黔北麻羊不同组织中FABP1、FABP3 基因表达水平的研究
2020-11-18艾锦新龙安炬罗卫星蔡惠芬
艾锦新,龙安炬,罗卫星,蔡惠芬*
(1.贵州大学动物科学学院,贵州贵阳 550025;2.高原山地动物遗传育种与繁殖教育部重点实验室,贵州贵阳 550025)
脂肪酸结合蛋白(Fatty Acid Binding Proteins,FABPs)是胞内脂质结合蛋白超家族成员[1-2],参与细胞内脂肪酸的运输和体内脂肪的合成与降解[3],对细胞信号传导、增殖及分化具有重要的调控作用[4-5]。目前为止,至少已经在哺乳动物中发现9 种FABPs,按发现时间的前后顺序和表达的组织特异性,可分为FABP1(肝脏型)、FABP2(小肠型)、FABP3(心脏型)、FABP4(脂肪型)、FABP5(表皮型)、FABP6(回肠型)、FABP7(脑型)、FABP8(髓鞘型)和FABP9(睾丸型)[6]。FABP1 主要在哺乳动物肝脏和小肠中表达[7];FABP3主要在哺乳动物心肌、骨骼肌和脂肪细胞中表达[8]。俞英等[9]研究发现,FABP1基因在杂种鸡CD(正交组合鸡)、DC(反交组合鸡)肝脏组织中的表达量均明显高于亲本纯种鸡CC(丝羽乌鸡)和DD(农大褐蛋鸡),由此推测FABP1基因高表达导致杂种鸡的脂肪沉积高于亲本纯种鸡。高妍等[10]研究发现,FABP1基因外显子2 的SNP 位点与松辽黑猪的大理石纹和肌内脂肪含量显著相关。朱梦婷等[11]研究发现,FABP3基因在黄羽肉鸡的胸肌、腿肌和腹脂中均有不同程度的表达。岳彩娟等[12]研究发现,FABP3基因表达量与滩羊股二头肌、背最长肌及腰大肌的肌内脂肪(IMF)含量呈正相关。
黔北麻羊是贵州优良的地方山羊品种[13-14],但与国外优质肉羊相比仍存在个体偏小、产肉性能低等缺点。本研究以黔北麻羊为研究对象,利用qRT-PCR 技术检测FABP1、FABP3基因在各个组织中的表达水平,以期丰富黔北麻羊肉质性状相关基因的遗传信息数据,为进一步探究FABP1、FABP3基因在羊生长发育和脂肪沉积中的调控作用提供理论参考。
1 材料与方法
1.1 样品采集 实验动物来源于贵州省习水县富兴牧业有限公司,选取6 月龄、1 周岁、1.5 周岁饲养管理条件相同且健康无病的公、母黔北麻羊各3 只,在现场屠宰后快速采集各个组织(心脏、肝脏、脾脏、肺脏、肾脏、胃脏、下丘脑、十二指肠、胸腺、背最长肌和皮下脂肪),分别用DEPC 水清洗上述11 个组织,随后置于装有RNA 保存液的冻存管中,投入液氮罐内快速冷冻后带回实验室-80℃冰箱长期保存备用。
1.2 试剂与仪器 主要试剂:Trizol 试剂盒购自大连宝生物工程有限公司,逆转录试剂盒和SYBR Green(ROX qPCR mix)荧光染料Mix 购自Thermo 公司,2×Taq PCR MasterMix、琼脂糖凝胶回收试剂盒购自北京鼎国生物技术工程有限公司,无水乙醇购自天津富宇精细化工有限公司,氯仿购自重庆川东化工有限公司。主要仪器:普通梯度PCR 扩增仪、RT-PCR 仪(CFX96 型)均购自Bio-Rad 公司,DYY-2C 型电泳槽、电泳仪购自北京六一仪器厂,超微量紫外分光光度计(NanoDrop-2000型)购自Thermo 公司。
1.3 组织总RNA 提取与反转录 根据TaKaRa 公司Trizol 试剂盒说明书,分别提取黔北麻羊各组织的总RNA,用1% 琼脂糖凝胶电泳检测RNA 的完整性,将各组织RNA 浓度稀释到1 000 ng/ μL,置于-80℃冰箱中保存;反转录合成黔北麻羊各组织的cDNA,经超微量紫外分光光度计检测其cDNA 的浓度及完整性,保存于-20℃冰箱中备用。
1.4 引物的设计与合成 参考GenBank 中山羊FABP1基因(登录号:NM_001287558.1)和FABP3基因(登录号:NM_001285701.1)的mRNA 序列设计qRT-PCR特异性引物,进行黔北麻羊FABP1和FABP3基因RTPCR 扩增,以甘油醛-3-磷酸脱氢酶(GAPDH)作为内参基因。利用Primer5.0 软件设计引物,引物序列信息见表1。引物均由成都擎科梓熙生物技术有限公司合成。
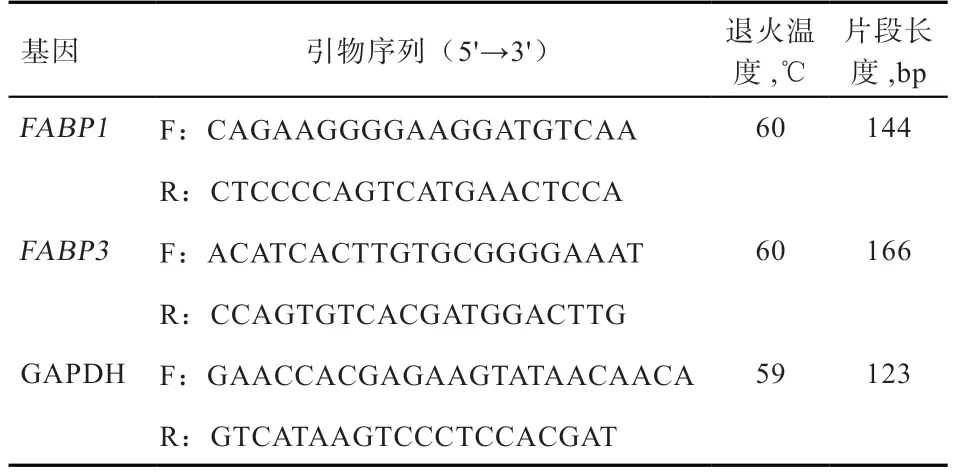
表1 引物序列
1.5 实时荧光定量PCR 普通PCR 和荧光定量PCR 检测荧光引物特异性,摸索最佳反应温度和程序。采用SYBR Green 荧光染料法,PCR 反应体系为10 μL:赛默飞SYBR Green(ROX qPCR mix)5 μL,上、下游引物各0.4 μL,cDNA 模板1 μL,ddH2O 3.2 μL。PCR反应条件:50℃ 2 min,95℃ 2 min,95℃ 15 s,60℃15 s,72℃ 1 min,共40 个循环;循环40 次后进行熔解曲线分析:95℃ 5 s,65℃ 5 s,温度以每5 s 递增0.5℃的速率从65℃上升到95℃。每个待测样品做3 个重复。
1.6 统计分析 从荧光定量PCR 仪中导出实验数据,运用Excel 2016 进行数据的整理与统计;采用2-△△Ct方法计算FABP1、FABP3基因在黔北麻羊不同组织中的相对表达量;利用SPSS 22.0 软件进行实验数据的统计分析,P<0.05 表示差异显著,结果用平均值±标准误表示。
2 结果与分析
2.1 总RNA 及PCR 检测结果 用1% 的琼脂糖凝胶电泳检测黔北麻羊不同组织总RNA,结果见图1,总RNA 条带清晰明亮,说明RNA 纯度较高、完整性较好。选取黔北麻羊肝脏和心脏组织样cDNA,经PCR 扩增后以1%的琼脂糖凝胶电泳检测其产物,得到FABP1、FABP3、GAPDH基因目的条带与预期相符,可用于下一步实验。
2.2 不同性别黔北麻羊FABP1基因的组织表达差异如图2 所示,FABP1基因在黔北麻羊不同组织中均能检测到表达,以脾脏组织为对照,FABP1基因在黔北麻羊肝脏中的表达量最高,其次是十二指肠和肺脏,而在心脏、脾脏、肾脏、胃脏、背最长肌、皮下脂肪、胸腺、下丘脑中的表达量都较低。不同性别中,FABP1基因的表达量存在一定差异,母羊肝脏、十二指肠、皮下脂肪组织中FABP1基因的表达量显著高于公羊,但在母羊背最长肌、下丘脑组织中FABP1基因的表达量却显著低于公羊,在其他组织中公、母羊FABP1基因的表达量均差异不显著。
2.3 不同生长时期黔北麻羊FABP1基因的组织表达差异 根据实时荧光定量PCR 实验数据分析(表2),不同生长时期黔北麻羊FABP1基因的组织表达量存在显著性差异,以脾脏组织为对照,6 月龄公、母羊心脏、肝脏组织中FABP1基因的表达量显著低于1 周岁和1.5周岁,1 周岁和1.5 周岁之间差异不显著。1.5 周岁公、母羊脾脏、下丘脑组织中FABP1基因的表达量显著高于6 月龄和1 周岁,6 月龄和1 周岁差异不显著。6 月龄、1 周岁和1.5 周岁母羊肺脏、肾脏、胃脏、十二指肠、皮下脂肪中FABP1基因的表达量无显著差异。公羊肺组织中FABP1基因的表达量显著高于1.5 周岁,6 月龄与其他2 个时期差异不显著;公羊肾脏中FABP1基因的表达量显著高于6 月龄和1.5 周岁,6 月龄和1.5周岁差异不显著;6 月龄公羊十二指肠中FABP1基因的表达量显著低于1 周岁和1.5 周岁,1 周岁和1.5 周岁差异不显著;1.5 周岁公羊皮下脂肪、胃中FABP1基因的表达量显著高于6 月龄和1 周岁。1 周岁公、母羊胸腺组织中FABP1基因的表达量显著高于6 月龄和1.5周岁,6 月龄和1.5 周岁差异不显著。不同生长时期公、母羊背最长肌中FABP1基因的表达量无显著性差异。
2.4 不同性别黔北麻羊FABP3基因的组织表达差异如图3 所示,FABP3基因在黔北麻羊不同组织中均能检测到表达,以背最长肌为对照,FABP3基因在黔北麻羊心脏中的表达量最高,其次是肾脏和背最长肌,而在肝脏、脾脏、肺脏、十二指肠、皮下脂肪、胃脏、胸腺、下丘脑中表达量都较低。不同性别中,FABP3基因的表达量存在一定差异,母羊心脏、皮下脂肪、胸腺组织中FABP3基因的表达量显著高于公羊,但在母羊肾脏组织中FABP3基因的表达量显著低于公羊,在其他组织中公、母羊FABP3基因的表达量差异不显著。
2.5 不同生长时期黔北麻羊FABP3基因的组织表达差异 由表3 可见,不同生长时期黔北麻羊FABP3基因的组织表达量存在显著差异,以背最长肌为对照。6 月龄公、母羊心脏、肾脏、肝脏、胃脏、十二指肠及背最长肌中FABP3基因的表达量均显著低于1 周岁和1.5周岁,1 周岁和1.5 周岁差异不显著;6 月龄、1 周岁和1.5 周岁公、母羊脾脏中FABP3基因的表达量差异不显著;1.5 周岁黔北麻羊公、母羊皮下脂肪、胸腺中FABP3基因的表达量显著高于6 月龄和1 周岁,6 月龄和1 周岁差异不显著;1.5 周岁公、母羊肺脏、下丘脑中FABP3基因的表达量显著高于6 月龄,1 周岁与6 月龄、1.5 周岁无显著性差异。

表2 不同生长时期黔北麻羊FABP1 基因在各组织中的差异表达

表3 不同生长时期黔北麻羊FABP3 基因在各组织中的差异表达
3 讨 论
肌内脂肪含量作为影响畜禽肉质的主要因素之一,增加肌内脂肪含量可以改善肉质风味。而FABPs 能特异性地结合游离脂肪酸和其他疏水基团,将细胞内脂肪酸运送到甘油三酯和磷酯合成或分解的部位,促进脂化反应的进行及甘油三酯的重新合成,从而增加肌内脂肪含量[5]。FABPs 基因编码脂肪酸结合蛋白,而脂肪酸结合蛋白是影响脂肪沉积的重要基因,对机体脂肪代谢具有重要调控作用[7]。因此,通过探究FABP1和FABP3基因的组织表达规律,运用分子生物技术手段提高肌内脂肪含量,可以在保持较高产肉性能的前提下提高肉品质。目前,对于FABP1和FABP3基因的研究大多集中在猪、牛、鸡等动物上,而在山羊的研究鲜见报道。本研究发现,FABP1和FABP3基因在黔北麻羊不同组织中均有表达,其中FABP1基因在黔北麻羊肝脏中相对表达量最高,其次是十二指肠和肺脏;FABP3基因在黔北麻羊心脏中相对表达量最高,其次是背最长肌和肾脏;在不同性别黔北麻羊各组织的表达中,FABP1和FABP3基因在黔北麻羊母羊各组织中的表达量普遍高于公羊;在不同生长时期黔北麻羊各组织的表达中,FABP1和FABP3基因表达量随黔北麻羊月龄的增加呈现小幅度上升趋势。
对FABP1基因在黔北麻羊各组织的表达情况进行分析,发现FABP1基因在肝脏和十二指肠的表达最为丰富,这与Hittel 等[15]研究FABP1基因在小鼠肝细胞和小肠黏膜细胞胞质中高丰度表达的研究结果相一致。姜延志等[16]研究发现,在猪各个组织中均能检测到FABP1基因的表达,但其在肝脏组织中的表达量最高。石慧等[17]采用Westernblot 技术对鸡FABP1基因的组织表达特性进行分析,结果仅检测到FABP1基因在肝脏和小肠中有较高表达,在其他组织未检测到表达。结合本研究结果,FABP1基因在肝脏的表达最丰富与前面研究结果相符,这可能是与肝脏需要FABP1参与脂肪酸的摄取、运输以及长链脂肪酸转化为甘油三酯有关[15]。对不同生长时期黔北麻羊背最长肌的FABP1基因表达情况进行分析,发现FABP1基因的表达量随着年龄的增加缓慢上升,这与郑程莉[18]研究FABP1基因在天府肉羊背最长肌呈下降-上升-下降的表达结果存在差异,可能是由于分组差异、羊种遗传基础及饲养环境不同所致。在整个生长阶段中,FABP1基因的表达量均处在一个较低水平,这可能与FABP1对软脂酸盐、油酸和硬脂酸盐等长链脂肪酸具有高度亲和力[15],参与脂肪酸的摄取和转运会降低其表达量相关。
对FABP3基因在黔北麻羊各组织的表达进行分析,发现FABP3基因在心脏的表达量最高,其次是背最长肌,这与胡江等[19]研究FABP3基因在甘南牦牛各组织的相对表达量结果相一致。但韩英等[20]研究发现,FABP3基因在兴凯湖翘嘴鲌的肝脏和胰脏中表达量最高,这与本实验结果存在差异,可能是由于物种跨度过大所致。赵雪等[21]研究绵羊FABP3基因的组织表达情况发现,FABP3基因在腿部半腱肌、背最长肌中表达量最高,其次才是心脏;黄河等[22]研究发现,FABP3基因在藏猪心脏的表达量高于背最长肌,背最长肌的表达量高于肝脏;邓龙华等[23]研究发现,FABP3基因在鸡心脏的表达量为最高,其次是腿肌;周泉勇等[24]研究发现FABP3基因主要在黑玉山猪心、肾、脂肪和骨骼肌组织中表达,且在心脏组织中的表达量最高。综合上述研究成果并结合本研究结果分析,FABP3基因在心脏的表达量最高,其次是骨骼肌,肝脏表达量最低,说明FABP3基因具有组织表达特异性。对FABP3基因在不同生长时期黔北麻羊背最长肌的表达情况进行分析,发现FABP3基因的表达量随着年龄的增加缓慢上升,这与郝称莉等[25]研究FABP3基因在湖羊背最长肌呈上升-下降-上升的表达结果不同,可能是由于年龄段差异、品种及饲养环境不同造成。同时,本研究发现FABP1、FABP3基因在不同性别黔北麻羊背最长肌中的表达量存在显著差异,公羊FABP1基因的表达量显著高于母羊,母羊FABP3基因的表达量显著高于公羊,这与孙伟等[26]研究GHR 和IGF-1 基因表达在湖羊肌肉中的发育性变化存在相似之处,说明了性别和年龄对基因在肌肉中的表达具有重要影响。从FABP1基因在肝脏和十二指肠表达量最高、FABP3基因在肝脏和十二指肠表达量较低、FABP3基因在心脏和背最长肌表达量最高、FABP1基因在心脏和背最长肌表达量较低的表达规律来看,FABP1基因和FABP3基因之间可能存在拮抗作用,但还有待进一步研究。
4 结 论
本研究结果表明,FABP1、FABP3基因广泛表达于黔北麻羊各组织中,其中FABP1基因在肝脏表达量最高,在心脏表达量极少,FABP3基因在心脏表达量最高,在肝脏表达量极少,推测二者可能存在拮抗作用。在不同性别中,母羊不同组织的FABP1、FABP3基因表达量普遍高于公羊;在不同生长时期,FABP1、FABP3基因的表达量会随月龄的增加而逐渐上升。
