Peniche等[22]通过进一步研究,发现双硫仑与二价金属盐(1 μmol/ L)合用,能显著提高杀灭利什曼原虫活性。例如,双硫仑与硫酸锌组合可使针对杜氏利什曼原虫的体外治疗指数增加1.8倍。研究人员通过对利什曼虫进行代谢网络计算机模拟评估,预测双硫仑的抗利什曼原虫活性可能是与其干扰虫体中由LmjF25.1170和LmjF25.1180基因编码的跨膜质子转运系统有关。这些基因编码的蛋白质是F型ATP酶β链复合物组成部分,参与寄生虫ATP合成,通过特异性跨膜转运质子(H+)产生电化学梯度,从而为ATP合成提供动力。在杜氏利什曼虫中,双硫仑导致F-ATP合成酶的抑制使线粒体膜电位去极化,ATP合成受损,从而增加细胞活性氧的产生,导致寄生虫DNA断裂而死亡。尽管目前尚无单用或联用双硫仑治疗皮肤或内脏利什曼病的临床数据,但这些研究确为双硫仑未来用于利什曼病的治疗奠定了理论基础。
贾第虫是一种常致人体肠道感染的寄生虫,可致贾第虫病。Nash等[23]发现体外条件下测得(1.23±0.32)μmol/L双硫仑可杀灭贾第虫。在该寄生虫感染的小鼠模型中,研究者进一步观测到每天2次,持续4 d灌胃给予双硫仑25 mg,可使40.0%小鼠治愈,未治愈小鼠的贾第虫负荷也有显著性降低(P<0.01),而对照组治愈率为0。Hill等[24]在体外、体内条件下观测了双硫仑对寄生性鼠鞭虫卵壳形态的影响。研究人员在小鼠感染鼠鞭虫后第25天开始每天分别给予剂量为2.5 mg/kg、5.0 mg/kg、7.5 mg/ kg的双硫仑,治疗26 d后,观察到治疗组所有成年雌性鞭毛虫所产虫卵均为畸形卵。此外,与对照组小鼠比较,5.0 mg·kg-1·d-1、7.5 mg·kg-1·d-1剂量的双硫仑治疗组小鼠尸检中发现的成虫数量显著降低(P<0.05)。一般认为,鼠鞭虫的卵壳可保护虫卵在发育过程中免受外界伤害,在卵壳形成过程中,联苯酚氧化酶发挥着重要作用。研究者推测,双硫仑作为一种氧化抑制剂,可能抑制该酶活性进而干扰虫卵孵化为成虫。双硫仑通过干扰卵壳形成,阻断由卵发育为鞭虫成虫的杀虫方式具有重要临床意义。
此外,大量报道还显示双硫仑和/或其代谢产物在体外对恶性疟原虫、克氏锥虫、阴道毛滴虫也具有抑制活性。Scheibel等[25]最早注意到双硫仑和DDC的抗疟活性,他们认为这两个化
合物具有的抑制多种金属蛋白酶加氧酶的活性、良好的脂/水分配系数、与金属离子的高结合常数等,均有利于针对疟原虫发挥选择性毒性。体外研究结果显示,双硫仑和DDC在浓度仅为0.1 mg/L时即表现出明显的抗疟活性。在该研究基础上,Meshnick等[26]还发现DDC可通过与外源性铜离子或内源性铜离子(存在于宿主细胞质和寄生虫溶酶体中负载超氧化物歧化酶的红细胞中)形成复合物,导致虫体细胞膜损伤,进而发挥抗疟作用。不过,DDC-Cu复合物诱导膜损伤的具体机制,目前仍不清楚。Lane等[27]在体外检测了双硫仑及其还原代谢产物二乙胺-N-二硫代碳酸钠(DECD)对克氏锥虫包括上鞭毛体、锥鞭毛体、无鞭毛体形式的杀灭活性。结果显示,在50 mg/L浓度下,双硫仑和DECD对上鞭毛体的抑制率分别为64.6%和69.7%,对组织培养锥鞭毛体的抑制率分别为 47.7%和46.1%,对无鞭毛体感染的3T3成纤维细胞抑制率分别为60%和67%。研究者同时指出双硫仑和DECD杀锥虫的潜在机制与干扰克氏锥虫的必需金属离子代谢有关,并认为二者均可作为预防或治疗锥虫病的药物候选物。另外,双硫仑和DDC对甲硝唑敏感及耐药的阴道毛滴虫和胎三毛滴虫也具有明显抑制活性,双硫仑和DDC对甲硝唑敏感、耐药阴道毛滴虫的MIC分别为0.1~0.7 μmol/ L、0.2~1.3 μmol/ L和0.3~9 μmol/ L、1.2~9 μmol/L;双硫仑和DDC对甲硝唑敏感、耐药的胎三毛滴虫的MIC分别为0.1~1.0 μmol/L、1.0~6.9 μmol/L和0.3~1.3 μmol/L、0.6~6 μmol/L[28]。总之,上述多项研究均显示双硫仑及其代谢产物在治疗寄生虫感染方面具有巨大应用潜力。
2.4 抗病毒活性
1975年,Carić-Lazar等[29]报道了双硫仑可影响包膜病毒的增殖。在病毒吸附前1 h和吸附期间加入双硫仑(0.1~0.3 mmol/L),对塞姆利基森林病毒(semliki forest virus,SFV)、禽瘟病毒(fowl plague virus)、新城疫病毒(newcastle disease virus)、水疱性口炎病毒(vesicular stomatitis virus)、伪狂犬病病毒(pseudorabies virus)的增殖均有明显抑制作用。不过,在病毒吸附至鸡胚细胞后再加入药物,则不能观察到病毒增殖受抑的现象。双硫仑既不干扰病毒的受体也不干扰红细胞,并且不会阻止病毒吸附,故研究者认为双硫仑可能影响病毒繁殖的早期步骤。
联合抗反转录病毒疗法(cART)是一种抑制人类免疫缺陷病毒(HIV)复制、延缓病情发展的有效治疗方案,但这一疗法无法根除病毒感染,主要是因为HIV的基因组可通过反转录过程稳定整合入宿主细胞如记忆性CD4+T细胞的染色体中,进入潜伏状态,表现为长期的转录沉默[30]。针对此现象,当前的主要治疗策略是重新激活潜伏的HIV,诱导受感染的细胞继续产生病毒颗粒,从而使受感染的宿主细胞死亡或诱导HIV特异性T 细胞杀伤。Xing等[31]用抗凋亡蛋白Bcl-2转染的原代CD4+T细胞模型高通量筛选能诱导病毒基因表达但不触发细胞活化的化合物,发现双硫仑可重新激活潜伏感染的HIV-1而不引起总体T细胞的活化。目前认为,双硫仑通过消耗细胞内磷酸酶和张力蛋白同源体(PTEN)上调蛋白激酶B(Akt)信号通路,从而诱导HIV-1潜伏感染的单核细胞系中HIV-1基因转录[32]。在此基础上,Spivak等[33]在临床试验中,连续14 d给予HIV-1患者双硫仑500 mg/ d,观察到该药确实可以重新激活人体静息记忆CD4+T细胞中潜伏的HIV-1。随后,Elliott等[34]又进行了一项剂量递增研究,他们对30例接受cART治疗的AIDS患者,连续3 d每天分别给予500 mg、1 000 mg和2 000 mg剂量的双硫仑,结果导致所有剂量组受试者均出现了细胞相关的未拼接 HIV RNA增加,且在高剂量下观察到血浆HIV RNA的增加,与激活HIV潜伏期一致。该试验同时发现,即使双硫仑用药量达到常规解酒用药量的4倍,患者依然耐受良好。鉴于其良好的安全性表现,双硫仑应该适用于长期、联合给药,用于cART中潜伏HIV的激活。
鉴于双硫仑能与铜离子螯合,Levinson等[35]研究了双硫仑及其代谢产物DDC对劳氏肉瘤病毒恶性转化、真核细胞合成和核酸结合的影响。结果发现,双硫仑可抑制劳氏肉瘤病毒RNA依赖性DNA聚合酶的活性,并使病毒丧失恶性转化鸡胚细胞的能力,但DDC对病毒无明显影响。双硫仑和DDC诱导正常鸡胚和人类包皮细胞中四种蛋白质的合成,当铜离子存在时,DDC可与HeLa细胞DNA和劳氏肉瘤病毒70S基因组RNA结合。研究者认为,双硫仑可能是通过与DNA聚合酶活性位点中的Zn2+结合而对病毒粒子DNA聚合酶产生抑制,从而抑制劳氏肉瘤病毒转化活性。
病毒蛋白中,Zn2+是维持天然蛋白质结构稳定的关键辅助因子。某些与Zn2+结合的半胱氨酸可与亲电试剂反应,致使Zn2+失去配体而被排出,从而导致天然蛋白质结构和功能受到破坏。故病毒的某些重要结构或功能蛋白中,这类含有反应性锌结合半胱氨酸残基的不稳定锌位点可能成为一种颇具希望的抗反转录病毒治疗靶标。双硫仑作为一种经临床证明、安全性颇高的锌驱逐剂,显然可通过靶向锌位点而成为一种潜在的抗病毒药物。Lee等[36]在丙型肝炎病毒(HCV)的NS5A蛋白中鉴定出不稳定的锌位点,并证明双硫仑可以从预测的靶位将Zn2+驱除,从而产生抑制HCV复制的活性,且抑制程度与抗病毒药物利巴韦林相似,由此认为,双硫仑与干扰素和/或直接靶向HCV的抗病毒药物联用可以治疗感染HCV并伴有酒精依赖的患者。Novo-Veleiro等[37]也指出,由于饮酒和HCV感染具有协同肝毒性作用,两种因素共存可增加晚期肝硬化和患肝癌的风险,因此对存在酒精依赖的HCV感染患者采用双硫仑治疗,可能会对抗病毒疗效和患者依从性产生积极影响,为治疗提供额外支持。此外,Chen等[38]通过RNA干扰基因组文库筛选,发现ZBTB25是多种细胞基因的转录抑制因子,涉及ZBTB25的蛋白质-蛋白质和蛋白质-RNA相互作用促进病毒RNA转录和复制,而ZBTB25相关功能需要完整的锌指结构域和ZBTB25的翻译后SUMO-1修饰。研究者发现用双硫仑处理过表达ZBTB25的细胞,由于RNA合成减少导致甲型流感病毒的产生显著降低,双硫仑破坏锌指结构的功能有效抑制了甲型流感病毒复 制。
另外,对于具有高致病性的严重急性呼吸综合征冠状病毒(SARS-CoV)和中东呼吸综合征冠状病毒(MERS-CoV),双硫仑对两者的木瓜蛋白酶样蛋白酶(PLpros)分别产生竞争性抑制和别构抑制作用。除去未结合的双硫仑后,病毒出现的缓慢结合抑制现象和酶活性的不可恢复性,表明双硫仑很可能通过共价结合灭活了SARS-CoV PLpro,而双硫仑和6-巯基鸟嘌呤或霉酚酸联用对MERS-CoV PLpro显示出的协同抑制作用则提示这3种临床药物联合使用,可能具有治疗冠状病毒感染的潜在价值[39]。
3 结语
双硫仑药用历史悠久,生物利用度高(80.0%),安全性良好。在当今药物研发领域,作为“老药新用”策略中的一个代表,双硫仑及部分代谢产物表现出广泛的药理学活性,具有极高的药用研究价值和发掘潜力。在抗感染治疗研究领域,双硫仑单用或与其他药物联用,在抑制耐药金黄色葡萄球菌、Mtb生长,杀灭多种真菌、寄生虫,抑制病毒转录、复制、激化等方面展现出良好活性,为其作为抗感染药物的进一步开发提供了可能。不过,与双硫仑抗感染相关的许多作用机制仍不明确,因此还需基于更多药理研究模型,提出合理科学假说,深入探索其有效作用靶标,并进行可能的结构改造,为其成为一类新型抗感染药物提供依据。
