缙云山细枝柃种群性比及空间分布
2020-11-13敖艳艳李文巧杨思琴邓洪平
王 茜,敖艳艳,李文巧,秦 菲,杨思琴,丁 博,邓洪平,4,*
1 西南大学生命科学学院资源植物保护与种质创新重庆市重点实验室,重庆 400715 2 重庆山地型城市森林生态系统国家定位观测研究站,重庆 400036 3 西南大学生物技术中心,重庆 400715 4 重庆市科学技术研究院低碳与生态环保研究中心,重庆 401123
柃木属(Eurya)原属山茶科(Theaceae)厚皮香亚科(Ternstroemoideae)[1],APG系统将其列入五列木科(Pentaphylacaceae)[2]。该属植物广布于我国长江以南各省区,是常绿阔叶林中常见的下木,其泌蜜量大,花粉充足,是重要的冬季野生蜜粉源,对于蜂群安全越冬和春季的快速繁殖起重要作用[3]。该属植物具有较严格的雌雄异株性系统,但偶尔也会出现一些同时具有两性结构的变异株,如在细枝柃(E.loquaiana)[4]、钝叶柃(E.obtusifolia)[5- 6]及柃木(E.japonica)[7]中已有报道。区别于典型“雌雄异株”性别系统,这种变异株与雌、雄株共存的性别系统被称为“亚雌雄异株”,常被视为“雌雄同花”向“雌雄异株”进化的过渡[8]。课题组在调查中发现,缙云山细枝柃种群较大,分布于不同生境,且有较大比例的性别变异株,可为研究该属植物性别系统的进化与生态适应机制提供良好材料。
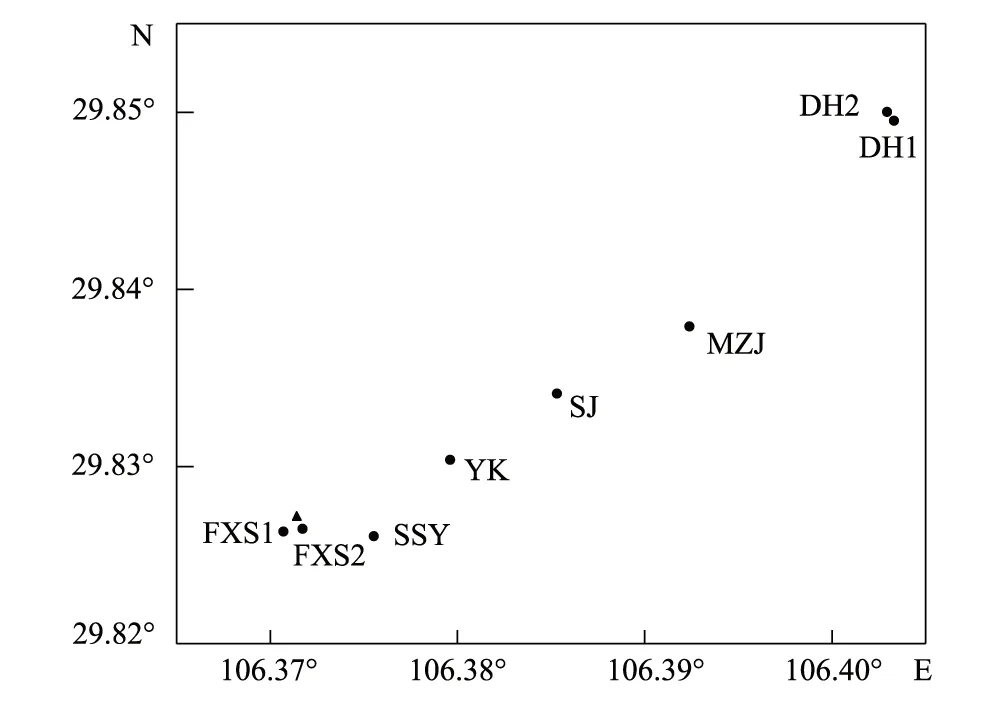
图1 样地分布位置图Fig.1 Distribution of sample plots “●”表示8个20 m×20 m样地中心点位置,DH1、DH2为黛湖1号和2号样地,MZJ为马中咀样地,SJ为三角花园样地,YK为垭口样地,SSY为舍身崖样地,FXS1、FXS2为复兴寺1号和2号样地;“▲”表示20 m×60 m大样地中心点位置
性别比例和个体空间分布是“雌雄异株”种群重要的结构参数,影响着种群的生存发展,甚至影响群落中生物间的相互关系[9]。“亚雌雄异株”种群由于包含一些两性个体,其结构较“雌雄异株”种群更复杂。本研究选择缙云山细枝柃种群为研究对象,通过分析细枝柃性比(包括雌雄比与性别变异株比例)及其随环境因子、年龄结构等的变化,同时分析其空间格局特点,为探索其性别格局成因,了解其性别系统进化及生态适应机制提供依据。
1 材料和方法
1.1 研究地概况
研究地位于重庆缙云山国家级自然保护区,地理坐标东经106 °17 ′43″—106 °24 ′50″,北纬29 °41 ′08″—29 °52 ′03″,最低海拔180 m,最高海拔952 m。该区为亚热带季风湿润性气候,年均温度13.6 ℃,年均降水量1243.8 mm,植被类型主要包括马尾松林、柏木林、栲树林、短刺米槠林、四川山矾林和竹林等[10]。
1.2 样地设置与群落调查
根据细枝柃分布,同时综合考虑海拔、植被类型、人为干扰等因素,在缙云山保护区内设置8个20 m×20 m的样地(图1)。调查样地内每株乔木的种名、高度、胸径和冠幅,再将各样地划分为16个5 m×5 m小样方,调查每个小样方内灌木的种名、高度、胸径和冠幅,草本的种名、平均高度和盖度。整理上述调查结果,使用以下公式计算重要值:重要值=(相对密度+相对优势度+相对频度)/300[11]。标记样地内每一株细枝柃,记录基径。2016—2018年,于细枝柃开花季节调查植株性别。为了避免微环境对花性别表型的影响,确定植株性别时至少观察不同方向的3个枝条,每枝至少观察50朵花。记录每个样地的海拔、郁闭度及人为干扰状况。在每个样地随机选取相距至少10 m的3个采样点,收集0—20 cm土样带回实验室检测。土壤全N采用凯氏消煮法(林业标准 LY/T 1228—1999),土壤全P采用酸熔—钼锑抗比色法(林业标准 LY/T 1232—1999),土壤全K采用酸熔—火焰光度法(林业标准 LY/T 1234—1999)。由于空间格局分析需要在一个面积较大的样地内进行[12],因此设置了1个20 m×60 m样地(图1),调查其中每株细枝柃的性别及坐标位置,分析个体空间分布格局。
1.3 数据处理与分析
以基径代替树龄进行年龄结构分析[13],根据细枝柃的生活习性和野外调查数据分布,将细枝柃划分为6个龄级,基径(D)≤1 cm为Ⅰ级,1 cm
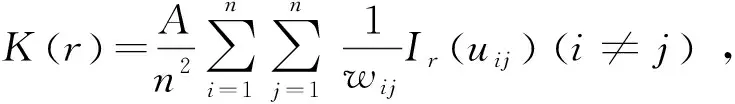
2 结果
2.1 细枝柃依存群落及生境
细枝柃种群主要分布于缙云山海拔大于500 m的区域。本次调查选择的8个样地,群落类型包括了暖性针叶林、暖性针叶阔叶混交林、竹林及常绿阔叶林(表1)。群落层次分明,乔木层针叶林中以马尾松、杉木为优势,常绿阔叶林中以香樟为优势,灌木层常以细枝柃、四川山矾为优势,草本层常以狗脊、红盖鳞毛蕨、落地梅为优势(表1)。
各样地均受到不同程度的人为干扰。其中SJ样地位于景区主干道旁,且周围住户较多,人为活动干扰度最高,DH1和SSY样地临近住户,受到的干扰也较大,FXS1和FXS2样地位于保护区缓冲区,受到的人为干扰较小(表2)。各样地土壤养分差异较大,DH1和DH2样地土壤相对贫瘠,全N、全P和全K的含量都相对较低,FXS1、FXS2及SSY样地土壤全N、全P含量相对较高,但土壤全K含量相对较低(表2)。

表1 各样地群落结构
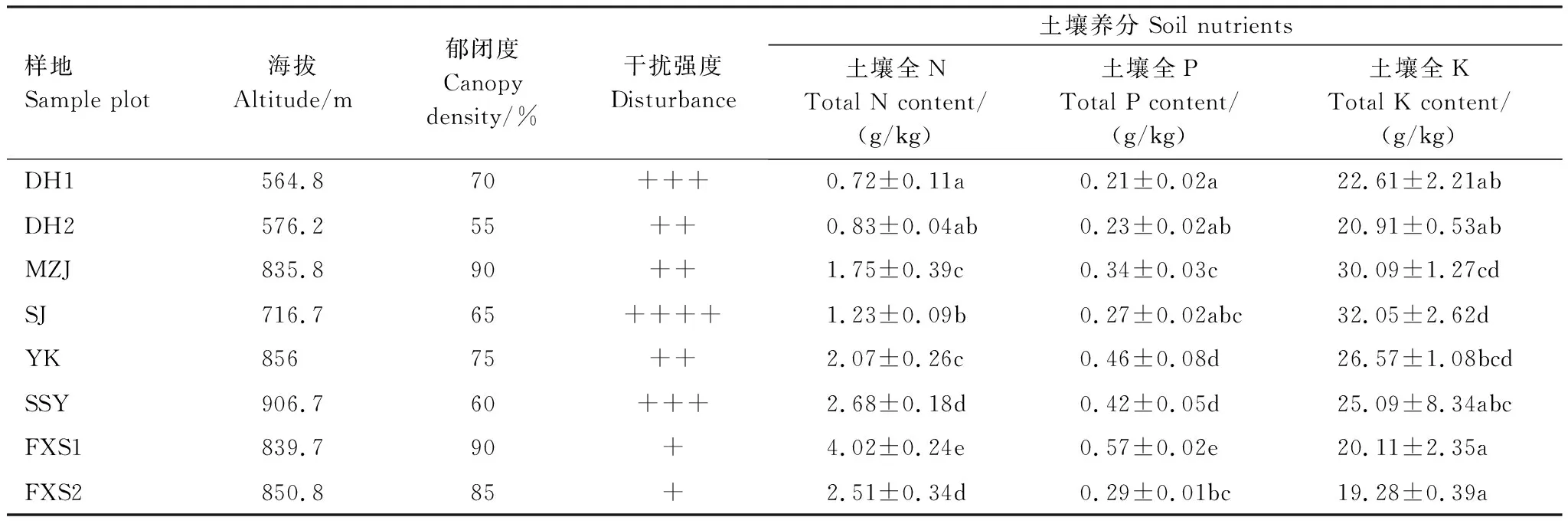
表2 各样地生境概况
2.2 细枝柃性比及其与环境因子等的相关性
8个20 m×20 m样地中共标记了312个个体,观察周期中个体性别表达稳定,仅有1株16年观测为雄性,后记录为性别变异株,统计中以性别变异株计。312个个体中雌株137株,雄株96株,性别变异株45株,性别不确定个体34株,性别变异株所占比例为14.42%。雌雄比1.43,卡方检验结果显示雌雄比明显偏离1∶1(Chi-Square=7.215,P<0.05),雌株显著多于雄株。各样地中仅DH1、SSY两个样地雄株偏多,其余均是雌株偏多。各样地中均有一定数量的性别变异株,其中YK样地性别变异株比例最高,达27.03%,FXS2样地性别变异株比例最低,仅2.63%(表3)。
RDA分析结果显示,细枝柃结构参数与环境因子的相关性较强,前两个环境因子轴的相关系数分别为0.974和0.992,且前两轴环境因子的解释度达98.4%(表4)。细枝柃各结构参数存在自相关:雌雄比与平均基径有一定正相关,与性别变异株比例有一定负相关;性别变异株比例与植株密度有较强负相关。性别变异株比例与干扰强度有强烈正相关而与海拔高度和郁闭度有强烈负相关,但雌雄比与各环境因素的相关性相对较弱(图2)。
Pearson相关性分析结果与RDA分析显示的趋势基本一致,中雌雄比与平均基径间具有显著相关性,Pearson相关系数达0.765(表5)。
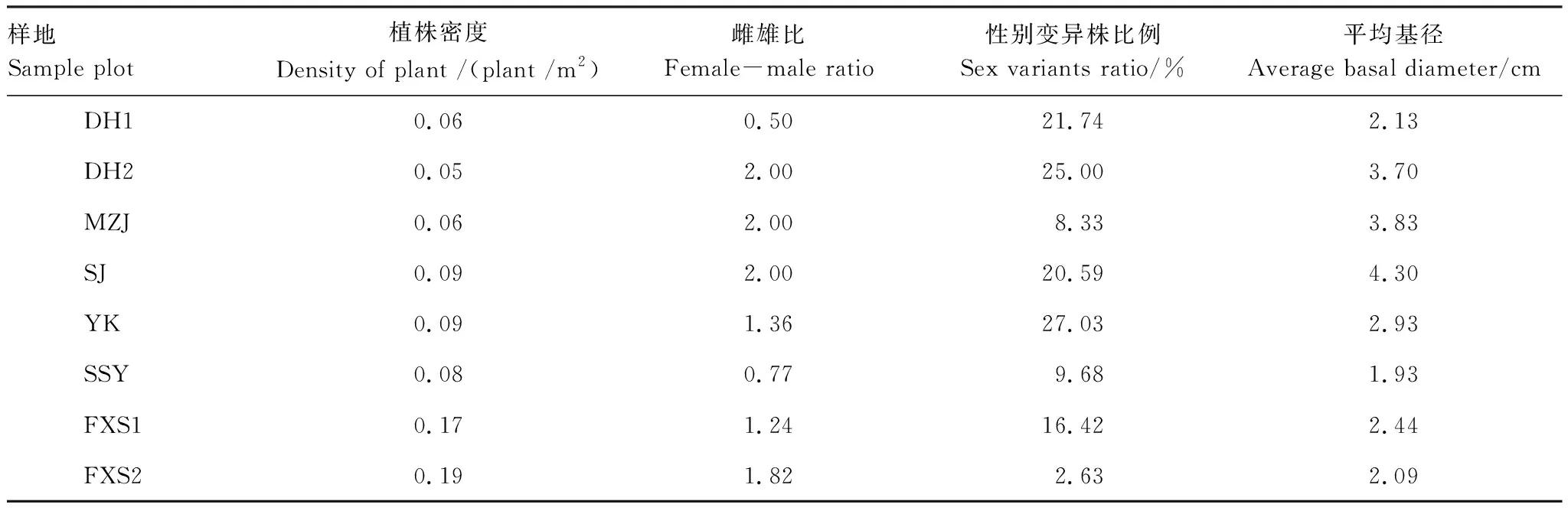
表3 各样地细枝柃结构参数
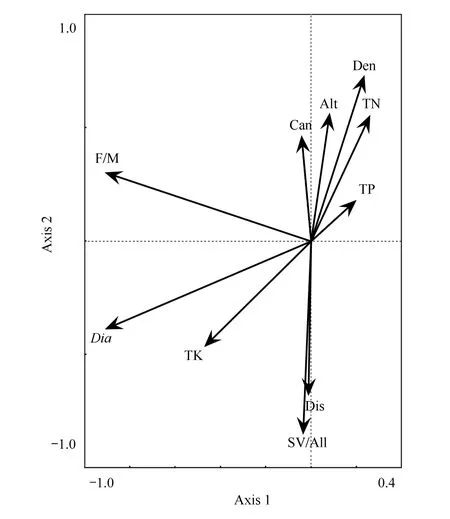
图2 细枝柃结构参数与环境因子之间的RDA二维排序图 Fig.2 RDA two-dimensional ordination diagram between environmental factors and structure parameters of Eurya loquaian Den:细枝柃密度,Density of plant;Dia:平均基径,Average basal diameter;F/M:雌雄比,Female-male ratio;SV/All:性别变异株比例,Sex variants ratio;Can:郁闭度,Canopy density;Alt:海拔,Altitude;Dis:干扰强度,Disturbance;TN:土壤全N,Total N content;TP:土壤全P,Total P content;TK:土壤全K,Total K content
2.3 性别比例在不同年龄结构中的表现
从龄级调查结果来看(图3),各样地中细枝柃以中龄和成龄个体为主,年龄结构为稳定型。其中SJ样地,缺少幼龄个体,而老龄个体相对较多,占总数的32%,偏向于衰退型。而FXS1、FXS2、SSY及DH1样地老龄个体缺乏或极少,幼龄和中龄个体较多,所占比例均大于70%,偏向于增长型。各样地中,雄株和雌株在各龄级中都有出现,性别不确定个体主要出现在幼龄阶段,各样地中性别变异株相对较少,一般分散出现在各龄级中。
将8个样地中所有细枝柃植株视为一个整体,性别变异株在各龄级中出现的比例差异不大,在Ⅴ级和Ⅵ级中相对较高,雌雄比在各龄级中均大于1,且在Ⅲ—Ⅵ级中,雌株的优势更为明显。但整体上,雌株、雄株及性别变异株基径无显著差异,仅性别不确定个体基径相对较小(图4)。
2.4 细枝柃植株的空间分布特征
20 m×60 m样地内共191株细枝柃,其中雌株82株,雄株63株,性别变异株13株,性别不确定个体33株,其空间分布如图5所示。点格局分析结果显示,细枝柃植株在0.5—5 m尺度上L(r)值高于上包迹线,呈聚集分布(图6)。雌株、雄株、性别变异株及性别不确定个体在0—5 m的尺度上L(r)值几乎都位于99%置信区间内,呈随机分布,仅雌株在0.8—1.7 m、雄株在大于1.5 m的尺度上、变异株在2 m的尺度上L(r)值稍高于或位于上包迹线附近,偏向于聚集分布(图6)。二元点格局分析结果显示,雌株和雄株有一定的正相关,且在0.8—2.5 m的尺度上达到显著水平(图6)。
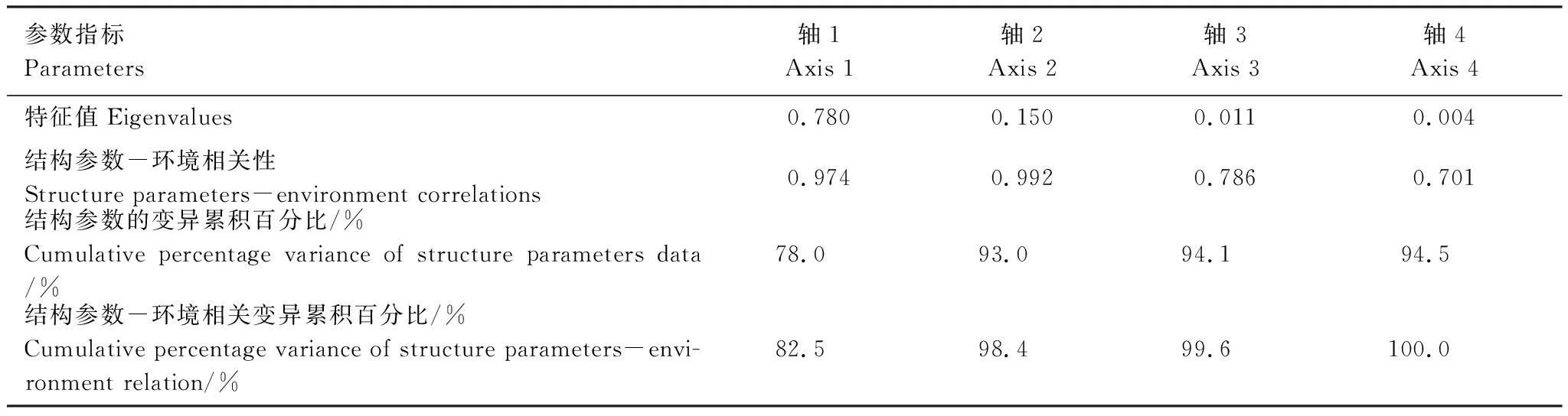
表4 细枝柃结构参数与环境因子之间的RDA分析结果
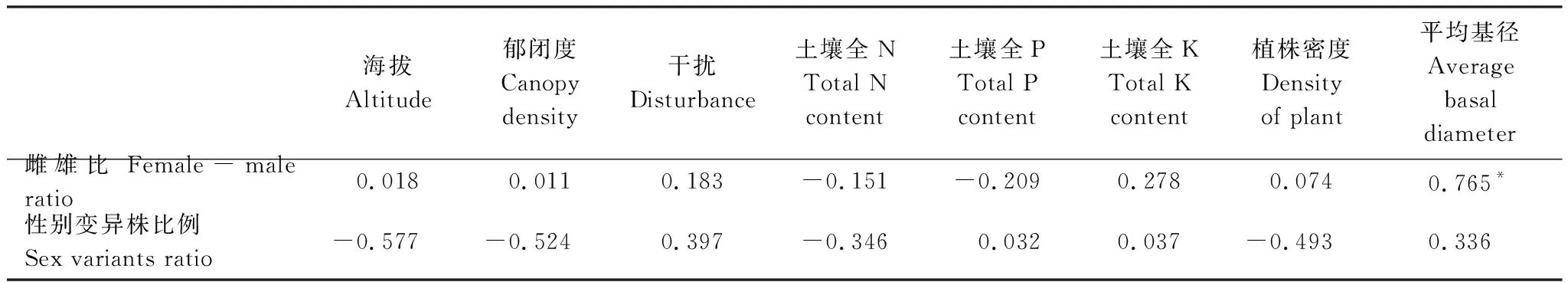
表5 细枝柃性比与其他结构参数及环境因子的Pearson相关系数
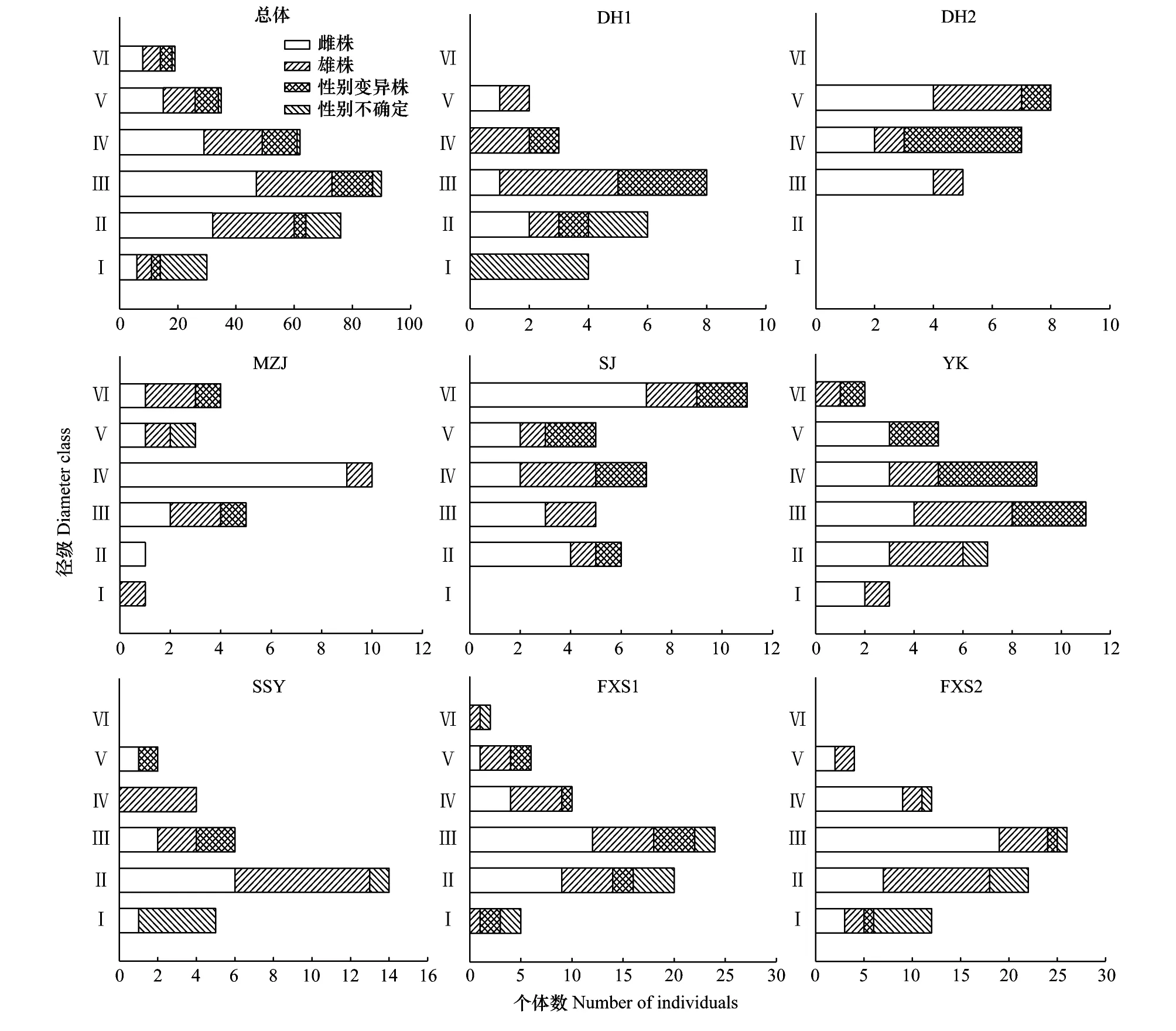
图3 各样地细枝柃年龄结构和性别组成Fig.3 Age structure and sex composition of Eurya loquaiana form each sample plotⅠ:基径≤1 cm,Ⅱ:1 cm<基径≤2 cm,Ⅲ:2 cm<基径≤3 cm,Ⅳ:3 cm<基径≤4 cm,Ⅴ:4 cm<基径≤5 cm,Ⅵ:基径 >5 cm
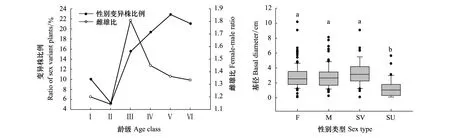
图4 性比在各龄级中的变化及各性别类型基径比较Fig.4 Variation of sex ratio in each age class and comparison of basal diameter between sex typesF:雌株,females;M:雄株,males;SV:性别变异株,sex uncertain ones;SU:性别不确定,sex uncertain ones
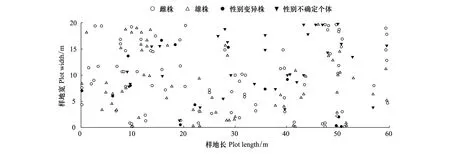
图5 细枝柃个体在20 m×60 m样地中的空间分布Fig.5 Spatial distribution of Eurya loquaiana plants in 20 m×60 m sample plots
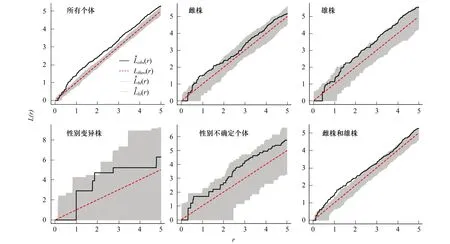
图6 细枝柃个体的空间分布格局Fig.6 Spatial distribution pattern of Eurya loquaiana plantsLobs为函数计算值,Ltheor为函数理论值,Lhi为上包迹线,Llo为下包迹线,阴影部分为99%置信区间
3 讨论
3.1 缙云山细枝柃种群的性比结构
缙云山细枝柃种群性别偏雌(雌雄比为1.43),并具有较高比例的性别变异株(占比14.42%),整体上呈现出“亚雌雄异株”状态。这一性比结构与Wang等研究的日本名古屋大学东山柃木种群相似:雌株占比33.2%,雄株占比30.1%,性别变异株占比34.1%[15]。雌雄比是性别分离物种的重要种群结构指标,影响着植物的繁殖成功率。在许多理论研究中,雌雄异株植物的雌雄比常被假定为1∶1[16- 17],但实际研究结果却显示大多数雌雄异株植物雌雄比偏离1∶1,且性别偏雄比性别偏雌更为常见[16]。性别偏向与多重因素相关,可能是由性别遗传机制决定,也可能跟植株寿命、种子传播和传粉机制相关[18]。虽然研究种群性别明显偏雌,但同期研究显示自然授粉与人工补充授粉下的雌株(或两性株)结实率没有显著差异,表明种群并没有因为雄株较少而出现花粉限制问题。另外,本研究中细枝柃种群并非典型雌雄异株种群,其中还包含一定比例的性别变异个体,它们在生殖中具有双重功能,也可能对偏离的雌雄性比起到一定调节作用。但变异株实际的雌性和雄性适合度并不清楚,进一步的研究可以考虑对变异株雌雄适合度进行量化,以便更准确地掌握种群的性别格局,并了解变异株在种群中的作用。
3.2 环境及年龄结构对性比的影响
虽然缙云山细枝柃种群整体偏雌,但在不同调查样地中雌雄比差异较大,且在DH1和SSY样地中,雄株偏多。虽然每个样地中都有性别变异株出现,但所占比例差异也较大。这可能与各样地中环境差异或细枝柃年龄结构差异有一定关系。有研究表明,海拔、光照、干扰等环境因素以及年龄结构都可能影响性别比例[19-22],甚至一些植物还会在生存期中出现性别转化[20,22]。本研究结果显示,细枝柃性比等特征与环境因子具有较强的相关性,其中变异株的比例与干扰强度有较强的正相关而与海拔高度和郁闭度有较强的负相关(图2)。柃木属植物在各植物志资料中均记载为雌雄异株[1,23],性别变异株较为少见。在对钝叶柃和细枝柃的形态学研究中发现,变异株中常存在退化的雌蕊和雄蕊[4,6]。Wang 等的研究也表明性别变异株与雌株相比具有更低的雌性适合度而与雄株相比具有更低的雄性适合度[24- 25]。一般认为,这种偶尔出现两性个体的“亚雌雄异株”状态,显示了雌雄同花向雌雄异株进化的最后阶段,具有两性功能的变异株由于雌雄适合度均较低而逐渐被单性个体代替,但也有人认为这一状态也可长期保持,或意味着雌雄异株性别系统的打破[8]。目前,并不清楚柃木属中性别变异株出现的原因,但与干扰强度的正相关性可能指示着变异株的出现与人为干扰有一定联系。在对非洲芙蓉属(Dombeya)和柿属(Diospyros)植物的研究中也显示性别变异株倾向于出现在受人类破坏、破碎化的种群中[26- 27]。变异株的出现可保证雌雄异株植物在单株、缺乏传粉媒介或种群较小的情况下完成生殖,可能是一种适应机制。
本研究设置的8个样地中,细枝柃年龄结构各有不同,SJ样地老龄个体相对较多,FXS1、FXS2、SSY及DH1样地则是幼龄和中龄个体较多。相关性分析结果显示,样地中细枝柃平均基径与雌雄比显著正相关(表5),对各龄级雌雄比的分析也显示,Ⅲ—Ⅵ级中雌株比例较Ⅰ—Ⅱ级更高(图4),表明可能随着林龄的增加,种群中的雌株比例有增加的趋势。类似的情况在秋枫属(Bischofia)、天南星属(Arisaema)等类群中也有报道,这可能与个体性别转化或雌株开花较晚等因素有关[20,28]。雌株在生殖中需要更多能量,因此可能需要更长时间的营养生长。本研究中还存在一些性别不确定个体,主要是一些幼株,若可以在不开花的情况下确定其性别,则可能对统计结果产生一定影响。柃木属植物中可能存在性别转化现象,Wang 等在对柃木的研究中发现,5年的观察期中,27.5%的个体至少发生一次性别转化,虽然观察期间性别比例没有明显波动,但长期来看有雌性个体增多雄性个体减少的趋势[15]。本实验3年的观察期中有1雄株,后记录为性别变异株(雄花两性花同株),因此缙云山细枝柃种群可能也存在性别转化现象,但发生的频率较低。
3.3 细枝柃植株的空间分布特征
植物营固着生长,每个种群都有自己的分布格局,是物种生物学特性、种间关系与环境条件综合作用的体现[29]。雌雄异株植物中常出现性别空间分离的现象,这主要是由不同性别个体对资源及相邻植株敏感性上的差异所引起的[30]。本研究结果显示缙云山细枝柃个体主要呈聚集分布,雌株、雄株及性别变异株主要呈随机分布,在部分尺度偏向于聚集分布。这种分布格局的形成,一方面与细枝柃的果实类型和种子传播机制有关:果实为浆果,主要靠鸟类传播种子;另一方面与个体的微环境偏好和个体竞争有关。与大多数雌雄异株植物出现性别空间分离不同,本研究中细枝柃雌株和雄株有一定的正相关,说明雌株和雄株对环境异质性的响应相似[31],这一特点可能在一定程度上增加授粉几率。
总的来看,缙云山细枝柃种群性别偏雌,雌雄比及性别变异株比例在不同样地和不同龄级中有一定波动;变异株的出现可能与干扰有关;随着林龄的增加,雌性比例趋向于增加;个体主要呈聚集分布,雌株和雄株有一定的正相关。这些特点反应了细枝柃种群结构的适应和调节机制。
